През_ант_syst.ppt
- Количество слайдов: 44
 Рекомендуемая литература Харитонов В. М. , Ожигова А. П. , Година Е. З. , Хрисанфова Е. Н. , Бацевич В. А. Антропология: Учебн. Пособие для студ. Высш. Учеб. Заведений. – М. : Гуччанит. Изд. центр ВЛАДОС, 2004. – 272 с. Хрисанфова Е. Н. , Перевозчиков И. В. Антропология. М. 1991. 319 с
Рекомендуемая литература Харитонов В. М. , Ожигова А. П. , Година Е. З. , Хрисанфова Е. Н. , Бацевич В. А. Антропология: Учебн. Пособие для студ. Высш. Учеб. Заведений. – М. : Гуччанит. Изд. центр ВЛАДОС, 2004. – 272 с. Хрисанфова Е. Н. , Перевозчиков И. В. Антропология. М. 1991. 319 с
 Дополнительная литература Алексеева Т. И. Адаптивные процессы в популяциях человека. М. : Изд во Моск. ун та, 1986. 138 с. Ван Гудолл, Дж. Лавик. В тени человека. М. , 1974. Джохансон Д. , Иди. Люси: Истоки рода человеческого. М. : Мир, 1984. 295 с. Дольник В. Р. Непослушное дитя биосферы. СПб. , 1994. Докинс Р. Эгоистичный ген. М. , 1993. Новоженов Ю. И. Филетическая эволюция человека. Свердловск: Изд во Свердловского педин та, 1983. 85 с. Фоули Р. Еще один неповторимый вид/ Экологические аспекты эволюции человека. М. : Мир, 1990, 367 с. Харрисон Дж. , Уайнер Дж. , Тэннер Дж. , и др. Биология Человека. М. : Мир, 1979. 438 с.
Дополнительная литература Алексеева Т. И. Адаптивные процессы в популяциях человека. М. : Изд во Моск. ун та, 1986. 138 с. Ван Гудолл, Дж. Лавик. В тени человека. М. , 1974. Джохансон Д. , Иди. Люси: Истоки рода человеческого. М. : Мир, 1984. 295 с. Дольник В. Р. Непослушное дитя биосферы. СПб. , 1994. Докинс Р. Эгоистичный ген. М. , 1993. Новоженов Ю. И. Филетическая эволюция человека. Свердловск: Изд во Свердловского педин та, 1983. 85 с. Фоули Р. Еще один неповторимый вид/ Экологические аспекты эволюции человека. М. : Мир, 1990, 367 с. Харрисон Дж. , Уайнер Дж. , Тэннер Дж. , и др. Биология Человека. М. : Мир, 1979. 438 с.
 • Веб-ресурсы • Природа и древний человек: http: //dino. disneyjazz. net/dr. html • Проблемы эволюции (антропогенез): http: //macroevolution. narod. ru/; http: //www. evolbiol. ru/index. html • Антропология (электронный учебник): http: //www. ido. edu. ru/psychology/anthropology/index. html • http: //imp. rudn. ru/psychology/anthropology/index. html • Достающее звено. http: //antropogenez. ru/zveno/
• Веб-ресурсы • Природа и древний человек: http: //dino. disneyjazz. net/dr. html • Проблемы эволюции (антропогенез): http: //macroevolution. narod. ru/; http: //www. evolbiol. ru/index. html • Антропология (электронный учебник): http: //www. ido. edu. ru/psychology/anthropology/index. html • http: //imp. rudn. ru/psychology/anthropology/index. html • Достающее звено. http: //antropogenez. ru/zveno/

 КОГОРТА ARCHONTA • Морфолого палеонтологические данные свидетельствуют в пользу монофилетического статуса всей группы архонт, включающей 4 отряда; молекулярно генетические данные подтверждают монофилию группы Euarchonta (включает Scandentia, Primates и Dermoptera). Согласно молекулярно генетическим данным, входящие сюда отряды относятся к Boreoeutheria. В некоторых схемах отсюда исключаются Primates.
КОГОРТА ARCHONTA • Морфолого палеонтологические данные свидетельствуют в пользу монофилетического статуса всей группы архонт, включающей 4 отряда; молекулярно генетические данные подтверждают монофилию группы Euarchonta (включает Scandentia, Primates и Dermoptera). Согласно молекулярно генетическим данным, входящие сюда отряды относятся к Boreoeutheria. В некоторых схемах отсюда исключаются Primates.
 ОТРЯД ШЕРСТОКРЫЛЫ — DERMOPTERA • Шерстокрылы, кагуаны или летающие лемуры (отряд Dermoptera или подотряд Dermoptera отряда Primates) известны с нижнего и среднего палеогена Северной Америки, Европы и Азии, где были представлены множеством видов, но сейчас сохранилось только два вида, которые живут на юге Индокитая, полуострове Малакка, Зондских островах и Филиппинах. Шерстокрылы имеют довольно своеобразный вид, поскольку между их передними и задними ногами вдоль тела до хвоста натянута кожная перепонка, позволяющая им планировать с деревьев, наподобие летяг. В связи с такой адаптацией конечности шерстокрылов длинные и тонкие, а локтевая кость редуцирована и сращена с лучевой. Кроме этого, зубная формула шерстокрылов примитивна, а шесть нижних резцов имеют особое щётковидное строение. Своеобразной чертой шерстокрылов является наличие сосков под мышками. Хотя по некоторым генетическим маркёрам шерстокрылы ближе к приматам, чем тупайи, их специализация к планирующему полёту показывает, что морфологически они ушли дальше от общего предка всех трёх групп.
ОТРЯД ШЕРСТОКРЫЛЫ — DERMOPTERA • Шерстокрылы, кагуаны или летающие лемуры (отряд Dermoptera или подотряд Dermoptera отряда Primates) известны с нижнего и среднего палеогена Северной Америки, Европы и Азии, где были представлены множеством видов, но сейчас сохранилось только два вида, которые живут на юге Индокитая, полуострове Малакка, Зондских островах и Филиппинах. Шерстокрылы имеют довольно своеобразный вид, поскольку между их передними и задними ногами вдоль тела до хвоста натянута кожная перепонка, позволяющая им планировать с деревьев, наподобие летяг. В связи с такой адаптацией конечности шерстокрылов длинные и тонкие, а локтевая кость редуцирована и сращена с лучевой. Кроме этого, зубная формула шерстокрылов примитивна, а шесть нижних резцов имеют особое щётковидное строение. Своеобразной чертой шерстокрылов является наличие сосков под мышками. Хотя по некоторым генетическим маркёрам шерстокрылы ближе к приматам, чем тупайи, их специализация к планирующему полёту показывает, что морфологически они ушли дальше от общего предка всех трёх групп.

