lecture_7.ppt
- Количество слайдов: 47




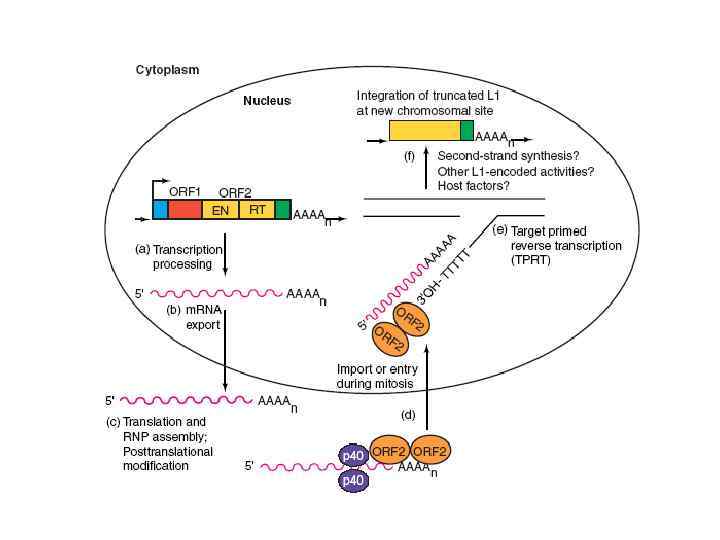
Ретроэлементы без длинных концевых повторов • В геноме человека есть несколько семейств LINE элементов; наиболее представлено семейство L 1. В геноме человека присутствует от 60, 000 до 100, 000 L 1 элементов. ORF 1 кодирует рнк-связывающий белок p 40. ORF 2 кодирует обратную транскриптазу и эндонуклеазу, которые необходимы для перемещения мобильных элементов. Красным цветом обозначены короткие прямые повторы, которые есть у всех мобильных элементов.

SINEs Short Interspersed Nuclear Elements Длина ~ 300 bp Не содержат функциональных генов (LINE элементы с делетированной внутренней частью) У человека наиболее известо семейство Alu элементов (от 700. 000 до 1. 000 Alu элементов на гаплоидный геном)

Перестройки иммуноглобулиновых генов VD J
J recombinase RAG 1 + RAG 2 (RAG")
RSS 23 RSS 12 RSS 23 V(D)J recombinase RAG 1 + RAG 2 (RAG – Recombination Activating Gene) Рекомбинация происходит только между разнонаправленными RSS 12 и RSS 23 При этом вырезается фрагмент ДНК, разделяющий RSS 12 и RSS 23
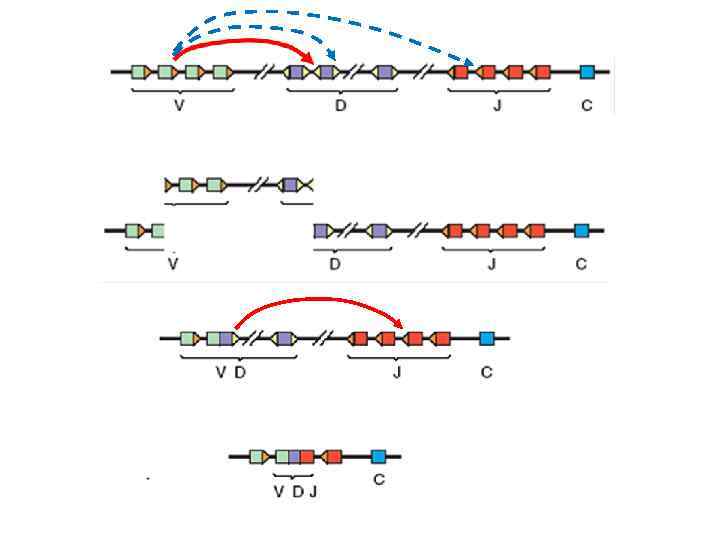
J Recombination D D")
Mechanism of V(D)J Recombination D D

