РОСТ_РАЗВИТИЕ.ppt
- Количество слайдов: 79



Рост и развитие растений

Особенности растений Способность к росту в течении всей жизни Рост растяжением Тотипотентность Вегетативное размножение

Рост необратимое увеличение размеров и массы клетки, органа или всего организма, связанное с новообразованием элементов их структур. отражает количественные изменения, сопровождающие развитие организма или его частей. Развитие • качественные изменения в структуре и функциональной активности растения и его частей (органов, тканей, клеток) в процессе онтогенеза. • возникновение качественных различий между клетками, тканями и органами – дифференцировка, а также возрастные изменения.
 биомасса (!но может и уменьшаться, напр. при стрессе) Сухая биомасса")
Параметры роста общая (сырая) биомасса (!но может и уменьшаться, напр. при стрессе) Сухая биомасса (!но может и уменьшаться, напр. при прорастании семян) Увеличение числа клеток (!но может и не изменяться, но происходит рост растяжением) Увеличение линейных размеров (высоты, ширины, длины и т. д. ) ростом можно назвать необратимое увеличение растения хотя бы по одному из параметров роста

Клеточные основы роста Онтогенез клетки – период от возникновения до смерти. Эбриональная фаза Фаза растяжения Дифференцировка, специализация (структурная, биохимическая, физиологическая или функциональная) ее

Клеточный рост Эмбриональная фаза – рост осуществляется за счет деления меристематической клетки с образованием дочерних. Дочерние клетки растут (за счет увеличения составных частей клетки), достигают размеров материнской клетки и снова делятся. Фаза растяжения – равномерное и неравномерное увеличение объема; за счет вакуоли - (рост колеоптиля, раскрывание почек, удлинение тычинок, вытягивание спорангия и др. )

Дифференцировка Фаза дифференциации – происходит превращение клетки в специализированную, т. е. выполняющую определенную функцию: водопроводящую (сосуды, трахеиды), проводящую орг. вещ-ва (ситовидные трубки), запасающую (паренхима), механическую и др.
 I- лаг-фаза, II - фаза экспоненциального роста; III")
Кривая роста клеток (S-образная кривая роста) I- лаг-фаза, II - фаза экспоненциального роста; III - фаза замедления роста; IV - стационарная фаза; V - фаза деградации

Дедифферинцировка. Тотипотентность. Дедифференцировка – это переход специализированных неделящихся клеток к делению, т. е. восстановление их меристематической активности, потеря специализации. Свойство только растительных клеток. Происходит, напр. , при образовании раневого каллуса в месте повреждения. Не все клетки могут ДД. Легко: паренхимные, эпидермальные, железистые. Не могут: мертвые клетки ксилемы, ситовидные трубки, механические волокна.
 - свойство клетки реализовать при")
Дедифферинцировка. Тотипотентность (totalis - общий, целый; potentia - способность) - свойство клетки реализовать при определенных условиях имеющуюся у нее генетическую информацию и дать начало целому организму. Морфогенетический ландшафт Уоддингтона Терминальная дифференцировка необратимый процесс, после которого клетка не может дедифференцироваться и реализовать свою тотипотентность.
 – индивидуальное")
Периодизация индивидуального развития Онтогенез – (ontos – существо, genesis – процесс образования) – индивидуальное развитие организма от зиготы (или вегетативного зачатка) до естественной смерти. В ходе онтогенеза реализуется наследственная информация организма (генотип) в конкретных условиях среды, в результате чего формируется фенотип, т. е. совокупность всех признаков и свойств данного индивидуального организма.
 генеративный сенильный")
Периодизация индивидуального развития эмбриональный ювенильный виргинильный (не всегда) генеративный сенильный