 известны") ОТРЯД ТУПАЙИ — SCANDENTIA • Тупайи (отряд Scandentia или подотряд Tupaiiformes отряда Primates) известны с эоцена Европы (Nycticonodon caparti), а сейчас обитают в Юго Восточной Азии – от северной Индии через Индокитай до Индонезии и Филиппин. Внешне эти зверьки очень похожи на белок, отчего раньше их относили к отряду насекомоядных. Сейчас существует пять родов и множество видов, среди которых есть полудревесные и полуназемные, дневные и сумеречные, растительно и насекомоядные. По сравнению с приматами, тупайи имеют очень примитивную морфологию и, видимо, по облику и образу жизни близки к древнейшим предкам приматов – плезиадаписовым. • В отличие от приматов, у тупай на пальцах имеются когти, большой палец не противопоставлен остальным, мордочка сильно вытянута вперёд, а глаза расположены по бокам головы. Зубная формула тупай примитивна, на нижней челюсти имеется шесть резцов.
ОТРЯД ТУПАЙИ — SCANDENTIA • Тупайи (отряд Scandentia или подотряд Tupaiiformes отряда Primates) известны с эоцена Европы (Nycticonodon caparti), а сейчас обитают в Юго Восточной Азии – от северной Индии через Индокитай до Индонезии и Филиппин. Внешне эти зверьки очень похожи на белок, отчего раньше их относили к отряду насекомоядных. Сейчас существует пять родов и множество видов, среди которых есть полудревесные и полуназемные, дневные и сумеречные, растительно и насекомоядные. По сравнению с приматами, тупайи имеют очень примитивную морфологию и, видимо, по облику и образу жизни близки к древнейшим предкам приматов – плезиадаписовым. • В отличие от приматов, у тупай на пальцах имеются когти, большой палец не противопоставлен остальным, мордочка сильно вытянута вперёд, а глаза расположены по бокам головы. Зубная формула тупай примитивна, на нижней челюсти имеется шесть резцов.

 ОТРЯД ПРИМАТЫ — PRIMATES • Вероятно, монофилетический таксон, входит в состав Euarchonta; в некоторых системах сближается с Rodentia. Не включает Tupaiidae; иногда сюда относят Dermoptera в ранге подотряда. Филогенетическая структура не вполне ясна: обычно выделяют 2– 3 подотряда, несколько инфраотрядов. Количество и состав семейств также требуют уточнения: в настоя щее время их признаётся от 10 до 16 современных и почти 20 ископаемых. С раннего палеогена. Тропики и субтропики Африки (вкл. Мадагаскар), Азии, Центр. и Юж. Америки.
ОТРЯД ПРИМАТЫ — PRIMATES • Вероятно, монофилетический таксон, входит в состав Euarchonta; в некоторых системах сближается с Rodentia. Не включает Tupaiidae; иногда сюда относят Dermoptera в ранге подотряда. Филогенетическая структура не вполне ясна: обычно выделяют 2– 3 подотряда, несколько инфраотрядов. Количество и состав семейств также требуют уточнения: в настоя щее время их признаётся от 10 до 16 современных и почти 20 ископаемых. С раннего палеогена. Тропики и субтропики Африки (вкл. Мадагаскар), Азии, Центр. и Юж. Америки.
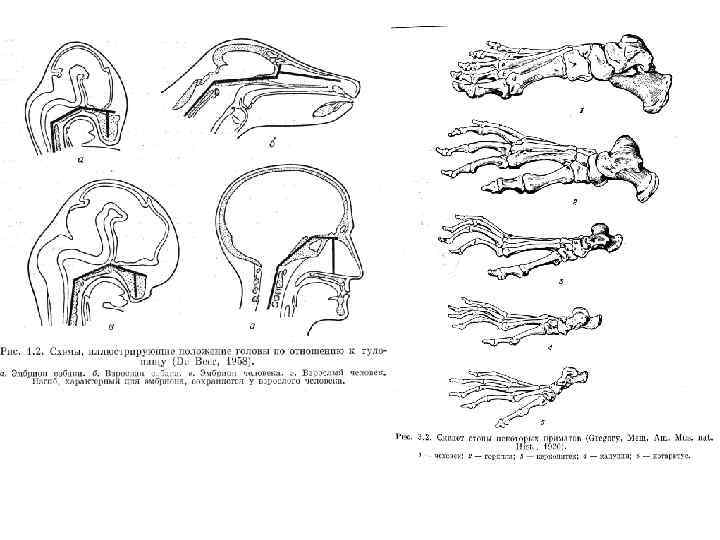
 • Приматы отличаются от других млекопитающих следующими признаками: • относительно большие размеры мозга при редукции обонятельных центров; • пять пальцев на всех конечностях (у специализированных видов большой или указательный (иногда и средний) палец могут редуцироваться, а прочие – срастаться); • способность конечностей и особенно запястья к вращению; • ногти на пальцах (у полуобезьян не отдельных пальцах остаются когти); • глаза повёрнуты вперёд, а глазница имеет тенденцию к закрытию сзади (полностью только у высших приматов); • преимущественно древесный образ жизни; • питание насекомыми (в основном у полуобезьян) и растительной пищей, чаще та или иная степень всеядности; • малая плодовитость и отсутствие сезонности размножения (не резко выраженная сезонность есть у полуобезьян); • жизнь группами, достаточно сложные социальные отношения и системы коммуникации, сложное, разнообразное и лабильное поведение.
• Приматы отличаются от других млекопитающих следующими признаками: • относительно большие размеры мозга при редукции обонятельных центров; • пять пальцев на всех конечностях (у специализированных видов большой или указательный (иногда и средний) палец могут редуцироваться, а прочие – срастаться); • способность конечностей и особенно запястья к вращению; • ногти на пальцах (у полуобезьян не отдельных пальцах остаются когти); • глаза повёрнуты вперёд, а глазница имеет тенденцию к закрытию сзади (полностью только у высших приматов); • преимущественно древесный образ жизни; • питание насекомыми (в основном у полуобезьян) и растительной пищей, чаще та или иная степень всеядности; • малая плодовитость и отсутствие сезонности размножения (не резко выраженная сезонность есть у полуобезьян); • жизнь группами, достаточно сложные социальные отношения и системы коммуникации, сложное, разнообразное и лабильное поведение.
