Лекция 8 СК 2012 Репродуктивная гибель клеток.ppt
- Количество слайдов: 42



Репродуктивная гибель делящихся клеток.

Вопросы. • 1. Реакции клеток на облучение. • 2. Количественные характеристики гибели облученных клеток. • 3. Молекулярные механизмы радиационноиндуцированной репродуктивной гибели клеток.

Из истории: • В начале ХХ века Г. Хейнеке и Е. C. Лондон выявили мощное цитопеническое действие ИИ на организм. Возникло представление о лучевом поражении как “cиндроме клеточной гибели”. • В дальнейшем, начиная с работ Бергонье и Трибондо, это представление получило убедительные подтверждения. • К настоящему времени оно углублено обнаружением разновидностей и биохимических механизмов гибели клеток, а также дополнено сведениями о нелетальных клеточных реакциях на облучение.

1. Реакции клеток на облучение Преходящие реакции – физиологические или кумулятивные эффекты облучения: остановка клеточного деления – блокировка клеточного цикла. Летальные реакции: 1. Интерфазная гибель клеток. 2. Репродуктивная гибель клеток.
 клеточной гибели: • 1. Интерфазная (интеркинетическая) гибель: ü до деления (под лучом);")
Типы (формы) клеточной гибели: • 1. Интерфазная (интеркинетическая) гибель: ü до деления (под лучом); ü без деления. • 2. Митотическая (репродуктивная) гибель: ü после первого деления; ü в последующих делениях; ü гибель делящихся клеток популяции.

В цитоплазме после облучения происходит: • изменение вязкости: при малых дозах она снижается, при больших - повышается; • вакуолизация цитоплазмы (отчетливо просматривается у клеток крови, красного костного мозга, паренхиматозных органов); • повышение проницаемости мембран для электролитов и воды - из клетки выходит калий, а в ней накапливается натрий (у эритроцитов это приводит к гемолизу); • повышение коэффициента лучевого преломления, что, по-видимому, связано с денатурацией белков плазмы.

Изменения ядра в облученных клетках. • В облученных клетках наблюдается увеличение размеров ядер. • В дальнейшем изменяется форма ядра, появляются гигантские ядра, тройные звезды в митозе, трех- и четырехъядерные и гиперсегментированные клетки, некротические явления - пикноз и реже лизис ядра. • Подвижные клетки после облучения обычно принимают округлую форму.

Репродуктивная гибель • Особенности гибели быстроделящихся клеток подчеркивают термины, которые в разное время были приняты для ее обозначения: ”митотическая гибель”, ”отсроченная гибель”, ”гибель при делении”, ”репродуктивная гибель”.

Репродуктивная гибель • Последний термин получил наибольшее распространение, так как в качестве критерия гибели используют методы количественной оценки репродуктивной способности клеток (способность к образованию колоний in vitro, селезеночных колоний и другие методы оценки клоногенной активности).

Репродуктивная гибель • Репродуктивная гибель клеток заключается в необратимой потере способности к делению, облученными клетками или их потомками. Репродуктивная гибель клеток имеет место при относительно невысоких дозах облучения. • Этот радиобиологический эффект хорошо изучен количественными методами на различных типах клеток.

Репродуктивная гибель • Репродуктивная гибель делящихся клеток происходит не сразу после облучения, а постепенно, в процессе нескольких циклов деления. • Так показано, что при облучении культуры фибробластов мышей в дозе 4 Гр, около 80 % клеток заканчивали 1 деление, а второе и третье деление завершали только 30 % начавшихся делиться клеток. • Репродуктивная гибель клеток проявляется в образовании гигантских клеток в результате слияния нескольких клеток или при длительной задержке цитокинеза.

Репродуктивная гибель • Характерна для всех тканей организма, имеющих высокую митотическую активность. • Она присуща, в частности, молодым, малодифференцированным клеткам кроветворной системы и эпителия тонкой кишки - тканевых систем, лимитирующих выживание млекопитающих при облучении в дозах 1 -20 Гр. • Эта летальная реакция клеток на облучение вносит основной вклад в опустошение костного мозга в первые дни острой лучевой болезни.

Репродуктивная гибель • Репродуктивно погибающие клетки элиминируются из ткани и замещаются клетками, не имеющими хромосомных аберраций. • Поэтому стохастические летальные эффекты не могут быть причиной отдаленной гибели клеток в обновляющихся тканях. • Тем не менее частота последней бывает повышена даже спустя много лет после облучения, особенно в условиях нанесения дополнительного митогенного стимула.