Эмбриональный Начинается со слияния яйцеклетки и спермия т. е. с образования зиготы Формирование зародыша (гетеротрофное питание за счет материнского растения). Формирование семени. У однодольных – одна семядоля, у двудольных – 2 семядоли. Заканчивается периодом покоя.
 От прорастания семени до начала заложения первых цветков. Образуются только вегетативные органы:")
Ювенильный (молодость) От прорастания семени до начала заложения первых цветков. Образуются только вегетативные органы: листья, стебли, корни. Максимальная активность всех физиологических функций, поэтому устойчивость минимальна. Большая способность к корнеобразованию: черенки, срезанные в этот период легко укореняются. Виргинильный (иногда) растение по всем признакам не отличается от генеративного, но еще не цветет.
 Период цветения и плодоношения Формирование репродуктивных органов растения. Оплодотворение, Образовнаие, рост и")
Генеративный (зрелость) Период цветения и плодоношения Формирование репродуктивных органов растения. Оплодотворение, Образовнаие, рост и созревание плодов и семян. Торможение роста вегетативных органов. В пределах одного онтогенеза начинается новый онтогенез – доминирующий над другими процессами.
 Период от полного прекращения плодоношения до отмирания всех вегетативных органов и смерти")
Сенильный (старость) Период от полного прекращения плодоношения до отмирания всех вегетативных органов и смерти всего организма. Старение- усиливающееся с возрастом ослабление жизнедеятельности, приводящее к естественной гибели. Изменение состава гормонов Снижение активности всех ферментов кроме гидролитических Прогрессирующее нарушение синтеза белков Уменьшение скорости физиологических процессов Процессы распада преобладают над синтезом Накопление веществ, не участвующих в обмене Нарушение структуры и роницаемости мембран

Гипотезы старения Первая группа гипотез предполагает, что старение и смерть — это процесс, заложенный в геноме. Программированная смерть. Отжившие свое особи неизбежно должны уступить место (пространство, ресурсы) своим молодым потомкам. Поэтому Природа позаботилась о непрерывности жизни, заложив в генах специальную программу старения, то есть приказ о самоуничтожении. Реальность программы старения подтверждается существованием предельного числа клеточных делений (предел Хейфлика), открытием теломеразного счетчика деления клеток. Вторая группа гипотез связывает старение с постепенным накоплением случайных ошибок в экспрессии генов. Аккумуляция мутаций. Действительно, эволюция старалась изо всех сил и приспосабливала организм к условиям среды, развивала устойчивость к заболеваниям, но вот отлаженный организм в какой-то момент перестает быть адекватным среде, стареет. Это означает, что старость неадаптивна, что это накопление неполадок в организме. Например, знаменитая среди биологов резуховидка Arabidopsis thaliana при накоплении мутаций становится менее устойчивой и менее плодовитой, сроки ее жизни сокращаются.

Гипотезы старения С другой стороны, у долгоживущих растений активизируется починка испорченных генов, количество мутаций неизменно снижается. Кроме того, несмотря на предполагаемое увеличение числа соматических мутаций, «растения могут вообще не проявлять признаков старения даже в конце жизни. Так, у 5000 -летней сосны Pinus longaeva отсутствуют признаки мутационного старения» . И первая и вторая группа гипотез логически оправданы и имеют в своем арсенале достаточное число фактов, но всё же диаметрально противоположны: в первом случае старость считается закономерным процессом, во втором — стохастическим.

Гипотезы старения Как естественное продолжение и дополнение гипотезы накопления мутаций предложены концепции «восстановления сомы» и близкие к ней идеи. Организм имеет ограниченные возможности на починку генетических неисправностей: если растение бросает все силы на выращивание цветов и семян, то на исправление неполадок в остальных частях растений ресурсов попросту не хватает. Потому во время цветения само растение быстро портится, то есть стареет. Исходя из этой гипотезы затраты на цветение и плодоношение должны снижать сроки жизни растения. У некоторых линий бобовых это действительно так.

Гипотезы старения Третья группа гипотез считает, что старение — это период жизни, упущенный отбором. Отбор призван обеспечить репродуктивный успех вида, следовательно отбору всё равно то, что случается после благополучного выведения и выживания потомства. Даже больше: гены, обеспечившие репродуктивный успех в молодости, могут потом оказаться вредными для организма. Чтобы подтвердить эти гипотезы, нужно сравнить эффективность плодоношения и скорость последующего отмирания. Чем успешнее плодоношение, тем быстрее должны постареть «отслужившие свое» организмы. Эта гипотеза подтверждается фактами и расчетами, в том числе и примерами из жизни растений. Однако есть примеры, которые не укладываются в эту гипотезу. Так, старение туи (Thuja occidentalis) зависит не от скорости созревания семян, а от условий произрастания.

Возрастные изменения Морфогенез – формообразование, включает процессы роста и развития клеток- цитогенез Тканей – гистогенез Органов – органогенез Которые генетически запрограммированы и скоординированы между собой.