 ПОДОТРЯД STREPSIRRHINI • Вероятно, монофилетический таксон; в концепции Prosimii сюда включают также Tarsiidae. 3 надсемейства (иногда трактуются как инфраотряды), 5– 6 семейств. • Полуобезьяны (подотряд Prosimii или Strepsirhini) включают лемуроподобных полуобезьян (Lemuriformes) и множество ископаемых родов и видов. Главные отличия полуобезьян от высших приматов заключаются в следующем:
ПОДОТРЯД STREPSIRRHINI • Вероятно, монофилетический таксон; в концепции Prosimii сюда включают также Tarsiidae. 3 надсемейства (иногда трактуются как инфраотряды), 5– 6 семейств. • Полуобезьяны (подотряд Prosimii или Strepsirhini) включают лемуроподобных полуобезьян (Lemuriformes) и множество ископаемых родов и видов. Главные отличия полуобезьян от высших приматов заключаются в следующем:
 • • • • • малые размеры тела; сравнительно небольшой мозг без извилин и крупная, выступающая мордочка; сравнительно хорошее развитие обоняния, большой нос сливается с голой верхней губой, образуя ринариум; имеются вибриссы – осязательные волоски на мордочке, есть подшёрсток; глазницы развёрнуты косо, а не строго вперёд, глазницы сзади не закрыты, хотя вокруг глазниц имеется костное кольцо; имеются слуховые капсулы, а наружного слухового прохода нет; половины нижней челюсти не срастаются, есть отросток в углу нижней челюсти; резцы и клыки на нижней челюсти образуют выступающую вперёд "щётку" или "гребёнку" для расчёсывания шерсти (кроме руконожки); имеется "подъязык" – вырост под языком с зубчатым краем, служащий для чистки описанной выше "зубной гребёнки"; на втором пальце стопы имеется коготь, используемый для расчёсывания шерсти; мимика не развита, воду лакают языком, а не пьют; кольца трахеи замкнутые, лёгкие и печень многодольчатые; в конечностях имеются "чудесные сплетения" – диффузные артериальные и венозные сети; матка двурогая, плацента диффузная с неотпадающей слизистой оболочкой матки; может быть пара паховых молочных желёз в дополнение к грудным или взамен грудных; чаще ночной или сумеречный образ жизни, отчего на сетчатке глаза нет жёлтого пятна; сезонное размножение. Кроме более примитивной морфологии, полуобезьяны отличаются от настоящих обезьян поведением. Это почти исключительно ночные или сумеречные животные, питаются как растительностью, так и насекомыми. Некоторые виды ведут одиночный образ жизни, некоторые живут группами.
• • • • • малые размеры тела; сравнительно небольшой мозг без извилин и крупная, выступающая мордочка; сравнительно хорошее развитие обоняния, большой нос сливается с голой верхней губой, образуя ринариум; имеются вибриссы – осязательные волоски на мордочке, есть подшёрсток; глазницы развёрнуты косо, а не строго вперёд, глазницы сзади не закрыты, хотя вокруг глазниц имеется костное кольцо; имеются слуховые капсулы, а наружного слухового прохода нет; половины нижней челюсти не срастаются, есть отросток в углу нижней челюсти; резцы и клыки на нижней челюсти образуют выступающую вперёд "щётку" или "гребёнку" для расчёсывания шерсти (кроме руконожки); имеется "подъязык" – вырост под языком с зубчатым краем, служащий для чистки описанной выше "зубной гребёнки"; на втором пальце стопы имеется коготь, используемый для расчёсывания шерсти; мимика не развита, воду лакают языком, а не пьют; кольца трахеи замкнутые, лёгкие и печень многодольчатые; в конечностях имеются "чудесные сплетения" – диффузные артериальные и венозные сети; матка двурогая, плацента диффузная с неотпадающей слизистой оболочкой матки; может быть пара паховых молочных желёз в дополнение к грудным или взамен грудных; чаще ночной или сумеречный образ жизни, отчего на сетчатке глаза нет жёлтого пятна; сезонное размножение. Кроме более примитивной морфологии, полуобезьяны отличаются от настоящих обезьян поведением. Это почти исключительно ночные или сумеречные животные, питаются как растительностью, так и насекомыми. Некоторые виды ведут одиночный образ жизни, некоторые живут группами.
 НАДСЕМЕЙСТВО LEMUROIDEA s. lato. В некоторых системах вместе с Daubentoniidae выделяется в ин фраотряд Lemuriformes. 1 вымершее (3 подсемейства и более 30 родов, Африка, Азия, Европа) и 3 современных (Мадагаскар) семейства
НАДСЕМЕЙСТВО LEMUROIDEA s. lato. В некоторых системах вместе с Daubentoniidae выделяется в ин фраотряд Lemuriformes. 1 вымершее (3 подсемейства и более 30 родов, Африка, Азия, Европа) и 3 современных (Мадагаскар) семейства
 СЕМЕЙСТВО ИНДРИЕВЫЕ – INDRIIDAE BURNETT, 1828 Возможно, заслуживает выделения в отдельное надсемейство. 2 подсемейства, 9 родов (6 вымерли в историческое время). Леса Мадагаскара.
СЕМЕЙСТВО ИНДРИЕВЫЕ – INDRIIDAE BURNETT, 1828 Возможно, заслуживает выделения в отдельное надсемейство. 2 подсемейства, 9 родов (6 вымерли в историческое время). Леса Мадагаскара.
 НАДСЕМЕЙСТВО DAUBENTONIOIDEA s. lato В некоторых системах сближается с Lemuroidea; иногда рассматривается как инфраотряд Chiromyiformes. СЕМЕЙСТВО РУКОНОЖКОВЫЕ – DAUBENTONIIDAE GRAY, 1863 1 род. Лесные (преимущественно прибрежные) районы Мадагаскара.
НАДСЕМЕЙСТВО DAUBENTONIOIDEA s. lato В некоторых системах сближается с Lemuroidea; иногда рассматривается как инфраотряд Chiromyiformes. СЕМЕЙСТВО РУКОНОЖКОВЫЕ – DAUBENTONIIDAE GRAY, 1863 1 род. Лесные (преимущественно прибрежные) районы Мадагаскара.
 НАДСЕМЕЙСТВО LOROIDEA s. lato В некоторых системах рассматривается как инфраотряд Loriformes. 2 семейства (ранее объединялись в одно); иногда сюда включают также Cheirogaleidae. СЕМЕЙСТВО ЛОРИЕВЫЕ – LORIDAE GRAY, 1821 = Lorisidae auct. С раннего неогена. Лесные об ласти ндо Малайского региона и Центр. И Африки. СЕМЕЙСТВО ГАЛАГОВЫЕ – GALAGONIDAE GRAY, 1825 = Galagidae auct. 3 рода. С поздн. палеогена. Лесные области (вкл. саванные леса) Африки к югу от Сахары.
