
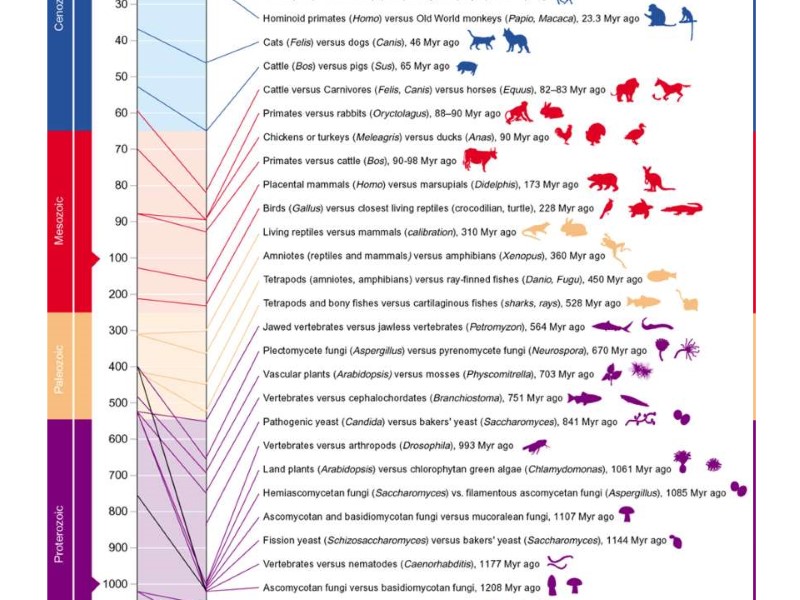
Рис. 8. Сравнение времени дивергенции основных групп органического








12398-prez_ant_temp.ppt
- Количество слайдов: 10
Рис. 8. Сравнение времени дивергенции основных групп органического мира по данным ископаемой летописи и молекулярных часов [117]. Масштаб шкалы времени меняется на границах интервалов 10-100, 100-1000 и 1000-5000 млн. лет. Конфликт данных палеонтологии и геномики заметно увеличивается по мере перехода к более ранним этапам биологической эволюции. Две древнейших датировки в столбце ископаемой летописи (слева) основаны на находках биомаркеров. Заключение Молекулярные часы обычно указывают более раннее время появления той или иной группы эвкариотных организмов в сравнении с известным возрастом древнейших окаменелостей соответствующей группы, который следует воспринимать как минимальный (рис. 8). Модели молекулярных часов, основанные на разных методиках, нередко противоречат не только ископаемой летописи, но и друг другу, однако диапазон расхождения результатов заметно уменьшается по мере роста количества полностью расшифрованных геномов и совершенствования методов. Особое значение в процедуре калибровки молекулярных часов имеет надежность интерпретации ископаемого материала, который должен включать не только окаменелости, но широкий спектр геохимических (в том числе, изотопных) и палеобиохимических сигналов, биоминералы и биолиты, другие объекты, формировавшиеся под влиянием биоты. Данные палеонтологии и геномики согласованно указывают на то, что бурной радиации эвкариот в середине неопротерозоя предшествовала их весьма длительная эволюция, детали которой нам пока что мало известны (возможно, в силу ограниченности первичных биотопов эвкариот в древней биосфере). Палеонтология и геномика не противоречат друг другу в том, что диверсификация многих таксонов высокого ранга из ныне живущих эвкариот происходила до неопротерозойских оледенений (рис. 7, 8). Факт выживания этих групп в течение ледниковых периодов (их дальнейшая эволюция в фанерозое) свидетельствует против крайних палеоклиматических моделей в виде гипотезы Snowball Earth [113, 114]. При этом невозможно отрицать существенного влияния физико-химических характеристик холодноводных биотопов неопротерозоя на дальнейшую эвкариотизацию экосистем [92, 115]. Определение времени основных этапов эвкариотизации биосферы, связанных с возникновением оксигенного фотосинтеза, становлением эвкариотной клетки и происхождением животных, имеет фундаментальное значение для постижения причинно-следственных связей в биологической эволюции, включая эволюцию генома. Прикладное значение этих исторических реконструкций видится в том, что с эвкариотизацией древней биосферы связаны радикальные изменения биогеохимических циклов и седиментогенеза (приведшие, в частности, к формированию характерных для протерозоя типов рудных скоплений железа, марганца, урана, фосфора и др., а также углеродистых толщ и углеводородов). Разрешение конфликта результатов, полученных методами молекулярных часов, видится не только в сравнении с палеонтологической летописью конкретных групп. Предстоит выяснить, как отражаются в эволюции генома крупные экосистемные перестройки, нередко связанные с важнейшими абиотическими изменениями в биосфере, насколько распознаваемы эти события в молекулярной летописи генома разных групп. Эта проблема требует синтеза данных наук о Земле, палеобиологии и геномики на системной (функциональной) основе, включающей геологическую историю не как сценическую декорацию «эволюционной драмы», а как органичную часть живой истории Земли.