Продолжительность онтогенеза и его типы продолжительность от 10 -14 дней до 3 -5 тыс. лет, разные этапы отногенеза могут иметь разную продолжительность эфемеры однолетние двулетние многолетние

Продолжительность онтогенеза и его типы Независимо от продолжительности онтогенеза растения делятся на 2 группы монокарпические растения, цветущие и плодоносящие один раз в жизни агава бамбук онтогенез разный по продолжительности, но после плодоношения погибают поликарпические растения, цветущие и плодоносящие много раз в жизни

Системный контроль развития Ростовые корреляции Корреляция влияние одних частей на скорость и рост других стимулирующие • Корень стимулирует рост побега. • Формирующиеся семена стимулируют рост околоплодника. тормозящие • Образование цветков тормозит заложение и рост листьев (гречиха при удалении цветков дает удвоенный урожай зеленой массы). • Пасынкование – удаление боковых побегов(пасынков) для улучшения роста плодов (томаты) • Апикальное доминирование
 под влиянием верхушки побега")
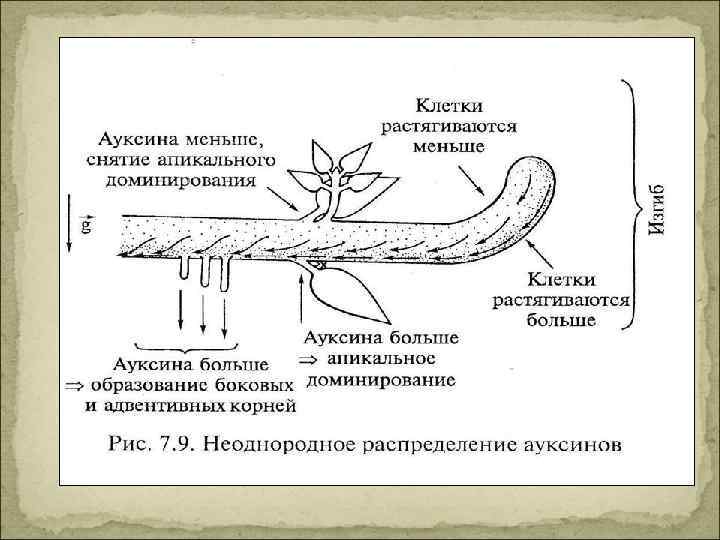
Ростовые корреляции Апикальное доминирование – торможение роста боковых побегов (корней) под влиянием верхушки побега (корня). Обусловлено фитогормонами. • Полное подавление ветвления • Замедление роста боковых побегов (корней) • Подавление вертикального роста боковых побегов (изменяется угол ветвления) Пикировка Прищипка гл. корня для увеличения роста боковых корней, (рассаживание сеянцев их общей емкости в индивидуальную) Вершкование Удаление соцветий или верхних листьев, для увеличения роста листьев (табак)

Генетические основы развития Растение имеет более 25 000 генов: конститутивные и индуцибельные. Активность более половины генов регулируется. Основа процесса дифференцировки – дифференциальная активность генов. Она выражается в различном составе фракций м. РНК или набора белков различно дифференцированных клеток.