НАДСЕМЕЙСТВО LOROIDEA s. lato В некоторых системах рассматривается как инфраотряд Loriformes. 2 семейства (ранее объединялись в одно); иногда сюда включают также Cheirogaleidae. СЕМЕЙСТВО ЛОРИЕВЫЕ – LORIDAE GRAY, 1821 = Lorisidae auct. С раннего неогена. Лесные об ласти ндо Малайского региона и Центр. И Африки. СЕМЕЙСТВО ГАЛАГОВЫЕ – GALAGONIDAE GRAY, 1825 = Galagidae auct. 3 рода. С поздн. палеогена. Лесные области (вкл. саванные леса) Африки к югу от Сахары.
 ПОДОТРЯД HAPLORHINI Имеются доводы в пользу как монофилии, так и парафилии. Делится на 2– 3 инфраотряда. • Обезьяны, высшие приматы, антропоиды или человекоподобные (Anthropoidea или Haplorrhini) – подразделяются на три большие группы: долгопятовых, широконосых и узконосых. Отделение долгопятовых произошло прежде прочих, в среднем эоцене или раньше, благодаря чему они имеют множество черт строения полуобезьян. Разделение на широконосых и узконосых произошло в позднем эоцене или олигоцене при миграции части обезьян в Южную Америку.
ПОДОТРЯД HAPLORHINI Имеются доводы в пользу как монофилии, так и парафилии. Делится на 2– 3 инфраотряда. • Обезьяны, высшие приматы, антропоиды или человекоподобные (Anthropoidea или Haplorrhini) – подразделяются на три большие группы: долгопятовых, широконосых и узконосых. Отделение долгопятовых произошло прежде прочих, в среднем эоцене или раньше, благодаря чему они имеют множество черт строения полуобезьян. Разделение на широконосых и узконосых произошло в позднем эоцене или олигоцене при миграции части обезьян в Южную Америку.
 • • • • • Обезьяны в целом имеют следующие главные признаки: большие размеры тела; сравнительно большой мозг с извилинами (нет извилин у долгопятовых, но и у них обонятельный мозг редуцирован); плохое развитие обоняния, нос отделён от обволошенной верхней губы; нет вибрисс (есть у долгопятовых) и обычно нет подшёрстка; глазницы развёрнуты строго вперёд, глазницы сзади закрыты, отделены от височной ямки (у широконосых в стенке есть большое глазнично височное отверстие); нет слуховых капсул у узконосых, есть у долгопятовых и широконосых, наружный слуховой проход замкнутый (у широконосых короткий, со щелью); половины нижней челюсти срастаются, нет отростка в углу нижней челюсти, угол округлый; резцы и клыки на нижней челюсти отличаются друг от друга, не образуют "зубной гребёнки"; нет "подъязыка"; на всех пальцах ногти (у игрунковых когтевидные); мимика хорошо развита, воду пьют (долгопятовые лакают языком); кольца трахеи полулунные, незамкнутые, лёгкие и печень немногодольчатые (у долгопятовых лёгкие и печень многодольчатые); в конечностях нет "чудесных сплетений" (есть у долгопятовых); матка простая (двурогая у долгопятовых), плацента дисковидная с отпадающей слизистой оболочкой матки; только две грудные молочные железы (у долгопятовых пара грудных и пара паховых); дневной образ жизни (кроме долгопятовых и некоторых ночных и сумеречных широконосых обезьян), на сетчатке глаза есть жёлтое пятно (нет у долгопятовых); нет сезонности размножения.
• • • • • Обезьяны в целом имеют следующие главные признаки: большие размеры тела; сравнительно большой мозг с извилинами (нет извилин у долгопятовых, но и у них обонятельный мозг редуцирован); плохое развитие обоняния, нос отделён от обволошенной верхней губы; нет вибрисс (есть у долгопятовых) и обычно нет подшёрстка; глазницы развёрнуты строго вперёд, глазницы сзади закрыты, отделены от височной ямки (у широконосых в стенке есть большое глазнично височное отверстие); нет слуховых капсул у узконосых, есть у долгопятовых и широконосых, наружный слуховой проход замкнутый (у широконосых короткий, со щелью); половины нижней челюсти срастаются, нет отростка в углу нижней челюсти, угол округлый; резцы и клыки на нижней челюсти отличаются друг от друга, не образуют "зубной гребёнки"; нет "подъязыка"; на всех пальцах ногти (у игрунковых когтевидные); мимика хорошо развита, воду пьют (долгопятовые лакают языком); кольца трахеи полулунные, незамкнутые, лёгкие и печень немногодольчатые (у долгопятовых лёгкие и печень многодольчатые); в конечностях нет "чудесных сплетений" (есть у долгопятовых); матка простая (двурогая у долгопятовых), плацента дисковидная с отпадающей слизистой оболочкой матки; только две грудные молочные железы (у долгопятовых пара грудных и пара паховых); дневной образ жизни (кроме долгопятовых и некоторых ночных и сумеречных широконосых обезьян), на сетчатке глаза есть жёлтое пятно (нет у долгопятовых); нет сезонности размножения.
 ИНФРАОТРЯД TARSIIFORMES Родственные связи не вполне ясны: обычно сближается с Platyrrhini, реже с Strepsirrhini; в виде ис ключения — с Dermoptera. Ранн. и средн. палеоген Сев. Америки, с ранн. палеогена в Старом Свете. 4 вымерших и 1 современное семейства. • • СЕМЕЙСТВО ДОЛГОПЯТОВЫЕ – TARSIIDAE GRAY, 1825 1 род. В поздн. палеогене и ранн. неогене в Юго Вост. Азии. В настояще время — Большие Зондские о ва (кроме Явы), южная часть Филиппин, острова между ними. Современные долгопяты (Tarsiidae) живут в Индонезии и на Филиппинах. Это животные размером с крысу, с очень большими глазами и специализированными для прыгания ногами. Они активны ночью и питаются насекомыми. Во многих чертах строения проявляется резкая специализация: глаза столь велики, что не могут вращаться, отчего подвижность шеи резко увеличена, она может поворачиваться задом наперёд; мордочка крайне маленькая; на нижней челюсти только два резца; кончики пальцев расширены в виде присосок, большая и малая берцовые кости срастаются, пятка резко удлинена в виде цевки для прыгания по ветвям, а на втором и третьем пальцах стопы находятся когти. Живут долгопяты семейными группами, общаясь друг с другом своеобразным щебетом, похожим на птичий.
