Регуляция обмена веществ (2).ppt
- Количество слайдов: 69



Регуляция обмена веществ

Уровни регуляции Внутриклеточная регуляция Нервная регуляция Гормональная регуляция
")
Внутриклеточная регуляция Регуляторы – метаболиты, которые образуются в самой клетке. Действуют 3 способами: 1) ингибируют и активируют ферменты; 2) изменяют количество молекул ферментов путем индукции или репрессии их синтеза; 3) изменяют активность транспортных белков мембраны → изменяется скорость трансмембранного переноса веществ.

Нервная регуляция Регуляторы – физические, химические, психические факторы → сигналы, вызывающие развитие нервных импульсов → высвобождение медиатора → действие на постсинаптическую мембрану, которая расположена на клетке-мишени → эффект.

Гормональная регуляция Регуляторы: гормоны – вещества, которые синтезируются и секретируются специальными железами, высвобождаются в ответ на стимул (нервный импульс, изменение концентрации веществ)
 Возможна регуляция обмена веществ в органах или клетках, удаленных")
Гормоны обладают следующими признаками: 1) Возможна регуляция обмена веществ в органах или клетках, удаленных от клеток – продуцентов – дистанционное действие. 2) Строгая специфичность эффекта. 3) Высокая биологическая активность.
 к клетке-мишени Синтез сигнальной молекулы Транспорт сигнальной")
Этапы передачи сигналов от сигнальных молекул (гормонов) к клетке-мишени Синтез сигнальной молекулы Транспорт сигнальной молекулы Связывание сигнальной молекулы с рецептором клеткимишени Активация рецептора и передача сигнала внутриклеточной молекуле-эффектору (фермент, транспортный белок, белок-регулятор) Активация (ингибирование) внутриклеточной молекулыэффектора Изменение метаболизма клетки-мишени Инактивация сигнальной молекулы
 Эндокринный - передача сигнала путем переноса гормона в кровотоке")
Пути передачи сигнала клетке 1) Эндокринный - передача сигнала путем переноса гормона в кровотоке от места синтеза до клеток мишеней. 2) Паракринный – действие на близлежащие клетки (клетки-мишени расположены близко к секреторным клеткам – тканевые гормоны). 3) Аутокринный – сигнальные вещества продуцируются и утилизируются в самих клетках (простагландины). 4) Юкстакринный – адгезия клеток другу (тромбоциты к эндотелию).

Пути передачи сигнала клетке клетка-мишень рецептор сигнальная молекула клетка-продуцент

Иерархия сигнальных молекул Сигнальные молекулы Микромолекулы Эйкозаноиды: простагландины, тромбоксаны, лейкотриены Представители О 2. NO ПГ Е 2, ПГ I 2 и др. ТХА 2 и др. Lt C 4 b др. Основные продуценты Путь передачи сигнала Клетки эпителия, эндотелиоциты АК ПК Лейкоциты, тромбоциты и др. ПК
 Многие клетки АК ПК ЭК интерлейкины")
Цитокины: факторы роста Васкулярный эндотелиальный фактор роста (VEGF) Многие клетки АК ПК ЭК интерлейкины Интерлейкин 1 -17 Лейкоциты АК ПК ЭК хемокины Адгезивные молекулы (RANTES и др. ) Клетки эпителия, эндотелия ЮК ПК факторы роста и некроза опухолей Фактор некроза опухолей альфа Лейкоциты ПК ЭК колониестимулирующие факторы Моноцитколониестимулирующий фактор и др. Лейкоциты ПК

Гистогормоны Гистамин, серотонин, гастрин, натрийуретический пептид и др. Гормоны: Эндокринные клетки в разных тканях ПК Эндокринны ЭК е железы белки, пептиды инсулин, антидиуретический гормон производные АМК катехоламины гормоны щитовидной железы стероиды половые гормоны (андрогены, эстрогены, прогестерон), минералокортикоиды,

Основные эффекты сигнальных молекул Микромолекулы – регулируют процессы в разных тканях: сосудистый тонус (вазоконстрикция, вазодилатация) пролиферацию, дифференцировку клеток апоптоз в любых тканях нейромедиаторы
: медиаторы воспаления агрегация тромбоцитов факторы хемотаксиса сосудосуживающее действие")
Эйкозаноиды – производные арахидоновой кислоты (липоксины): медиаторы воспаления агрегация тромбоцитов факторы хемотаксиса сосудосуживающее действие
 – активаторы пролиферации (усиление репликации в зависимости от вида фактора")
Цитокины: Факторы роста (GF) – активаторы пролиферации (усиление репликации в зависимости от вида фактора → как правило при репарации); ВЭФР (VEGF) – усиление пролиферации эндотелия при атеросклерозе, ИБС. Интерлейкины (ИЛ) – участвуют в координации деятельности лейкоцитов эпителия и развития различных патологических состояний.