ТРАНСКРИПЦИЯ

РНК полимераза E Coli была выделена в начале 60 х годов 20 в
![Dissociation of RNAP and purification of by ion-exchange chromatography ’ [Na. Cl] [protein] Carboxymethyl-](https://present5.com/presentation/5088598_148141310/image-11.jpg "Dissociation of RNAP and purification of by ion-exchange chromatography ’ [Na. Cl] [protein] Carboxymethyl-")
Dissociation of RNAP and purification of by ion-exchange chromatography ’ [Na. Cl] [protein] Carboxymethyl- (-CO 2 -2) or phospho- (-PO 3 -2) cellulose Fraction number ’
 ’ Core enzyme Может")
’ Holoenzyme Может транскрибировать интактную (не содержащую разрывов ДНК) ’ Core enzyme Может транскрибировать только ДНК, содержащую большое число однонитевых разрывов

Т 4 и большинство других фагов имеют три группы генов, которые транскрибируются на разных этапах репликационного цикла фага Immediate Early (2 мин) Работает только клеточный транскрипционный аппарат Delayed Early (2 -10 мин) нужен хотя бы один вирус-специфичные белок, так как экспрессия этих генов блокируется ингибиторами синтеза белка Late (10 – 25 мин) после начала синтеза фаговой ДНК IE IE + DE IE+DE+L
 специфически связывается с РНК полимеразу инкубировали с радиоактивно меченной. ДНК")
РНК полимераза II (холоэнзим) специфически связывается с РНК полимеразу инкубировали с радиоактивно меченной. ДНК фага T 7, определенными последовательностями ДНК после чего добавляли избыток немеченой ДНК и инкубировали в течение различных промежутков времени. Связанную с ДНК РНК полимеразу сорбировали на нитроцеллюлозных фильтрах ’ + Core enzyme неспецифическое связывание с ДНК (Kd ~5 x 10 -12 M) ’ Holoenzyme Специфическое связывание с опреленными последовательностями ДНК (Kd ~10 -7 M)

Стабильность специфических комплексов РНК полимеразы с ДНК увеличивается при повышении температуры локальная денатурация ДНК ?

Структура промоторов Е. Coli Pribnow box TATA box -10 box Сильные промоторы: хорошее соответствие консенсусу и правильный спейсинг Слабые промоторы: существенные отличия от консенсуса в -35 and -10 районах

сигма фактор определяет специфичность связывания РНК порлимеразы с промоторами Сравнение аминокислотных последовательностей ряда сигма факторов выявило наличие четырех консервативных участков участок 1 – подавление неспецифического связывания с ДНК участок 2 (наиболее консервативный) – связывание с -10 боксом промотора участок 3 - (факультативный) связывание с -10 боксом промотора участок 4 – связывание с -35 боксом промотора

сигма фактор узнаваемые промоторы 70 32 28 38 -35 бокс -10 бокс Большинство генов TTGACAT TATAAT Гены, активирующиеся при тепловом шоке TCTCNCCCTTGAA CCCCATNTA Гены подвижности и хемотаксиса CTAAA CCGATAT Гены стационарной фазы и ответа на стрессы ? ? 54 Гены метаболизма азота структура промотора -24 Region CTGGNA -12 Region TTGCA

The 3 D structure of bacterial RNA polymerase holoenzyme 3 s factor domains : N-term s 1 s 2 s 3 s 4 Inhibition -10 binding -35 binding

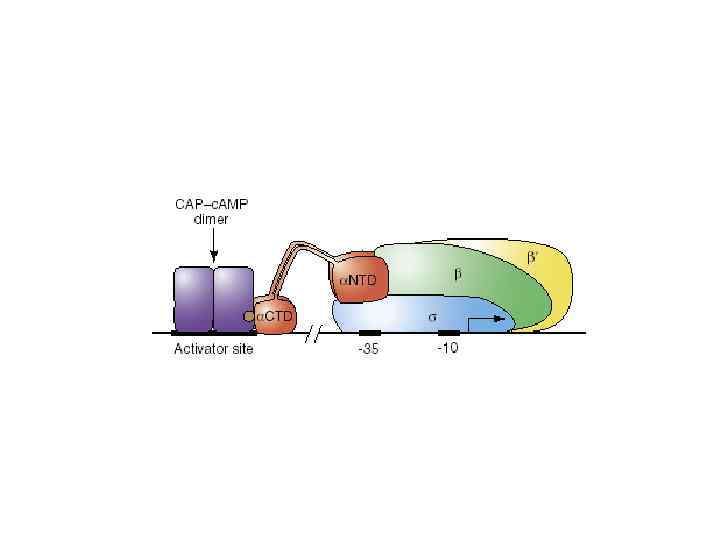
UP элемент является частью промотора, так как этот элемент стимулирует транскрипцию РНК полимеразой в отсутствие дополнительных факторов сигма-фактор не связывается с UP элементом ?