ИНФРАОТРЯД TARSIIFORMES Родственные связи не вполне ясны: обычно сближается с Platyrrhini, реже с Strepsirrhini; в виде ис ключения — с Dermoptera. Ранн. и средн. палеоген Сев. Америки, с ранн. палеогена в Старом Свете. 4 вымерших и 1 современное семейства. • • СЕМЕЙСТВО ДОЛГОПЯТОВЫЕ – TARSIIDAE GRAY, 1825 1 род. В поздн. палеогене и ранн. неогене в Юго Вост. Азии. В настояще время — Большие Зондские о ва (кроме Явы), южная часть Филиппин, острова между ними. Современные долгопяты (Tarsiidae) живут в Индонезии и на Филиппинах. Это животные размером с крысу, с очень большими глазами и специализированными для прыгания ногами. Они активны ночью и питаются насекомыми. Во многих чертах строения проявляется резкая специализация: глаза столь велики, что не могут вращаться, отчего подвижность шеи резко увеличена, она может поворачиваться задом наперёд; мордочка крайне маленькая; на нижней челюсти только два резца; кончики пальцев расширены в виде присосок, большая и малая берцовые кости срастаются, пятка резко удлинена в виде цевки для прыгания по ветвям, а на втором и третьем пальцах стопы находятся когти. Живут долгопяты семейными группами, общаясь друг с другом своеобразным щебетом, похожим на птичий.
 ИНФРАОТРЯД PLATYRRHINI = Simiiformes • . Монофилетический таксон (что подчеркивается объединением входящих сюда семейств в единственное надсемейство Ceboidea). • В некоторых системах считается сестринской группой для Catarrhini, объединяется вместе с ними в группу Anthropoidea. В классических и некоторых кладистических системах признаются 2 семейства, в наиболее дробных молекулярно генетических от 5 до 7 семейств; в настоящей системе — 3 семейства. Тропические и субтропические леса Юж. и Центр. Америки.
ИНФРАОТРЯД PLATYRRHINI = Simiiformes • . Монофилетический таксон (что подчеркивается объединением входящих сюда семейств в единственное надсемейство Ceboidea). • В некоторых системах считается сестринской группой для Catarrhini, объединяется вместе с ними в группу Anthropoidea. В классических и некоторых кладистических системах признаются 2 семейства, в наиболее дробных молекулярно генетических от 5 до 7 семейств; в настоящей системе — 3 семейства. Тропические и субтропические леса Юж. и Центр. Америки.
 • • • СЕМЕЙСТВО ИГРУНКОВЫЕ – CALLITHRICHIDIAE GRAY, 1821 2 подсемейства (иногда считаются семействами), 4 рода. С ранн. палеогена. Дождевые тропи ческие, листопадные и саванные леса Юж. Америки (Амазонский ре гион, Бразильское нагорье) и Центр. Америки. СЕМЕЙСТВО КАПУЦИНОВЫЕ – CEBIDAE BONAPARTE, 1831 Возможно, сборная группа, часть подсемейств которой следует включать в Atelidae. В при нятой здесь трактовке — 4 подсемейства, 5 современных (1 вымер в историческое время) и 5 вымерших родов. С ранн. палеогена. Тропические и субтропиче ские леса ж. и Центр. Ю Америки; острова Карибского бассейна. СЕМЕЙСТВО КОАТОВЫЕ – ATELIDAE Gray, 1825 Саковые. Возможно, сюда относится часть подсемейств из Cebidae. 2 подсемейства (иногда рас сматриваются как семейства), 7– 8 родов; 1 подсемейство и более 5 родов — ископаемые. С ранн. палеогена. Равнинные—горные тропические леса Юж. и Центр. Америки; в средн. неогене также острова Карибского бассейна.
• • • СЕМЕЙСТВО ИГРУНКОВЫЕ – CALLITHRICHIDIAE GRAY, 1821 2 подсемейства (иногда считаются семействами), 4 рода. С ранн. палеогена. Дождевые тропи ческие, листопадные и саванные леса Юж. Америки (Амазонский ре гион, Бразильское нагорье) и Центр. Америки. СЕМЕЙСТВО КАПУЦИНОВЫЕ – CEBIDAE BONAPARTE, 1831 Возможно, сборная группа, часть подсемейств которой следует включать в Atelidae. В при нятой здесь трактовке — 4 подсемейства, 5 современных (1 вымер в историческое время) и 5 вымерших родов. С ранн. палеогена. Тропические и субтропиче ские леса ж. и Центр. Ю Америки; острова Карибского бассейна. СЕМЕЙСТВО КОАТОВЫЕ – ATELIDAE Gray, 1825 Саковые. Возможно, сюда относится часть подсемейств из Cebidae. 2 подсемейства (иногда рас сматриваются как семейства), 7– 8 родов; 1 подсемейство и более 5 родов — ископаемые. С ранн. палеогена. Равнинные—горные тропические леса Юж. и Центр. Америки; в средн. неогене также острова Карибского бассейна.


 ИНФРАОТРЯД CATARRHINI Монофилетический таксон, чаще всего считается сестринской группой для Platyrrhini, вместе с которым обычно объединяется в группу Anthropoidea (реже прямое родство между ними отвергается). 2 надсемейства, 3 семейства (плюс 1 вымершее). Старый Свет. • • НАДСЕМЕЙСТВО CERCOPITHECOIDEA s. lato СЕМЕЙСТВО МАРТЫШКОВЫЕ – CERCOPITHECIDAE GRAY, 1821 Монофилетический таксон с 2 современными и 1 ископаемым подсемействами. На родовом уровне сис тема неустойчива: количество признаваемых родов от 11 до 21 (в ископаемом состоянии около 15 родов). С ранн. неогена. Африка к югу и востоку от Сахары, Юж. , Юго Вост. (вкл. Малайский арх. ) и Вост. (на север до Японии) Азия; крайний юго запад Европы (акклиматизация).
ИНФРАОТРЯД CATARRHINI Монофилетический таксон, чаще всего считается сестринской группой для Platyrrhini, вместе с которым обычно объединяется в группу Anthropoidea (реже прямое родство между ними отвергается). 2 надсемейства, 3 семейства (плюс 1 вымершее). Старый Свет. • • НАДСЕМЕЙСТВО CERCOPITHECOIDEA s. lato СЕМЕЙСТВО МАРТЫШКОВЫЕ – CERCOPITHECIDAE GRAY, 1821 Монофилетический таксон с 2 современными и 1 ископаемым подсемействами. На родовом уровне сис тема неустойчива: количество признаваемых родов от 11 до 21 (в ископаемом состоянии около 15 родов). С ранн. неогена. Африка к югу и востоку от Сахары, Юж. , Юго Вост. (вкл. Малайский арх. ) и Вост. (на север до Японии) Азия; крайний юго запад Европы (акклиматизация).