Хемокины – управляют перемещением в пределах тканей различных клеток и образование контактов с тканью и между собой. Хемоаттрактанты – вещества, привлекающие клетки; Адгезивные молекулы – вещества, прикрепляющие клетки.
 в нормальных клетках и")
Факторы роста и некроза тканей управляют апоптозом (ингибируют и активируют) в нормальных клетках и тканях. Колониестимулирующие факторы – способствуют пролиферации клеток из одной клетки.

Способы регуляции продукции гормонов эндокринными железами. 1. Контроль метаболитами Пример: глюкоза + рецепторы ПЖЖ → изменение биосинтеза глюкагона и инсулина → регуляция глюкозы в крови. внутренние сигналы + эндокринные железы − − клетки-мишени

2. Контроль нервной системой Пример: биосинтез и секреция катехоламинов надпочечниками регулируется импульсами вегетативными центрами гипоталамуса → выброс катехоламинов в ответ на эмоции, боль и т. д. внешние и внутренние сигналы + + ЦНС + + эндокринные железы клетки-мишени

3. Нейро-эндокринный контроль Под прямым контролем нервной системы находится мозговое вещество надпочечников и гипоталамуса. Другие эндокринные железы контролируются нервной системой опосредованно – через гормоны гипоталамуса и гипофиза.

ЦНС Сигналы + Гипоталамус либерины статины + - Гипофиз тропные гормоны + - Эндокринные железы + Клетки-мишени - -
 либерины и статины, которые, соответственно, стимулируют (+) или")
Гипоталамус – синтезирует рилизинг-гормоны (пептидной природы) либерины и статины, которые, соответственно, стимулируют (+) или ингибируют (-) синтез и секрецию тропных гормонов передней доли гипофиза. Тропные гормоны гипофиза: - регулируют образование и секрецию гормонов периферических эндокринных желёз и частично действуют на обмен веществ периферических тканей и органов.
, трийодтиронин (Т 3), кальцитонин. Паращитовидные –")
Периферические эндокринные железы: Щитовидная - тироксин (Т 4), трийодтиронин (Т 3), кальцитонин. Паращитовидные – паратгормон. ПЖЖ – инсулин, глюкагон. Надпочечники (кора) – кортикостероиды, минералокортикоиды, андрогены. Надпочечники (мозговое вещество) – катехоламины (дофамин, адреналин, норадреналин). Половые железы – семенники (андрогены), яичники (эстрогены, прогестины).

Гормоны периферических эндокринных желёз поступают в общий кровоток и взаимодействуют с клетками-мишенями. Поддержание уровня гормонов в организме обеспечивает механизм отрицательной связи. Изменение концентрации метаболитов в клеткахмишенях по механизму отрицательной обратной связи подавляет синтез гормонов, действуя либо на эндокринные железы, либо на гипоталамус. Синтез и секреция тропных гормонов подавляется гормонами эндокринных желез.

Механизмы передачи гормональных сигналов в клетки Гормоны пептидной природы, адреналин, гормоны местного действия (цитокины, эйкозаноиды) – взаимодействуют через рецепторы мембран и G-белок (аденилатциклазным, гуанилатциклазным, инозитолфосфатным) и тирозинкиназным механизмами. Стероидные гормоны, тироксин – взаимодействуют через рецепторы, расположенные внутри клетки (в цитозоле или в ядре) – ядерный механизм.
 действуют на образование циклических")
Аденилатциклазный механизм Внеклеточные регуляторы-гормоны (первичные посредники: глюкагон, адреналин через β-рецептор) действуют на образование циклических нуклеотидов через сигнальную систему – G-белок, аденилатциклазу (ГЦ), которая контролирует образование ц. АМФ (ц. ГМФ) (вторичных посредников, мессенджеров). Аденилатциклаза (ГЦ) встроена в мембрану. Гормон + рецептор → G-белок →АЦ (ГЦ)→ АТФ (ГТФ)→ ц. АМФ (ц. ГМФ)→ активирует протеинкиназу А→ фосфорилирование белка → ответ клетки-мишени.