«неклассическая» репродуктивная гибель • Одно из возможных объяснений этого явления (60 -е годы И. Б. Бычковская и соавт. ) - феномен нестохастических скрытых летальных повреждений клеток. Нестохастичность данного эффекта выражается в его появлении у 100% облученных клеток; его скрытый характер проявляется в нормальном функционировании клетки в ряде поколений до момента гибели при очередной попытке вступить в митоз. • В отличие от “классической” репродуктивной гибели, описываемая форма элиминации клеток наблюдается уже после облучения в дозах, на три порядка меньших.

«неклассическая» репродуктивная гибель • По-видимому, изменения, обусловливающие данную разновидность репродуктивной клеточной гибели, носят необратимый характер. Об этом свидетельствует неизменный в течение ряда лет процент превышения смертности потомков облученных клеток над контролем. • Установление конкретных молекулярных механизмов и морфологических субстратов данной разновидности репродуктивной клеточной гибели могло бы оказаться полезным в плане борьбы с поздними нарушениями иммунитета у лиц, подвергшихся радиационным воздействиям при авариях на АЭС.

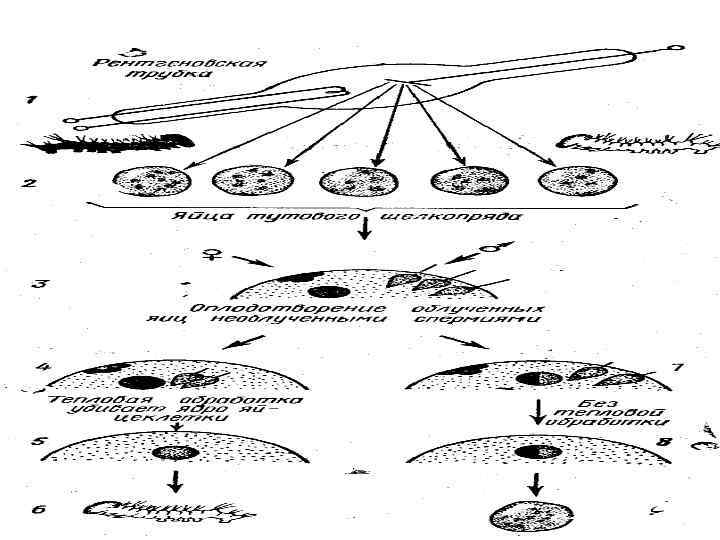
Экспериментальные доказательства роли ядра в репродуктивной гибели: • Опыты Астаурова – феномен андрогенеза – мужской партеногенез – возможность развития организма из мужской половой клетки без оплодотворения (тутовый шелкопряд).

2. Количественные характеристики гибели облученных клеток • Методы оценки жизнеспособности облученных клеток • Существует несколько методов оценки жизнеспособности облученных клеток, основанных на определении их способности к неограниченному размножению путем образования колоний или бляшек, возникающих из одиночных клеток.

Методы оценки жизнеспособности облученных клеток • Определение выживаемости клеток in vitro Количественный метод определения выживаемости клеток млекопитающих после облучения впервые был разработан в 1956 г. Т. Паком и П. Маркусом для культуры клеток He. La. И в настоящее время он является основным методом, применяемым в количественной радиобиологии. • Определение выживаемости клеток in vivo В 1961 году канадские ученые Дж. Тилл. и Е. Мак-Кулох предложили метод определения радиочувствительности стволовых клеток гемопоэтической ткани.

Метод Г. Пака и П. Маркуса на культуре клеток He. La. • Определенное количество культуральных клеток высевают на твердую питательную среду в чашках Петри, которые облучают различными дозами радиации и инкубируют в оптимальных условиях. • Через определенный промежуток времени подсчитывают число образовавшихся колоний на поверхности питательной среды. • Выживаемость клеток при определенной дозе облучения определяют как отношение числа колоний выросших в облученных чашках, к числу колоний в контрольной чашке Петри (без облучения).

Метод Дж. Тилла и Е. Мак-Кулоха • Клетки костного мозга, печени или селезенки вводят в вену летально облученных мышей. Через неделю на селезенке мышей появляются колонии, состоящие из потомства введенных клеток. Число колоний, образующихся на селезенке, показывает пролиферативную (клоногенную) способность введенных клеток, которая зависит от дозы облучения вводимых клеток. • Путем сравнения числа колоний, полученных от облученных в различных дозах клеток, и числа колоний на селезенках контрольных мышей (с клетками без облучения), получают кривые выживания.