Генетическая система растительной клетки

Геном – суммарная ДНК клетки, содержащая все гены, а так же все межгенные области (контроль транскрипции) Транскриптом - совокупность всех м. РНК (контроль трансляции) Протеом – совокупность белков, которые определяют функциональную специфичность клетки (биосинтез белков, распад белков, биогенез клеточных органелл); Метаболом – совокупность всех метаболитов, свойственных клетке, ткани или органу на определенном этапе онтогенеза в зависимости от условий среды.
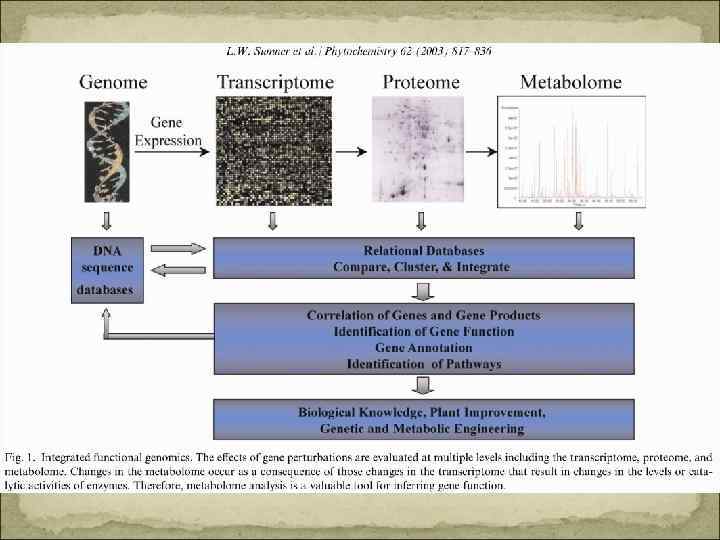
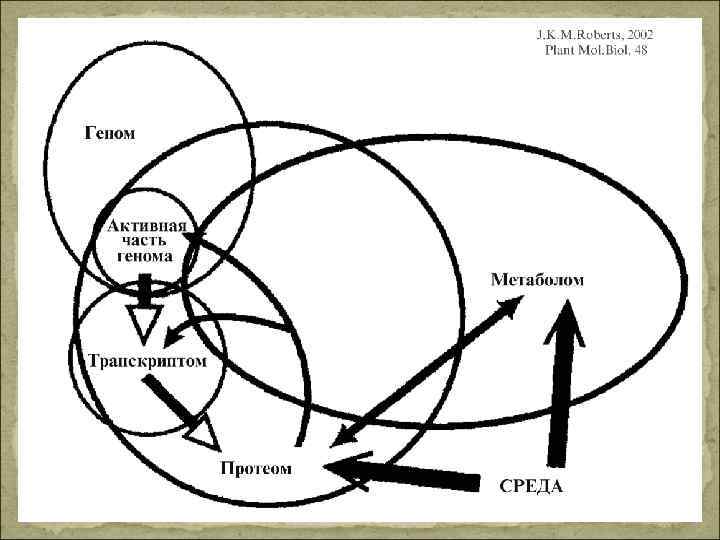

Контроль развития фитогормонами Гормоны – низкомолекулярные сигнальные вещества, которые синтезируются у всех или у многих растений в низких концентрациях (10 – 6 М) и вызывают характерные физиологические реакции. Места образования и действия гормонов разделены в пространстве. Однако могут образовываться вблизи места действия (аналог паракринного или автокринного сигнального вещества животных)- передвижение диффузией. Малая ткане и органоспецифичность – широкий спектр действия: фитогормон как раздражитель. Действие гормона зависит от от набора активных генов. Концентрация гормона (синтез, распад, коньюгация, запасание, отток) точно регулируется. Чувствительность органов сильно различается. Недосатток и избыток – нарушение. Для межклеточной регуляции в многоклеточном организме. Они контролируют процессы роста и дифференцировки, взаимодействуют. Редко регулируют обмен веществ уже дифференцированного организма.

Ауксины: ИУК Наиболее распространен ИУК – индолил-3 -уксусная кислота. Встречается у всех про-и эукариот, но только у высших – сигнальное вещество. Другие ауксины – фенилуксусная кислота у табака, индолакриловая и галогенпроизводная индолуксусной кислоты у бобовых не имеют всеобщего значения. Синтетические – 2, 4 -дихлорфеноксиуксусная (2, 4 -Д), 1 нафтилуксусная (НУК) и индолмасляная кислота (ИМК), из которых в растении может образовываться ИУК.

Строение, синтез Места образования – эмбриональные ткани, фотосинтезирующие органы, корневая система. ИУК образуется из триптофана. Может синтезироваться эпифитными бактериями и микроорганизмами (грибами) ризосферы. Транспортная и запасная формы гормона. Распад при участии ИУК-оксидазы. Транспорт – с током ассимилятов по флоэме, паренхимный транспорт полярный транспорт: активный базипетальный и пассивный акропетальный.

Действие ауксинов 1. Стимуляция деятельности камбия для увеличения числа ксилемных элементов 2. Стимуляция образования и развития семян и плодов (пыльца с ИУК, семяпочка с ИУК). Величина плода пропорциональна числу семян. Партенокарпия – обработка ИУК (бессемянные плоды) Стимуляция закладки боковых и адвентивных корней – ризогенез Ауксин как гербицид (при избытке ауксина – синтез этилена и ингибирование) Индукция регенерации в культуре клеток Апикальное доминирование - ингибирование прорастания боковых почек (цитокинин стимулирует боковые, а ауксин – ингибирует апикальную) Ингибирование опадения листьев, цветков и плодов (недостаток ауксина – активация АВК и этилена) Стимуляция роста растяжением. Участие ауксина в гравитропизме (крахмальные зерна – статолиты реагируют на восприятие силы тяжести и это приводит к перераспределению потоков ауксина)