RNAP NTD CTD UP 4 -35 2 -3 -10 +1

РНК полимераза E. coli, связанная с ДНК субъединица стехиометрия в составе холофермента 2 ’ функция связывает регуляторные последовательности ДНК и белки Каталитический центр Узнавание промотора сборка РНК полимеразы
 2. Плавление ДНК (образование «открытого» комплекса)")
1. Связывание холофермента с промотором (образование «закрытого» комплекса) 2. Плавление ДНК (образование «открытого» комплекса) 3. Синтез короткой цепи РНК ( ~ 10 нуклеотидов) 4. Высвобождение сигма-фактора 5. Начало элонгации кор-фермент холофермент

локальная денатурация ДНК

РНК полимераза защищает от нуклеазной атаки фрагмент ДНК протяженностью ~ 46 п. н. (от -44 до +3)
 12 п. н. (от")
Размер денатурированного участка 10 п. н. гиперхромный эффект (открытый комплекс) 12 п. н. (от -9 до +3) DMS футпринтинг 17 п. н. подсчет числа супервитков в кольцевой ДНК SV 40 (открытый комплекс + элонгирующие комплексы) В ДНК SV 40 случайно присутствует промотор, который узнает сигма 70
 многократное использование сигма-фактора + кор-фермент устойчивый к рифампицину")
включение радиоактивной метки (количество инициаций транскрипции) многократное использование сигма-фактора + кор-фермент устойчивый к рифампицину + рифампицин транскрипция холоферментом в условиях, когда освобождение фермента по завершении транскрипции невозможно избыток матрицы время (мин)

Структура РНК полимеразы Thermus aquaticus сделана с разрешением 4 А Активный центр находится в углублении (ДНК канале) и содержит ион Mg++, связанный с тремя остатками аспартата в составе β′ субъединицы Структура активного центра похожа у разных РНК полимераз

Характерной особенностью РНК полимераз является наличие вторичного канала, через который транспортируются предшественники Нематричная цепь ДНК выпетливается из активного центра β’ DNA β RNA Secondary RNA exit channel Active site

Молекула ДНК изгибается внутри активного центра
")
Transcriptional elongation: Movement of transcription bubble (17 -bp, 1. 6 turns of B-DNA duplex) Speed of movement: 50 -90 -nt/sec Supercoiling of DNA during transcription causes a requirement for topoisomerases

Терминация транскрипции Существует Rho-зависимая и Rho-независимая терминация транскрипции Rho-независимые терминаторы состоят из последовательности, способной образовывать шпильку и Т-богатой последовательности

Терминация Предпологается, что движение РНК полимеразы замедляется на Тбогатой последовательности. Во время паузы формируется шпилька, которая дестабилизирует транскрипционный комплекс Терминация является вероятностным событием. Иногда полимераза «проскакивает» терминатор.

Зависящая от Rho терминация Rho состоит из 6 субъединиц, каждая из которых обладает ATФ-азной активностью Rho связывается с определенными Сбогатыми последовательностями в новосинтезированной РНК и начинает перемещаться, догоняя движущуюся полимеразу. Терминация происходит в тот момент, когда полимераза делает паузу на шпилечной структуре. Rho разрушает ДНК-РНК гибрид за счет присутствующей в его составе хеликазной активности Зависящие от Rho терминаторы не содержат Т-богатой последовательности