 Мартышкообразные • • Мелкие виды мартышкообразных – мартышки и мангобеи – обычно сугубо древесные формы, средние – макаки – одинаково ловко бегают и по земле, и по ветвям, а крупные – мандрилы и павианы – наземные. Они заняли достаточно широкий спектр экологических ниш, но все являются дневными, стадными и растительноядными. Среди бесчисленных видов мартышек выделяются мартышки гусары, приспособившиеся к жизни в саванне, где они бегают подобно мелким антилопам. У мангобеев хвост имеет очень слабую хватательную способность – уникальный случай среди узконосых приматов; хотя мангобеи ведут древесный образ жизни, они наиболее близки павианам. Макаки – самые холодоустойчивые обезьяны (кроме человека), особенно выделяются в этом отношении тибетские и японские макаки, каждую зиму сталкивающиеся со морозом и снегом. Наиболее устрашающе выглядят целебесские макаки или "чёрные павианы". Мандрилы и павианы – крупные животные с клыками как у леопарда, очень сильные и агрессивные. Среди них особенно цветасты мандрилы с их ярко раскрашенной мордой Выделются и гелады – эфиопские павианы – в группах которых главенствуют самки. Мартышка мона (Cercopithecus mona)
Мартышкообразные • • Мелкие виды мартышкообразных – мартышки и мангобеи – обычно сугубо древесные формы, средние – макаки – одинаково ловко бегают и по земле, и по ветвям, а крупные – мандрилы и павианы – наземные. Они заняли достаточно широкий спектр экологических ниш, но все являются дневными, стадными и растительноядными. Среди бесчисленных видов мартышек выделяются мартышки гусары, приспособившиеся к жизни в саванне, где они бегают подобно мелким антилопам. У мангобеев хвост имеет очень слабую хватательную способность – уникальный случай среди узконосых приматов; хотя мангобеи ведут древесный образ жизни, они наиболее близки павианам. Макаки – самые холодоустойчивые обезьяны (кроме человека), особенно выделяются в этом отношении тибетские и японские макаки, каждую зиму сталкивающиеся со морозом и снегом. Наиболее устрашающе выглядят целебесские макаки или "чёрные павианы". Мандрилы и павианы – крупные животные с клыками как у леопарда, очень сильные и агрессивные. Среди них особенно цветасты мандрилы с их ярко раскрашенной мордой Выделются и гелады – эфиопские павианы – в группах которых главенствуют самки. Мартышка мона (Cercopithecus mona)
 Тонкотелые • • • Тонкотелые – все исключительно древесные – специализированы к питанию листьями, что привело к усложнению их пищеварительного тракта – желудок имеет три отдела. В связи с сильной географической расчленённостью Юго Восточной Азии и Индонезии разнообразие тонкотелых крайне велико. До сих пор среди приматологов нет согласия относительно числа не только видов, но и родов тонкотелых. Лангуры послужили прототипом бандерлогов Р. Киплинга. Хохлатые лангуры имеют высокий острый хохол на голове, отчего несколько напоминают хохлатых мангобеев. Обезьяны носачи имеют запоминающуюся внешность: у самцов нос огромный, свисающий вниз, а у самок остренький и курносый. Африканские колобусы менее разнообразны, но не менее эффектны, особенно чёрно белые гверецы, имеющие при общем чёрном фоне длинную белую шерсть в виде мантии на плечах и огромной кисти на хвосте. Лангур Франсуа (Presbytis francoisi)
Тонкотелые • • • Тонкотелые – все исключительно древесные – специализированы к питанию листьями, что привело к усложнению их пищеварительного тракта – желудок имеет три отдела. В связи с сильной географической расчленённостью Юго Восточной Азии и Индонезии разнообразие тонкотелых крайне велико. До сих пор среди приматологов нет согласия относительно числа не только видов, но и родов тонкотелых. Лангуры послужили прототипом бандерлогов Р. Киплинга. Хохлатые лангуры имеют высокий острый хохол на голове, отчего несколько напоминают хохлатых мангобеев. Обезьяны носачи имеют запоминающуюся внешность: у самцов нос огромный, свисающий вниз, а у самок остренький и курносый. Африканские колобусы менее разнообразны, но не менее эффектны, особенно чёрно белые гверецы, имеющие при общем чёрном фоне длинную белую шерсть в виде мантии на плечах и огромной кисти на хвосте. Лангур Франсуа (Presbytis francoisi)
 павианы Наземные павианы представляют вариант жёстко построенного иерархического общества, где каждая особь кому либо подчинена и кого либо подчиняет. Иерархия основана на физической силе и жизненном опыте. Всё дозволено лишь самым маленьким детёнышам и вожакам стада. При переселениях стадо павианов представляет собой образец построения группы на марше. В центре стада движутся самки с детёнышами, возглавляет шествие вожак, а по сторонам охраняют самцы. Жёсткая иерархическая система держится на самцах вожаках. Интересно, что почти все нововведения и инициативы рождаются в "низах общества" – среди павианов самого низшего ранга, чаще среди самок, которые не так заняты поддержанием своего социального статуса. Тот же эффект ярко был продемонстрирован в наблюдениях над группой японских макак, которых исследователи подкармливали бататами. Когда одна самка низшего ранга случайно уронила батат в воду, она обнаружила, что чистым его гораздо приятнее есть. Она стала регулярно мыть клубни, от неё научился её детёныш, а после нововведение распрастранилось "от низов к верхам". Раньше часто павианов предлагали в качестве модели первобытного человеческого стада. Однако закреплённость и регламентированность взаимоотношений среди членов такого "общества" не дает возможности развития. Поэтому маловероятно, что отношения среди древнейших людей были похожи на отношения среди павианов. Мандрил