Апикальное доминирование

Стимуляция растяжения колеоптилей ИУК

Механизм кислого роста ИУК индуцирует подкисление апопласта, индуцирует выход ионов водорода и активирует синтез структурных элементов клеточной стенки Ослабление водородных связей в клеточной стенке и активация ферментов разрушающих ковалентные связи в клеточной стенке В клеточную стенку вставляются вновь синтезируемые элементы

Геотропизм ауксины
")
Транспорт ауксинов Открытие факта передвижения ростового вещества, синтезируемого в апикальной меристеме (ауксина)

Транспорт ауксинов 1. Полярный транспорт и движение в клетку по градиенту концентрации; выход активный с трансмембранным переносчиком 2. ИУК из апопласта 3. Канал – белок (AUX 1 b) , белокпереносчик ( influx carrier) 4. ИУК – ИУК + К

Молекулярный механизм действия Регулируемые ауксином процессы протекают путем изменения экспрессии генов ИУК индуцирует протеолитический распад репрессорного белка и начинается транскрипция регулируемых ауксином генов.

Цитокинины – N 6 - замещенные пурины. Работы Ф. Скута – 50 е годы ХХ века
")
Биосинтез и свойства Образуются из аденина. Изопентениладенин – самый простой цитокинин (самая высокая активность) – процессы гидроксилирования, окисления и восстановления – другие цитокинины. Активная и неактивная формы. Неактивная – связана с сахаром , а активная - свободная. Выделены гены – их белки рецепторы цитокининов. Синтез цитокининов – апексы корня, молодые листья и развивающиеся семена. Транспорт – по ксилеме, транспортная форма – транс-зеатин-рибозид; на близкие расстояния – диффузия. Транспортируется не полярно и пассивно. Форма – транспортная (без сахара) , запасная (с сахарами), инактивирующая (образуется за счет удаления пренилового остатка в молекуле)

Действие цитокининов Стимуляция деления клеток. Способствуют синтезу ДНК в клетке, контролируют S-фазу клеточного цикла у растений. Эффект связан с действием ауксина. Взаимодействие ауксинов и цитокининов в культуре in vitro. Цитокинин ответственнен за регенерацию корней, а ауксины – побегов. Опухоли (коррончатые галлы) – повышение содержания обоих гормонов. Генетически обусловленные опухоли (у межвидовых гибридов) – нарушение контроля клеточного деления.
 – стимулируется цитокинином, вырабатываемым грибом Бактерии")
Нарушение апикального доминирования «Ведьмины метлы» (прорастание боковых почек) – стимулируется цитокинином, вырабатываемым грибом Бактерии вызывают образование каллуса (накопление цитокинина), который приводит к притоку ассимилятов. Образование лентовидных, расплющенных побегов (fascian) – одновременно растущие боковые побеги Цитокинины стимулируют: растяжение клеток при развитии листа; развитие хлоропластов; индукцию почек на каулонеме мхов.
 Монокрапики. Старение и смерть после цветения и плодоношения (")
Замедление процессов старения (гормон омоложения) Монокрапики. Старение и смерть после цветения и плодоношения ( высокая потребность в цитокининах репродуктивных органов приводит к дефициту в других органах). Поликарпики (недостаток питания). В процессе старения – снижение уровня фитогормонов. В большей степени цитокинина.

Аттрагирующее действие - регуляция донорно - акцепторных отношений Вещества в растении перемещаются от места низкой концентрации цитокинина к высокой. При высокой концентрации цитокининов: ингибируется образование боковых корней и индуцируется боковые почки. Цитокинины снимают апикальное доминирование. При обработке цитокининами бессемянных плодов – стимуляция роста. В норме в семянных плодах накапливаются цитокинины, а в бессемянных нет.
")
Физиологическая роль Влияние на устьичные клетки: если воды в устьица из корней (богата цитокининами) – устьица открываются, если из других органов (бедна цитокининами) устьица закрываются.
– двухкомпонентные")
Молекулярные механизмы действия цитокининов Под контролем генов. Рецепторы цитокининов (СKII и CREI )– двухкомпонентные рецепторы киназ и локализованы в плазмалемме Цитокинин связывается рецептором, фосфорилируются, фосфатная группа переносится на аспартат и белки АНР – семейства. Фосфорилированные белки АНР переносятся в ядро и инициируют транскрипцию.