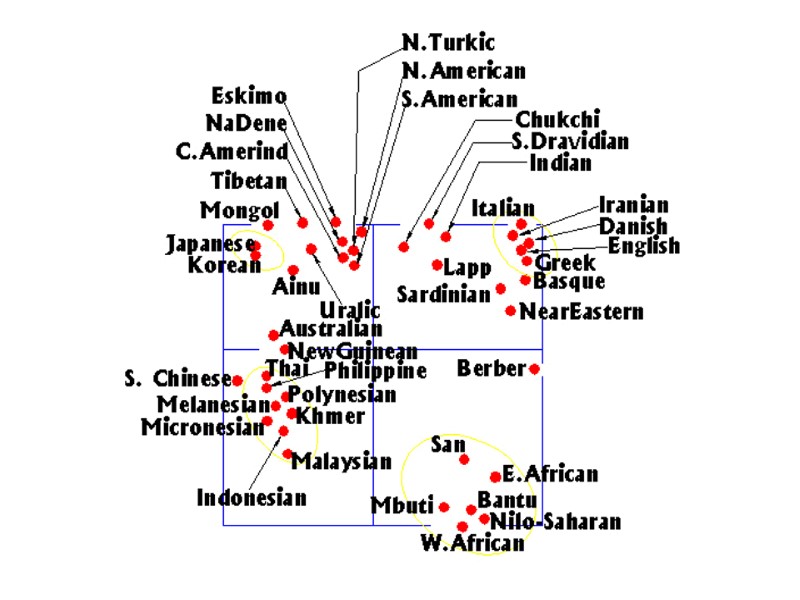
Genetic Distance Between Autochthonous Human Populations
Language Affinities Beween Autochthonous Populations
Language Affinities
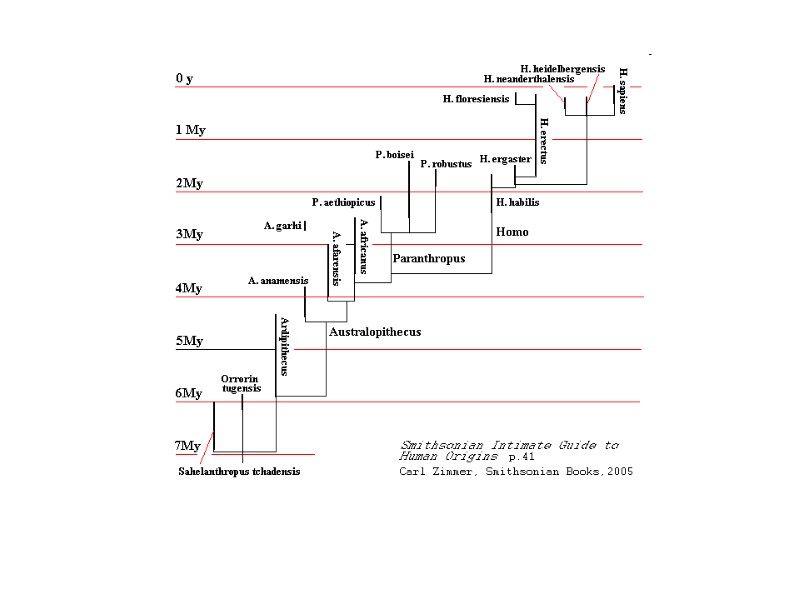
Human Evolution
The Semitic and Other Afroasiatic Languages