Аденилатциклазный механизм
 Адреналин (через α-рецепторы), СТГ – активируют G-белок → фосфолипазу")
Инозитолфосфатный механизм (см. Обмен липидов) Адреналин (через α-рецепторы), СТГ – активируют G-белок → фосфолипазу С → ФЛ → вторичные посредники (ИТФ и ДАГ) → активируют протеинкиназу С и увеличение внутриклеточной концентрации Са 2+ → фосфорилирование белка → ответ клетки-мишени.

Инозитолфосфатный механизм

Рецепторы с тирозинкиназной активностью Тирозиновые протеинкиназы – ферменты, фосфорилирующие специфические белки по тирозину. Рецептор инсулина – тирозиновая протеинкиназа, фосфорилирующая белки по ОН-группам тирозина. Рецептор состоит из 2 α- и β-субъединиц, связанных дисульфидными и нековалентными взаимодействиями.

α-субъединицы на поверхности мембраны: центр связывания инсулина образован Nконцевыми доменами α-субъединиц. β-субъединицы пронизывают мембрану, не связывают инсулин. Инсулин связывается с α- субъединицами, идет фосфорилирование β-субъединиц по нескольким тирозиновым остаткам → и эта тирозиновая протеинкиназа способна фосфорилировать другие внутриклеточные белки. В отсутствие гормона инсулиновые рецепторы не проявляют тирозинкиназной активности.
 Взаимодействие с рецептором в цитоплазме или")
Механизм с участием внутриклеточных рецепторов и ДНК (ядерный) Взаимодействие с рецептором в цитоплазме или ядре (стероидные гормоны) Взаимодействие гормонрецепторного комплекса со специфическими участками ДНК (гормончувствительными элементами) Индукция транскрипции Изменение метаболизма клетки путем усиления биосинтеза белков

Ядерный механизм гормон
 – активируется частичным протеолизом,")
Гормоны гипоталамуса - - Пептидной природы. Либерины: Тиреолиберин (3 АМК) – активируется частичным протеолизом, ↑АД, ↑двигательную активность. Кортиколиберин (41 АМК) – стимулирует секрецию кортикотропина. Гонадолиберин (10 АМК) – стимулирует гормоны гипофиза (ЛГ, ФСГ). Соматолиберин (40 -44 АМК) – стимулирует секрецию СТГ.
 – в ЦНС, (28 АМК) – в кишечнике -")
Статины: - Соматостатин (14 АМК) – в ЦНС, (28 АМК) – в кишечнике - ↓ уровень внутриклеточных ц. АМФ и Са 2+ в цитозоле клеток; ↓ секрецию гормона роста, глюкагона, инсулина, гастрина, секретина, ренина, паратгормона.
 синтезируются тропные гормоны (3 группы) – стимулируют синтез")
Гормоны гипофиза В передней доле (аденогипофизе) синтезируются тропные гормоны (3 группы) – стимулируют синтез и секрецию гормонов других эндокринных желёз. 1 группа: - Гормон роста, СТГ (191 АМК) – в печени ↑ глюконеогенез, синтез белка; ↑ рост костей, ↑ липолиз в жировой ткани, в мышцах ↓ утилизация глюкозы и ↑ синтез белка. - Пролактин (197 АМК) – стимуляция лактации.
 – стимулирует синтез и секрецию йодтиронинов в")
2 группа: - ТТГ (96 АМК, гликопротеин) – стимулирует синтез и секрецию йодтиронинов в щитовидной железе. - Лютеинизирующий гормон (96 АМК, гликопротеин) – стимулирует овуляцию у женщин, у мужчин синтез андрогенов в клетках Лейдига. - ФСГ (96 АМК, гликопротеин) – стимулирует рост фолликулов у женщин и сперматогенез у мужчин.
 – стимулирует рост надпочечников и синтез кортикостероидов.")
3 группа: - Кортикотропин (АКТГ, 39 АМК) – стимулирует рост надпочечников и синтез кортикостероидов. В задней доле (нейрогипофизе) – секретируются гормоны, регулирующие водный баланс (вазопрессин (АДГ), 9 АМК) и лактацию (окситоцин, 9 АМК).
: 1) Глюкокортикоиды (С 21 – стероиды, кортизол) -")
Стероидные гормоны Гормоны коры надпочечников (кортикостероиды): 1) Глюкокортикоиды (С 21 – стероиды, кортизол) - роль адаптации к стрессу, стимуляция глюконеогенеза. 2) Минералокортикоиды (С 21 – стероиды, альдостерон) – роль поддержания уровня Na+ и K+. 3) Андрогены (С 19 – стероиды).
. Отщепление боковой цепи ХС :")
Синтез стероидных гормонов Предшественник – холестерин (поступает с ЛПНП). Отщепление боковой цепи ХС : 2 реакции гидроксилирования (гидроксилаза Р 450) по атому С 22 и С 20 → прегненолон → прогестерон. В коре надпочечников 3 зоны: - в пучковой, сетчатой – прогестерон гидроксилируется с образованием глюкокортикоидов (кортизол), андрогенов; - в клубочковой – минералокортикоиды (альдостерон).
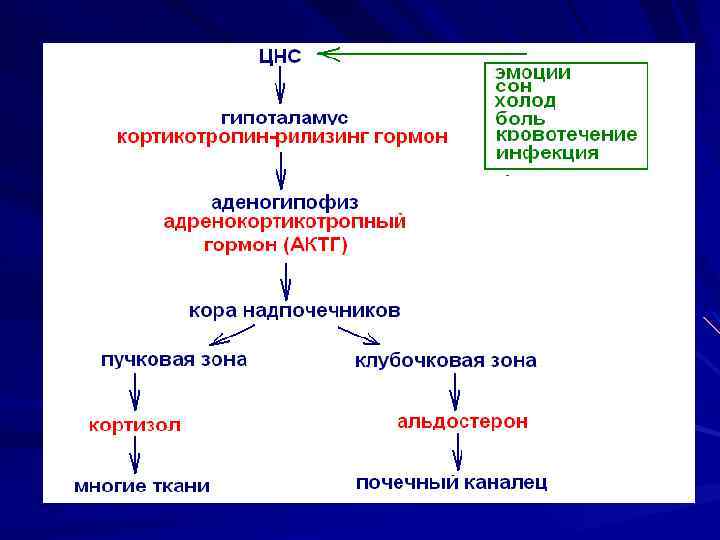