Гиббереллины Гибреллины – дитерпены с общим скелетом – тетрациклический энт -гиббереллан. Всего описано более 100 структур, физиологическую активность имеют только некоторые.
.")
Открытие ГБ Рис в Японии – болезнь «баканэ» - бешеные проростки (слабые механические ткани). Фактор – гибберелловая кислота. Номенклатура гшиббереллинов: гиббереллин +А+ число. Известно А 1 – А 116 (GA 1 - GA 116). Важнейшие активные гиббереллины – GA 1, GA 4 и GA 3 (гибберелловая кислота)

Синтез гиббереллинов

Синтез, транспорт ГБ Место синтеза – быстро растущие ткани (меристемы побега, растущие листья, кончик корня) , на ранних этапах развития семян. Больший синтез в листьях, но могут и в корнях. Транспорт – в побеге не полярный, в корне – слабо полярный. На малые расстояния – диффузия, на большие – с массовым потоком по ксилеме и флоэме. Транспорт в связанной с сахаром форме – гибберелин-гликозиды или эфиры. Активные – свободные гиббереллины, не активны – гиббереллин – гликозиды или сложные эфиры.

Молекулярные механизмы действия гиббереллинов у прорастающих семян

Физиологическое действие ГБ 1. Стимуляция растяжения междоузлий. Гиббереллин индуцирует синтез фермента, который гидролизует гемицеллюлозную сеть первичных клеточных стенок. Это обеспечивает легкий доступ экспансина к водородным связям и приводит к нарушению связей между целлюлозой и гемицеллюлозой. В отличие от ауксина нет подкисления. Интеркалярный рост Весенний рост за счет интеркалярных меристем. Почки яблони, березы, тополя покрыты почечными чешуями (листьями - катафиллами). Между ними есть междоузлия, которые растут слабо (мало ГА). Все вместе – почечное кольцо. Выше настоящие листья и междоузлия большие. Чем крупнее лист, тем сильнее синтез гиббереллинов, тем больше растяжение междоузлия. У крушины, облепихи почечных колец нет. Их почки покрыты хорошо развитыми листьями, которые дают гиббереллиновый сигнал вставочным меристемам , междоузлия растягиваются и образуется листовая мозаика. У розеточных растений сигнал ГА поступает из ниже лежащих листьев в апикальную меристему и формируется длинный цветонос.

2. Стимуляция прорастания семян. При прорастании из зародыша идет гиббереллиновый сигнал от щитка в эндосперм. В алейроновом слое синтезируются ферменты для гидролиза запасных метаболитов. Крахмал распадается до мальтозы и глюкозы, они поступают в щиток и затем к тканям зародыша. Можно использовать искусственно низкие концентрации активного гиббереллина. Использование в пивоварении. Пророщенный ячмень- разрушение крахмала- варка в воде – упаривание и получение солода. Можно получать солод из плохопрорастающих семян при обработке их ГК (разрушение крахмала).

Гиббереллин и проявление пола у растений Огурец и конопля. Обработка ГБ – увеличение закладки мужских цветков(гормон антагонист – цитокинин). Кукуруза. Обработка ГБ в мужской метелке женские цветки. Мужская стерильности у риса.

Гиббереллин и цветение растений ГБ стимулируют цветение. При увеличении длины дня увеличивается количество ГБ. У растений с фотопериодическим контролем цветения на длинном дне, можно вызвать цветение добавкой гиббереллина. К ГБ чувствительны каланхое, морковь, рудбекия.
 путем")
Абсцизовая кислота как БАВ Синтез. В высших растениях из каротиноидов (неоксантин и виолаксантин) путем ослительного расщипления до абсцизовой кислоты. Накопление. Во всех органах. Больше осенью в покоящихся семенах, а так же в семенах и плодах. Может синтезироваться в неблагоприятных условиях в корнях. Транспорт. По флоэме и ксилеме. На короткие – диффузия. В черешках – транспорт поляризован.

1. Регуляция водного режима. При недостатке воды: АБК стимулирует закрытие устьиц, повышение гидравлической проводимости корня и стимулирует рост корня при ингибировании роста побега. Регуляция работы устьиц При закрытых устьицах соджержание АБК в замыкающих клетках в 20 -25 раз выше, чем при открытых. У мутантов по АБК, ее содержаниесоставляет 10% от дикого типа.