Lac оперон

индуктор

Работа lac-оперона в клетках, содержащих нормальный геном и частичную копию генома с мутациями в lac-опероне и регуляторных генах (merodiploids) В составе меродиплоида мутация не проявляется (индуцибельная экспрессия lac оперона Рецессивная Мутация Lac оперон работает в отсутствие лактозы есть репрессор, работающий in trans В составе меродиплоида конститутивная экспрессия lac оперона Доминантная Мутация Lac оперон работает в отсутствие лактозы есть cis-регуляторный элемент, мутации в котором препятствуют связыванию trans-репрессора

Нормальная копия оперона не работает на среде с лактозой Негативная доминантная мутация Репрессор состоит из нескольких субъединиц. Мутации, затрагивающие одну субъединицу, влияют на способность всего репрессора связываться с индуктором, либо с оператором
 в лактозном опероне есть два «вспомогательных» оператора (О")
Наряду с главным оператором (О 1) в лактозном опероне есть два «вспомогательных» оператора (О 2 и О 3). Уровень репрессии значительно снижается, если делетировать одновременно О 2 и О 3 тетрамер репрессора связывается одновременно с двумя участками узнавания на молекуле ДНК

Репресор не препятствует связыванию РНК полимеразы с промотором Репрессия осуществляется на этапе перехода от открытого комплекса к элонгации

Почему lac-оперон не работает, когда в среде есть и глюкоза и лактоза? Катаболитная репрессия ? В действительности - активация CAP – Catabolite Activator Protein (активатор катаболитных оперонов) Работает в комплексе с цикло-АМФ, уровень которого существенно повышается при недостатке глюкозы CAP активирует целый ряд оперонов, обеспечивающих синтез ферментов, необходимых для использования альтернативных продуктов питания (gal, галактозный оперон; ara, арабинозный оперон)

CAPстимулирует образование открытого комплекса Участок связывания CAP находится перед промотором и содержит TGTGA последовательность Lac промотор и многие другие катаболитные промоторы являются очень слабыми. -35 бокс в этих промоторах сильно отличается от консенсуса Существуют мутанты laс промотора, которые обеспечивают транскрипцию без САP активатора

Анаболитные опероны Должны активироваться в условиях недостатка тех веществ, в синтезе которых участвуют кодируемые опероном ферменты триптофановый оперон: триптофан (корепрессор) связывается с неактивной формой репрессора, превращая ее в активную
 обеспечивает дополнительную репрессию В начале оперона находится короткая ORF, кодирующая")
Преждевременная остановка транскрипции (attenuation) обеспечивает дополнительную репрессию В начале оперона находится короткая ORF, кодирующая так называемый лидерный пептид После этой рамки считывания находится сигнал терминации транскрипции 90% транскрипционных комплексов диссоциируют
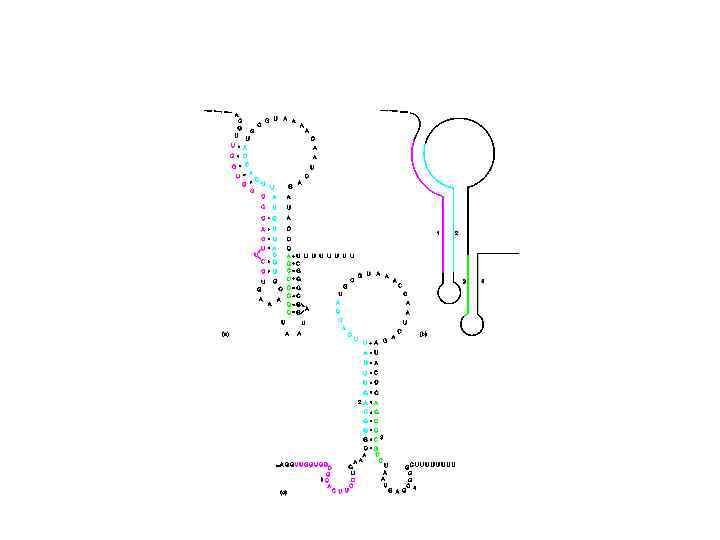

У прокариот транскрипция и трансляция могут происходить одновременно При отсутствии триптофана рибосома задерживается на лидерной последовательности. Это смещает равновесие в сторону большой шпильки Регулируемые терминаторы транскрипции есть и в других анаболитных оперонах В гистидиновом опероне лидер содержит 7 кодонов гистидина
lecture_7.ppt