павианы Наземные павианы представляют вариант жёстко построенного иерархического общества, где каждая особь кому либо подчинена и кого либо подчиняет. Иерархия основана на физической силе и жизненном опыте. Всё дозволено лишь самым маленьким детёнышам и вожакам стада. При переселениях стадо павианов представляет собой образец построения группы на марше. В центре стада движутся самки с детёнышами, возглавляет шествие вожак, а по сторонам охраняют самцы. Жёсткая иерархическая система держится на самцах вожаках. Интересно, что почти все нововведения и инициативы рождаются в "низах общества" – среди павианов самого низшего ранга, чаще среди самок, которые не так заняты поддержанием своего социального статуса. Тот же эффект ярко был продемонстрирован в наблюдениях над группой японских макак, которых исследователи подкармливали бататами. Когда одна самка низшего ранга случайно уронила батат в воду, она обнаружила, что чистым его гораздо приятнее есть. Она стала регулярно мыть клубни, от неё научился её детёныш, а после нововведение распрастранилось "от низов к верхам". Раньше часто павианов предлагали в качестве модели первобытного человеческого стада. Однако закреплённость и регламентированность взаимоотношений среди членов такого "общества" не дает возможности развития. Поэтому маловероятно, что отношения среди древнейших людей были похожи на отношения среди павианов. Мандрил
") Гамадрил (Papio hamadryas)
Гамадрил (Papio hamadryas)
") Японский макак (Macaca fuscata)
Японский макак (Macaca fuscata)
 НАДСЕМЕЙСТВО HOMINOIDEA s. lato Монофилетический таксон. 2 семейства • • • • • • СЕМЕЙСТВО ГИББОНОВЫЕ – HYLOBATIDAE GRAY, 1871 Сестринская группа для Hominidae, иногда рассматривается как подсе мейство в его составе (или в составе Pongidae). В традиционных системах признаётся 1 род; в наиболее дробной кладистической выделяются 4 рода, группируемые в 2 подсемейства. С ранн. неогена. Тро пические листвен ные леса Индокитая, крайнего юга Юньнаня, на Больших Зондских о вах (кроме Сулавеси). Род Сиаманги – Symphalangus Gloger, 1841 1 вид. П ов Малакка, о. Суматра. syndactylus Raffles, 1821. Распространение — как указано для рода. Род Хулоки – Bunopithecus Matthew et Granger, 1923 1 вид. Сев. Зап. Индокитай. hoolock Harlan, 1834. Распространение — как указано для рода. Род Гиббоны – Hylobates Illiger, 1811 5– 7 видов. Юг Индокитая, п ов Малакка, арх. Ментавай, о ва Ява, Суматра, Борнео. klossi Miller, 1903. Арх. Ментавай. pileatus Gray, 1861. Юг Индокитая. moloch Audebert, 1798. Западная часть о. Ява. agilis Cuvier, 1821. П ов Малакка, центр и юг Суматры. ? albibarbis Lyon, 1911. Юго запад Борнео ? mulleri Martin, 1841. О. Борнео (кроме юго западной части). lar Linnaeus, 1771. Индокитай, Юж. Юньнань, п ов Малакка, северная часть Суматры. Род Номаскусы – Nomascus Miller, 1933 Наиболее близок к Hylobates (возможно, его подрод). 3– 5 видов. Индоки тай. concolor Harlan, 1826. Сев. Индокитай. leucogenys Ogilby, 1840. Сев. Индокитай. hainanus Thomas, 1892. Сев. Индокитай. ? gabriellae Thomas, 1909 (? siki Delacour, 1951). Юго Вост. Индокитай.
НАДСЕМЕЙСТВО HOMINOIDEA s. lato Монофилетический таксон. 2 семейства • • • • • • СЕМЕЙСТВО ГИББОНОВЫЕ – HYLOBATIDAE GRAY, 1871 Сестринская группа для Hominidae, иногда рассматривается как подсе мейство в его составе (или в составе Pongidae). В традиционных системах признаётся 1 род; в наиболее дробной кладистической выделяются 4 рода, группируемые в 2 подсемейства. С ранн. неогена. Тро пические листвен ные леса Индокитая, крайнего юга Юньнаня, на Больших Зондских о вах (кроме Сулавеси). Род Сиаманги – Symphalangus Gloger, 1841 1 вид. П ов Малакка, о. Суматра. syndactylus Raffles, 1821. Распространение — как указано для рода. Род Хулоки – Bunopithecus Matthew et Granger, 1923 1 вид. Сев. Зап. Индокитай. hoolock Harlan, 1834. Распространение — как указано для рода. Род Гиббоны – Hylobates Illiger, 1811 5– 7 видов. Юг Индокитая, п ов Малакка, арх. Ментавай, о ва Ява, Суматра, Борнео. klossi Miller, 1903. Арх. Ментавай. pileatus Gray, 1861. Юг Индокитая. moloch Audebert, 1798. Западная часть о. Ява. agilis Cuvier, 1821. П ов Малакка, центр и юг Суматры. ? albibarbis Lyon, 1911. Юго запад Борнео ? mulleri Martin, 1841. О. Борнео (кроме юго западной части). lar Linnaeus, 1771. Индокитай, Юж. Юньнань, п ов Малакка, северная часть Суматры. Род Номаскусы – Nomascus Miller, 1933 Наиболее близок к Hylobates (возможно, его подрод). 3– 5 видов. Индоки тай. concolor Harlan, 1826. Сев. Индокитай. leucogenys Ogilby, 1840. Сев. Индокитай. hainanus Thomas, 1892. Сев. Индокитай. ? gabriellae Thomas, 1909 (? siki Delacour, 1951). Юго Вост. Индокитай.
 • • Гиббоны замечательны своим способом локомоции – брахиацией, то есть передвижением только с помощью рук. Гиббон раскачивается на ветке на руках, перебрасывается на другую ветку, а ноги при этом никак не участвуют. Причём скорость быстрой брахиации практически равна скорости полёта птицы. Специализация к брахиации у гиббонов так велика, что они не могут передвигаться по земле на четырёх ногах и при необходимости спуститься на землю ходят на двух, поднимая руки вверх. Самый большой гиббон – сиаманг – отличается срастанием пальцев на руках, отчего образуется крючок, удобный при брахиации; за такое строение рук сиамангов называют ещё симфалангами. У других гиббонов может почти полностью редуцироваться большой палец на руке. Гиббоны уникальны своими вокальными способностями – они поют по утрам на вершинах деревьев, у некоторых видов – особенно у сиаманга – для этого есть даже увеличенные горловые мешки. Живут гиббоны семейными группами. В отличие от большинства приматов, у некоорых видов гиббонов самцы, самки и детёныши имеют разную окраску.
• • Гиббоны замечательны своим способом локомоции – брахиацией, то есть передвижением только с помощью рук. Гиббон раскачивается на ветке на руках, перебрасывается на другую ветку, а ноги при этом никак не участвуют. Причём скорость быстрой брахиации практически равна скорости полёта птицы. Специализация к брахиации у гиббонов так велика, что они не могут передвигаться по земле на четырёх ногах и при необходимости спуститься на землю ходят на двух, поднимая руки вверх. Самый большой гиббон – сиаманг – отличается срастанием пальцев на руках, отчего образуется крючок, удобный при брахиации; за такое строение рук сиамангов называют ещё симфалангами. У других гиббонов может почти полностью редуцироваться большой палец на руке. Гиббоны уникальны своими вокальными способностями – они поют по утрам на вершинах деревьев, у некоторых видов – особенно у сиаманга – для этого есть даже увеличенные горловые мешки. Живут гиббоны семейными группами. В отличие от большинства приматов, у некоорых видов гиббонов самцы, самки и детёныши имеют разную окраску.