Стимул для синтеза кортизола – стресс, травма; для синтеза и секреции альдостерона – ангиотензин II, АКТГ, простагландин Е, ↑ концентрация К+ и ↓ Na+.

Влияние глюкокортикоидов Кортизол: - стимулирует образование глюкозы в печени, усиливая глюконеогенез; - увеличивает скорость освобождения АМК из периферических тканей для глюконеогенеза; - в печени индуцирует синтез ферментов катаболизма АМК; - тормозит потребление глюкозы периферическими тканями (при голодании и недостатке инсулина).

В печени кортизол стимулирует синтез белков и нуклеиновых кислот. В мышцах, лимфоидной и жировой ткани, коже и костях кортизол тормозит синтез белков, РНК и ДНК и стимулирует распад белка и РНК. Избыточное количество кортизола стимулирует липолиз в конечностях и липогенез в тканях туловища и лица, усиливает липолитическое действие катехоламинов и гормонов роста.

Высокая концентрация глюкокортикоидов: - подавляют иммунные реакции, вызывая гибель лимфоцитов; - подавляют воспалительную реакцию снижая число лейкоцитов и индуцируя синтез липокортинов, которые ингибируют фосфолипазу А 2 (↓ медиаторы воспаления); - тормозят рост и деление фибробластов, синтез коллагена и фибронектина.

Действие альдостерона:

Главный механизм регуляции синтеза и секреции альдостерона – ренин- ангиотензиновая система. Ренин – протеолитический фермент, продуцируемый юкстагломерулярными клетками почечных афферентных артериол. Кровотечение, ↓АД приводит к высвобождению ренина → субстрат для него ангиотензиноген (α 2 -глобулин, синтезируется в печени).

Ренин гидролизует пептидную связь в ангиотензиногене и отщепляет Nконцевой декапептид → ангиотензин I. Ангиотензинпревращающий фермент (АПФ) с С-конца ангиотензина I отщепляет 2 АМК → ангиотензин II (октапептид). Ангиотензин II стимулирует выработку альдостерона через ИТФ, вызывает сужение артериол → ↑АД, вызывает жажду → реабсорбция Na+ и воды.