Физиологическое действие АБК 1. Регуляция водного режима. Нехватка воды – денатурация биологически активных молекул и нужно сохранить онформационную структуру ДНК, РНК и белков. В ответ на АБК – повышение уровня осмотически активных веществ (пролина и др. ), которые связывают воду, и появление гидрофильного белка – осмотина. В ответ на АБК – синтез полиаминов, котрорые ассоциируют с ДНК и РНК и становятся более устойчивыми к обезвоживанию. В ответ на АБК – устьица закрываются через 10 -15 минут. При сильной засухе – опадение листьев. При увеличении концентрации осмотически активных веществ - синтез абсцизинов, что приводит к повышению АБК и растения летом могут перейти в состояние физиологического покоя.

2. Регуляция покоя семян АБК ингибирует прорастание – важный фактор покоя. АБК – эмбриональный фактор прорастания. Покой определяется содержанием АБК зародыша. Когда зародыш семени достигает окончательного размера синтезируется АБК индуцирует синтез запасных веществ семени. ДНК и РНК упаковываются белкми-шаперонами. Полиаминами, рост прекращается и идет обезвоживание. АБК мешает прорастанию. Она может разрушаться после дождей (даже осенью) или зимней прохлады. У некоторых видов она легко вымывается, у других – трудно. У семян грецкого ореха. Яблони стратификация уменьшает АБК и стимулирует синтез гиббереллинов. Яровизация семян приводит к снижению синтеза АБК и повышению содержания гиббереллина и цитокинина – выход из покоя.

3. Синдром дефицита АБК При недостатке АБК растения жизнеспособны, но не переносят стрессы. Мутанты по АБК: семена не накапливают запасные вещества, нет состояния покоя, смена прорастают сразу после созревания в плоде (вивипария).

4. Регуляция формы листьев АБК Два типа листьев: подводные листья над водой – синдром засухи и синтез АБК – другая форма. Смена генераций листьев у полупустынных растений контролируется АБК.

Этилен – газообразный гормон Открыт в 1901 Д. Н. Нелюбовым в СПГУ Синтез. Цикл Янга синтез этилена и регенерация метионина.
. Которые")
Рецепция этилена Имеется семейство белков-рецепторов этилена. (ETR 1, ETR 2 и др. ). Которые гидрофобны и входят в состав мембран. Для связывания этилена необходим атом меди, который входит в рецепторный комплекс. Пути восприятия этилена продублированы несколькими рецепторами. Поэтому нечувствительные растения получить трудно. Это говорит о важности этилена.
 Тройной ответ (по Нелюбову). В ответ на препятствия росту")
Физиологическое действие этилена (механический стресс) Тройной ответ (по Нелюбову). В ответ на препятствия росту в почве растение в апикальной меристеме синтезирует этилен и растет по пути наименьшего сопротивления. У этиолированных проростков добавка этилена вызывает: угнетение роста растяжением, усиление поперечного роста и нарушение отрицательного гравитропизма, образование изогнутой апикальной петельки. При выходе на свет синтез этилена ингибируется и идет нормальный рост проростка. Усиленный поперечный рост идет за счет с переориентацией кортикальных микротрубочек из поперечного в продольное положение – изменяется ориентация целлюлозных микрофибрилл в клеточных стенках.
")
Этилен и прикосновение Показано, что при опрыскивании (изменение влажности, тень водяных брызг, механическая нагрузка) водой у арабидопсиса синтезируются новые м. РНК. Оказалось – механическая нагрузка – сопрокосновение. Эти гены – ТСН (touch – прикосновение). Похожи на кальмодулины. Растения при частом беспокойстве отстают в росте. Но становятся механически более прочными. Этот эффект похож на действие этилена.

Этилен и заживление ран Растения с млечниками образуют латекс. Внутри растения он жидкий, а при выступлении на поверхность при ранении затвердевает, закупоривая рану. Латекс застывает при действии этилена (заживление ран). При сборе каучука – растения обрабатывают ингибиторами синтеза этилена. Этилен активизирует раневую перидерму: образуется пробковый камбий, который формирует пробку, отделяющую здоровую ткань от поврежденной. Пробка – гидрофобный барьер (от испарения воды и попадания инфекции). При нападении насекомых и клещей выделяется этилен и в основании листа синтезируется перидерма – лист опадает.
, эта раневая поверхность закрывается")
Регуляция листопада При опадении листа формируется отделительный слой (раневая перидерма), эта раневая поверхность закрывается пробкой. Отделение протекает либо через растворение срединных пластинок, срединных пластинок и первичной стенки, либо всей клетки. Этот процесс контролирует этилен.