Кривые выживаемости доза – эффект: А – линейные координаты, Б – полулогарифмические координаты.

n")
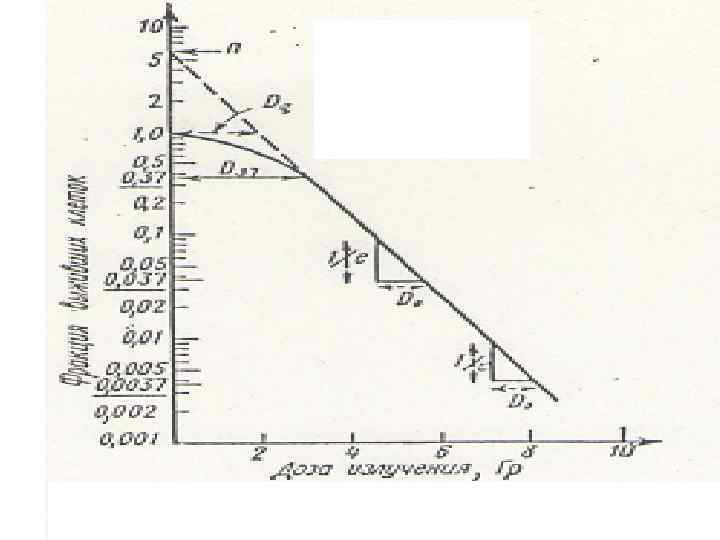
Кривые выживания описываются уравнением N/ N 0 = 1 - (1 - e-D/D 0)n • где n - экстраполяционное число, которое определяется как значение ординаты в месте ее пересечения с экстраполированным прямолинейным участком кривой выживания. • D 0 - приращение дозы, снижающее выживаемость клеток в е раз на прямолинейном участке кривой выживания.

Зависимость доза-эффект носит экспоненциальный характер: N=N 0 e –D/D 0, где N – число выживших после облучения клеток из общего их числа, N 0 – исходное число клеток, D – любая доза излучения, D 0 - доза излучения, при которой выживает 37 клеток: N/N 0 = e-1 = 0, 367

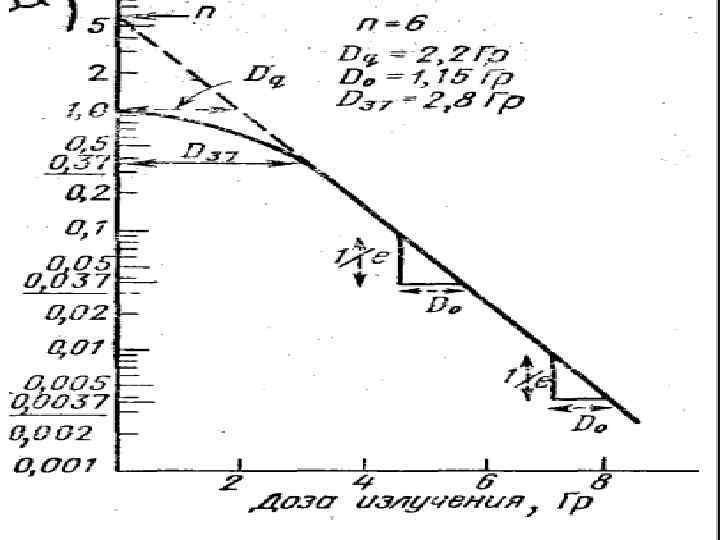
Кривые «доза — эффект» с «плечом» . • n — экстраполяционное число, определяемое как значение ординаты в месте ее пересечения с экстраполированным прямолинейным участком кривой выживаемости, • Do — определяет исходную радиочувствительность и вычисляется как приращение дозы, снижающей выживаемость в е раз на прямолинейном участке кривой «доза — эффект» . • Dq — «квазипороговая доза» - характеризует работу систем репарации.

Мерой способности клеток к репарации является величина плеча, оцениваемая квазипороговой дозой Dq. • Эта доза соответствует точке пересечения экстраполированного прямолинейного участка кривой выживания с прямой, параллельной оси абсцисс, проведенной на уровне 100% выживаемости. • При уменьшении способности клеток к репарации величина n уменьшается до 1; а величина Dq до 0 Гр.
 погибает 37")
Дозы D 37, D 0, Dq • При дозе D 37 (среднелетальной) погибает 37 % облученных клеток. • D 0, Dq - характеризуют регенерационную способность клеток, которая в конечном счете характеризует устойчивость клеток к облучению. Чем выше значение Dq, чем длиннее плечо, тем выше способность клеток посрадиационному восстановлению и следовательно, их устойчивость к облучению.

Дозы D 37, D 0 • При дозе D 0 выживает 36, 7% и погибает 63, 3%. Величина D 0 служит показателем радиочувствительности клеток и определяется по кривой выживания как доза, при которой выживает 37% клеток от исходного количества. • Поэтому эту величину иногда называют D 37, что справедливо в случае экспоненциальных кривых, тогда как в случае кривых, имеющих плечо, D 0 и D 37 различны.