Система ренин-ангиотензин-альдостерон:

Гормоны щитовидной железы Тиреолиберин гипоталамуса стимулирует секрецию тиреотропина (пик секреции перед сном, в течении ночи ↓). ТТГ – гликопротеид аденогипофиза, стимулирует синтез и секрецию йодтиронинов (Т 3 , Т 4) в щитовидной железе (через аденилатциклазную систему).
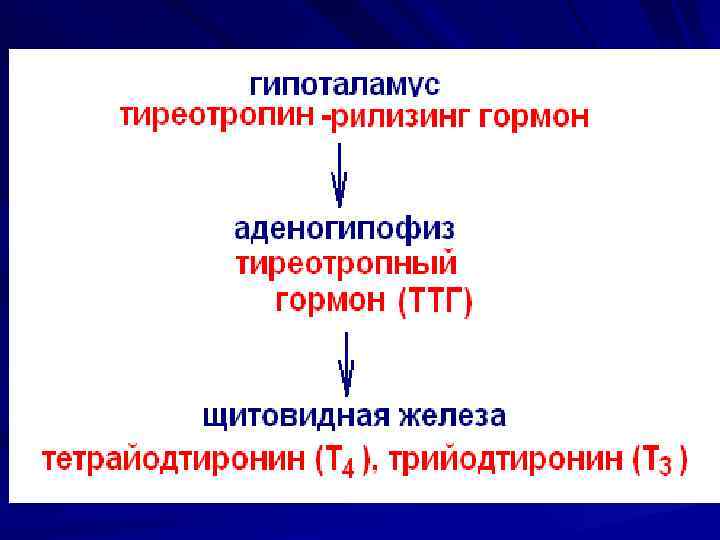

Гормоны щитовидной железы:
 в фолликулах щитовидной")
Йодтиронины – йодированные производные тирозина, синтезируются в составе белка тиреоглобулина (Тг) в фолликулах щитовидной железы. Тг – гликопротеин, содержит 115 остатков тирозина, синтез идет на рибосомах шероховатой ЭПС, вторичная и третичная структура формируется в цистернах ЭПС → в аппарат Гольджи → секретируется во внеклеточный коллоид (хранение и йодирование).
 Транспорт йода в клетки щитовидной железы – 25 -30% йодидов,")
Этапы образования йодтиронинов 1) Транспорт йода в клетки щитовидной железы – 25 -30% йодидов, от поступивших соединений йода через ЖКТ с пищей, захватывается щитовидной железой при участии йодидпереносящего белка против градиента. 2) Окисление йода - I- → I+ с участием гемсодержащей тиреопероксидазы и Н 2 О 2. 3) Йодирование тирозина - I+ + тирозин (Тг). 4) Образование йодтиронинов –монойодтирозин (МИТ) и дийодтирозин (ДИТ). Конденсируются: ДИТ+ДИТ = Т 4 ДИТ + МИТ = Т 3 → фолликул.

Основные эффекты йодтиронинов – результат взаимодйствия с рецепторами ядра, которые взаимодействуют с ДНК и регулируют экспрессию генов → ускорение белкового синтеза, стимуляция процессов роста и клеточной дифференцировки. Повышают поглощение клетками кислорода (во всех органах, кроме мозга, РЭС и гонад). Стимулируют работу Na+/К+-АТФ-азы в разных клетках. В печени ускоряют гликолиз, синтез холестерола и желчных кислот.

В печени и жировой ткани йодтиронины повышают чувствительность клеток к адреналину и косвенно стимулируют липолиз в жировой ткани и мобилизацию гликогена в печени. В мышцах увеличивают потребление глюкозы, стимулируют синтез белков и увеличение мышечной массы. Увеличивают теплопродукцию как реакцию на охлаждение, повышают чувствительность симпатической нервной системы к норадреналину.
. Недостаточное поступление йода")
Недостаточность функции щитовидной железы – гипотиреоз (микседема – тяжелая форма гипотиреоза). Недостаточное поступление йода с пищей – эндемический зоб (ниже 100 мкг/сут) Гипертиреоз – диффузный токсический зоб (базедова болезнь).

Гормоны, поддерживающие гомеостаз кальция Кальцитриол Паратиреоидный гормон Кальцитонин

Кальцитонин Секретируется парафолликулярными Кклетками щитовидной железы или Склетками паращитовидной железы: полипептид, 32 АМК. Секреция ↑ при увеличении концентрации Са 2+ и ↓ при понижении Са 2+ в крови. Кальцитонин ингибирует высвобождение Са 2+ из кости и стимулирует экскрецию почками с мочой.