Опадение листьев Фаза 1. Приток ИУК, клетки отделительного слоя не реагируют на этилен. Фаза 2. Поток ИУК снижается, клетки чувствительены к этилену. Фаза 3. Клектки чувствительны к этилену и образуют гидролитические ферменты. Идет дифференцировка отделительного слоя и опадению листьев

Формирование и созревание плодов Оплодотворение завязи. На рыльце пыльцевые зерна, прорастают и давят на ткань столбика – ткани столбика выделяют этилен. Реакция частей цветка: все органы, привлекающие опылителей отмирают; у белокрыльника изменяется цвет покрывала соцветия; тычинки увядают, а завязи активно растут. Созревание сочных плодов: плод перестает расти, начинается размягчение (пектиназы выделяются в апопласт); в ножках плодов активируется отделительтный слой; меняется р. Н – плоды становятся менее кислыми; меняется желтая окраска на красную. Климатрические плоды (яблоки, груши, бананы, авокадо) –этилен ускоряет созревание. Бананы. При транспортировке их держат в атмосфере лишенной этилена (воздух пропускают через бромированный угольный фильтр и примешивают как онтогонист СО 2). Перед продажей – повышают температуру, удаляют СО 2 и обрабатывают этиленом. При нарушении синтеза этиолена плоды не дозревают. «Гнилое яблоко» - источник этилена, который вызывает размягчение и других яблок.

Биотический стресс Травоядные животные повреждают растения, а они в ответ выделяют этилен, который запускает синтез часто токсичных для животных веществ. Африка. Антилопы в загоне, рядом заросли акации – природного корма антилопы. Животные гибли от отравления таннинами. Оказалось, что антилопы повреждают деревья. А они в ответ выделяют этилен. Который индуцирует синтез таннинов. В природе антилопы не живут на одном месте и уходят от поврежденных деревьев с таннинами. В растении под действием этилена могут вырабатываться ингибиторы протеиназ – ферменты необходимые для переваривания пищи. При механическом повреждении через этилен идет синтез ингибиторов протеиназ, которые попадают в желудок травоядных животных и вызывают нарушенич пищеварения.

Этилен и цветение У огурца этилен повышает долю мужских цветков. А у бромелиевых индуцирует цветение. Для синхронизации цветения на плантациях ананаса растения обрабатываются этефоном, которая в водном растворе медленно разлагается до этилена, фосфата и хлорида.

Регуляция цветения у бромелиевых Это эпифиты. Листья в розетке, в которой собирается вода, пыль, листовой опад, выводят птомство комары и др. Воду поглощают листья. В центре розетки – цветонос, который может повредиться и вода выливается. Этот стресс вызывает образование этилена – сигнал к быстрому развитию- образованию семян.

Рецепция и передача сигнала этилена У Арабидопсиса известно 5 рецепторов этилена. Этилен связывается с Си-ионами рецепторов. В отсутствии этилена рецепторы подавляют сигнальный путь этилена, активируя протеинкиназу, которая является отрицательным регулятором этого сигнального пути. Если этилен связывается с рецептором, тотрецепторы перестают взаимодействовать с протеинкиназой и киназа активируется.

СИНТЕТИЧЕСКИЕ ГОРМОНЫ Экзогенные стимуляторы используются при предпосевной обработке семян, укоренении черенков, пересадке, для предупреждения преждевременного опадения плодов. Например, синтетический гиббереллин используется для получения бессемянных плодов винограда, получения большого числа листьев у чая и т. д.

Синтетические ингибиторы Гербициды – для борьбы с травянистыми сорняками, Арборициды – для борьбы с древесными сорняками Они обладают избирательностью – нпр. уничтожают двудольные сорняки злаковых, не повреждая самих злаковых. Ретарданты – для предовращения полегания растений. Ингибируют рост стебля и активируют развитие механической ткани. Дефолианты и десиканты – при опрыскивании ими происходит опадение или подсыхание листьев, нпр. у хлопчатника, картофеля, что облегчает механизированную уборку урожая.
РОСТ_РАЗВИТИЕ.ppt