,") СЕМЕЙСТВО ГОМИНИДЫ – HOMINIDAE GRAY, 1825 Монофилетический таксон. 2 подсемейства (иногда рассматриваются как семейства), 4 современных рода и около 15 вымерших. С поздн. палеогена в Азии, Европе и Африке. В настоящее время Экватор. Африка и Большие Зондские о ва; 1 вид всесветно. • • • • Подсемейство Ponginae Elliot, 1913 В традиционных системах часто рассматривается как семейство, включающее всех челове кообразных обезьян (в том числе ylobatidae). 1 род. Большие Зондские о ва. H Род Орангутаны – Pongo Lacepede, 1799 1 вид. Тропические леса на Суматре, Борнео (реликтовый ареал). pygmaeus Linnaeus, 1760 (? abeli Lesson, 1827). Распространение — как указано для рода. Подсемейство Homininae S. STR. Род Гориллы – Gorilla Geoffroy, 1852 1 вид. Тропические леса Центр. Африки. gorilla Savage, 1847 (? beringei Matschie, 1904). Распространение — как указано для рода. Род Шимпанзе – Pan Oken, 1816 Наиболее близок к Homo. 2 вида. Тропические и саванные равнинные—горные (до 3000 м) леса Экватор. Африки. troglodytes Blumenbach, 1775. Распространение — как указано для рода. paniscus Schwartz, 1929. Центр. Африка. Род Люди – Homo Linnaeus, 1758 Наиболее близок к Pan. 1 вид. Всесветно (вкл. околоземной Космос). sapiens Linnaeus, 1758. Распространение — как указано для рода.
СЕМЕЙСТВО ГОМИНИДЫ – HOMINIDAE GRAY, 1825 Монофилетический таксон. 2 подсемейства (иногда рассматриваются как семейства), 4 современных рода и около 15 вымерших. С поздн. палеогена в Азии, Европе и Африке. В настоящее время Экватор. Африка и Большие Зондские о ва; 1 вид всесветно. • • • • Подсемейство Ponginae Elliot, 1913 В традиционных системах часто рассматривается как семейство, включающее всех челове кообразных обезьян (в том числе ylobatidae). 1 род. Большие Зондские о ва. H Род Орангутаны – Pongo Lacepede, 1799 1 вид. Тропические леса на Суматре, Борнео (реликтовый ареал). pygmaeus Linnaeus, 1760 (? abeli Lesson, 1827). Распространение — как указано для рода. Подсемейство Homininae S. STR. Род Гориллы – Gorilla Geoffroy, 1852 1 вид. Тропические леса Центр. Африки. gorilla Savage, 1847 (? beringei Matschie, 1904). Распространение — как указано для рода. Род Шимпанзе – Pan Oken, 1816 Наиболее близок к Homo. 2 вида. Тропические и саванные равнинные—горные (до 3000 м) леса Экватор. Африки. troglodytes Blumenbach, 1775. Распространение — как указано для рода. paniscus Schwartz, 1929. Центр. Африка. Род Люди – Homo Linnaeus, 1758 Наиболее близок к Pan. 1 вид. Всесветно (вкл. околоземной Космос). sapiens Linnaeus, 1758. Распространение — как указано для рода.
 Род Орангутаны – Pongo Lacepede, 1799
Род Орангутаны – Pongo Lacepede, 1799
 Род Гориллы – Gorilla Geoffroy, 1852
Род Гориллы – Gorilla Geoffroy, 1852
 Род Шимпанзе – Pan Oken, 1816
Род Шимпанзе – Pan Oken, 1816


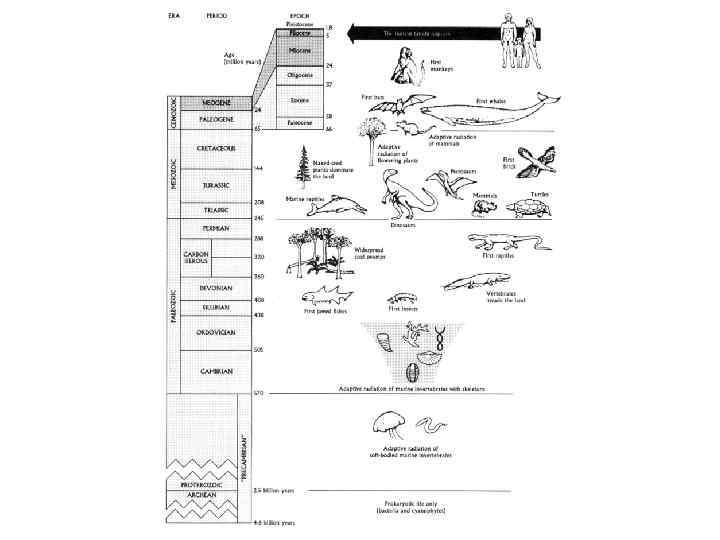
 • Антропогенез – раздел антропологии, изучающий происхождение человека. • Иначе, антропогенез – раздел антропологии, изучающий изменчивость человека во времени.
• Антропогенез – раздел антропологии, изучающий происхождение человека. • Иначе, антропогенез – раздел антропологии, изучающий изменчивость человека во времени.
 • • • • • Антропогенез – мультидисциплинарная наука. Комплекс подходов к изучению прошлого человечества включает массу подходов. Биологические науки: биология человека – анатомия, морфология, физиология, церебрология, палеонтология человека, антропометрия; приматология – палеонтология приматов, современная приматология, этология приматов; палеонтология – палеонтология позвоночных, палеоботаника, палинология; общая биология – эмбриология, генетика, молекулярная биология, сравнительная анатомия, сравнительная этология, биометрия. Физико-химико-математические науки: геология – геоморфология, геофизика, стратиграфия, геохронология; химия; тафономия (наука о захоронении ископаемых остатков); методы датирования – абсолютные: торий урановый, радиоуглеродный, термолюминесцентный, относительные: стратиграфический, сравнительно флористический и сравнительно фаунистический. статистика; Социальные науки: археология – археология палеолита, археология позднейших времен; этноархеология; сравнительная этнология (этнография); лингвистика; психология.
• • • • • Антропогенез – мультидисциплинарная наука. Комплекс подходов к изучению прошлого человечества включает массу подходов. Биологические науки: биология человека – анатомия, морфология, физиология, церебрология, палеонтология человека, антропометрия; приматология – палеонтология приматов, современная приматология, этология приматов; палеонтология – палеонтология позвоночных, палеоботаника, палинология; общая биология – эмбриология, генетика, молекулярная биология, сравнительная анатомия, сравнительная этология, биометрия. Физико-химико-математические науки: геология – геоморфология, геофизика, стратиграфия, геохронология; химия; тафономия (наука о захоронении ископаемых остатков); методы датирования – абсолютные: торий урановый, радиоуглеродный, термолюминесцентный, относительные: стратиграфический, сравнительно флористический и сравнительно фаунистический. статистика; Социальные науки: археология – археология палеолита, археология позднейших времен; этноархеология; сравнительная этнология (этнография); лингвистика; психология.