КАЛЬЦИТРИОЛ – АКТИВНАЯ ФОРМА ВИТАМИНА Д ОБРАЗОВАНИЕ 7 -дегидрохолестерин УФ в коже Витамин Д (холекальциферол) печень 25 -гидроксихолекальциферол почки 1, 25 -дигидроксихолекальциферол (кальцитриол) СИНТЕЗ зависит от обеспеченности витамином Д, стимулируется в почках ПГ ГИПЕРКАЛЬЦИЕМИЧЕСКИЙ ЭФФЕКТ КИШЕЧНИК стимулирует всасывание кальция и фосфатов КОСТИ в норме способствует минерализации, поддерживая необходимую концентрацию кальция и фосфатов во внеклеточной жидкости, при гипокальциемии стимулирует резорбцию остеокластами
 СИНТЕЗИРУЕТСЯ ОКОЛОЩИТОВИДНЫМИ ЖЕЛЕЗАМИ СИНТЕЗ стимулируется гипокальциемией подавляется кальцитриолом ГИПЕРКАЛЬЦИЕМИЧЕСКИЙ")
ПАРАТИРЕОИДНЫЙ ГОРМОН ПОЛИПЕПТИД (84 АМК) СИНТЕЗИРУЕТСЯ ОКОЛОЩИТОВИДНЫМИ ЖЕЛЕЗАМИ СИНТЕЗ стимулируется гипокальциемией подавляется кальцитриолом ГИПЕРКАЛЬЦИЕМИЧЕСКИЙ ЭФФЕКТ ПОЧКИ стимулирует реабсорбцию кальция, подавляет реабсорбцию фосфатов стимулирует образование кальцитриола КОСТИ при гипокальциемии стимулирует резорбцию остеокластами

Половые гормоны Репродуктивные функции организма регулируются половыми гормонами: у мужчин – тестостероном, у женщин – эстрогенами и прогестероном. Находятся под контролем ФСГ и ЛГ связываются с рецепторами на мембранах своих клеток-мишеней в яичниках и яичках → активация аденилатциклазной системы.

Синтез половых гормонов:

Мужской гаметогенез:

Женский гаметогенез:

Гормоны поджелудочной железы Инсулин – полипептид (2 цепи: 21 и 30 АМК, соединены дисульфидными мостиками). Синтез - β-клетки островков Лангерганса. Разрушение инсулина – в печени (инсулиназа) Глюкоза регулирует экспрессию гена инсулина напрямую взаимодействуя с транскрипционными факторами или через влияние на секрецию инсулина и глюкагона. Синтез стимулируется глюкозой, а секреция – Са 2+- зависимый процесс (при дефиците Са 2+ снижается даже при высоком содержании глюкозы).

Адреналин через α 2 -рецепторы тормозит секрецию инсулина, β-адренергические агонисты её стимулируют за счет повышения ц. АМФ. Высокие концентрации гормона роста, кортизола, эстрогенов – стимулируют секрецию инсулина. Гликопротеиновые рецепторы инсулина – почти во всех клетках, но больше их в гепатоцитах и адипоцитах. Рецептор инсулина относят к типу рецепторов с тирозинкиназной активностью.

Эффекты инсулина Анаболический гормон – влияет на: -ключевые ферменты метаболизма белков, жиров и углеводов (активация, индукция, репрессия) (стр. 582 Северин); - транспорт глюкозы в мышцы и жировую ткань (Glut – 4); - процессы репликации и транскрипции, участвуя таким образом в регуляции дифференцировки клеток, пролиферации; - метаболизм глюкозы (↑ глюкокиназы, фосфофруктокиназы, пируваткиназы)
 Синтез - α-клетки островков Лангерганса. На секрецию влияют: глюкоза,")
Глюкагон – полипептид (29 АМК) Синтез - α-клетки островков Лангерганса. На секрецию влияют: глюкоза, аминокислоты, жирные кислоты, кетоновые тела, нейромедиаторы. Углеводная пища – снижает синтез, белковая – стимулирует (аланин). Распад глюкагона – в печени протеазами. Эффекты глюкагона – противоположны инсулину, стимулирует глюконеогенез, через АЦ стимулирует липолиз (↑ ТАГ-липазу). Клетки мишени – печень и жировая ткань.
Регуляция обмена веществ (2).ppt