3. Молекулярные механизмы репродуктивной гибели клеток. • Морфологической основой инактивации репродуктивной способности клеток служат хромосомные аберрации - фрагменты, внутрихромосомные и межхромосомные обмены. • Существует тесная связь между частотой возникновения хромосомных аберраций и клеточной гибелью. • Молекулярным субстратом хромосомных аберраций являются нерепарированные двутяжевые разрывы ДНК.

Основная причина репродуктивной гибели клеток - повреждение ДНК. Типы лучевых повреждений ДНК: • одиночные и двойные разрывы (именно они ответственны за летальный исход облучения); • повреждение азотистых оснований нуклеотидов с последующим их удалением из ДНК; • внутри- и межмолекулярные сшивки типа ДНКДНК, ДНК-белок;

Основные типы повреждения ДНК
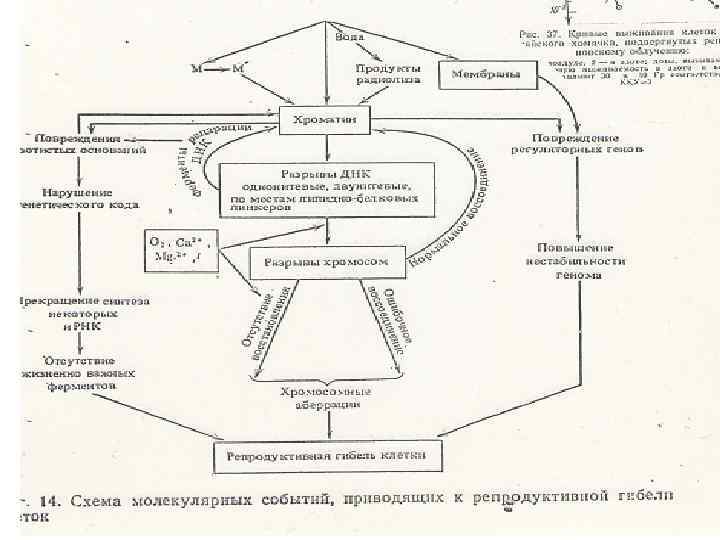

Схема возникновения аберраций хромосомного типа

Схема возникновения аберраций хроматидного типа
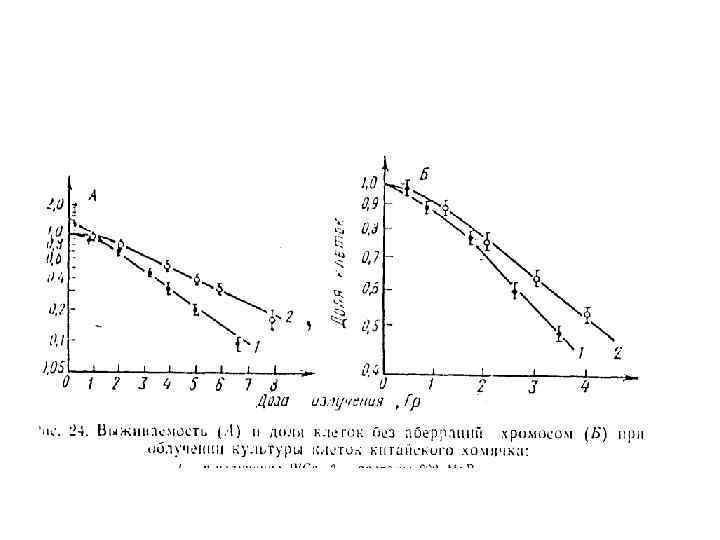
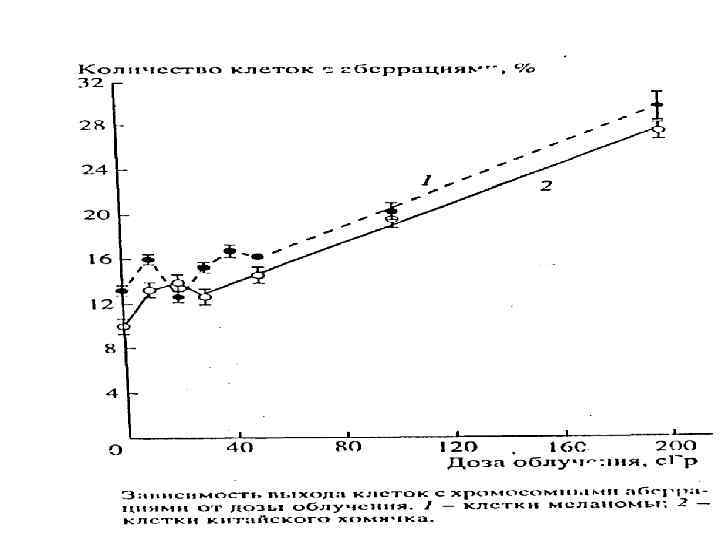

Модификация репродуктивной гибели


Лекция 8 СК 2012 Репродуктивная гибель клеток.ppt