1 Regulation1.ppt
- Количество слайдов: 75



РЕГУЛЯЦИЯ КЛЕТОЧНОЙ АКТИВНОСТИ КЛАССИФИКАЦИЯ СПОСОБОВ РЕГУЛЯЦИИ

СПОСОБЫ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ I. ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ II. ХИМИЧЕСКАЯ МОДИФИКАЦИЯ БЕЛКОВ III. ИЗМЕНЕНИЕ КОМПАРТМЕНТАЛИЗАЦИИ ВЕЩЕСТВ В КЛЕТКЕ IV. ТОПОДИНАМИЧЕСКАЯ РЕГУЛЯЦИЯ V. ЭКСПРЕССИЯ ГЕНОМА VI. РЕГУЛЯТОРНЫЕ СИСТЕМЫ С УЧАСТИЕМ ВТОРИЧНЫХ ПОСРЕДНИКОВ
 ЦЕНТРА")
I. ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ 1. РЕГУЛЯЦИЯ НА УРОВНЕ КАТАЛИТИЧЕСКОГО (АКТИВНОГО) ЦЕНТРА АКТИВНОСТЬ ФЕРМЕНТА ОПРЕДЕЛЯЕТСЯ ЕГО КАТАЛИТИЧЕСКИМ ДЕЙСТВИЕМ

ВЗАИМОДЕЙСТВИЕ МЕТАБОЛИТОВ И КОФАКТОРОВ С ФЕРМЕНТАМИ 2. РЕГУЛЯЦИЯ НА УРОВНЕ АЛЛОСТЕРИЧЕСКОГО ЦЕНТРА АКТИВНОСТЬ ФЕРМЕНТА РЕГУЛИРУЕТСЯ ЛИГАНДАМИ: СУБСТРАТОМ, КОНЕЧНЫМ ПРОДУКТОМ РЕАКЦИИ, КОФЕРМЕНТОМ , КОТОРЫЕ СВЯЗЫВАЮТСЯ С АЛЛОСТЕРИЧЕСКИМ ЦЕНТРОМ
 МОДИФИКАЦИЯ")
II. ХИМИЧЕСКАЯ МОДИФИКАЦИЯ БЕЛКОВ (ПОСТТРАНСЛЯЦИОННАЯ БЕЛКОВ) МОДИФИКАЦИЯ

СИНТЕЗ БЕЛКА НА РИБОСОМЕ В клетках животных многие белки синтезируются в виде молекул—предшественников, требующих модификации для приобретения биологической активности.

ковалентные модификации белка • ГИДРОКСИЛИРОВАНИЕ • ГЛИКОЗИЛИРОВАНИЕ • АЦЕТИЛИРОВАНИЕ • МЕТИЛИРОВАНИЕ • ФОСФОРИЛИРОВАНИЕ • ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ

ГИДРОКСИЛИРОВАНИЕ - присоединение ОН- к определенным остаткам аминокислот ПРИМЕР: коллаген синтезируется в виде проколлагена. Гидроксилирование остатков пролина и лизина проколлагеновых цепей приводит к образованию стабилизирующих перекрестных сшивок. Затем – отщепление концевых пептидов и образование конечного продукта – прочной нерастворимой молекулы коллагена.

ГЛИКОЗИЛИРОВАНИЕ - присоединение углеводных остатков, образование гликопротеинов Перенос олигосахаридов Происходит в ЭПС и комплексе Гольджи Укорачивание олигосахаридов

АЦЕТИЛИРОВАНИЕ и МЕТИЛИРОВАНИЕ Присоединение ацильной или метильной группы ПРИМЕР: ацетилирование или метилирование гистонов, что влияет на транскрипцию Аминокислотная последовательность гистона Н 4

ПОСТТРАНСЛЯЦИОННАЯ МОДИФИКАЦИЯ ГИСТОНОВ ОКАЗЫВАЕТ ВЛИЯНИЕ НА ТРАНСКРИПЦИОННУЮ АКТИВНОСТЬ ГЕНОВ АЦЕТИЛИРОВАНИЕ НЕЙТРАЛИЗУЕТ ПОЛОЖИТЕЛЬНЫЙ ЗАРЯД ЛИЗИНА, ФОСФОРИЛИРОВАНИЕ ПРИДАЕТ ОТРИЦАТЕЛЬНЫЙ ЗАРЯД СЕРИНУ ВОЗМОЖНЫЙ МЕХАНИЗМ: ИЗМЕНЕНИЕ ВЕЛИЧИНЫ ПОЛОЖИТЕЛЬНОГО ЗАРЯДА ГИСТОНОВ, ЧТО ВЛИЯЕТ НА ПРОЧНОСТЬ СВЯЗИ ГИСТОНОВ С ОТРИЦАТЕЛЬНО ЗАРЯЖЕННЫМ ОСТОВОМ ДНК.

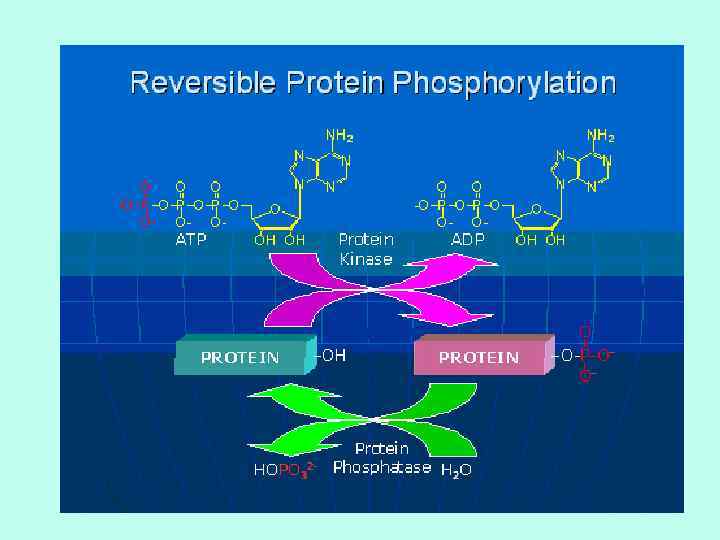
ФОСФОРИЛИРОВАНИЕ • Присоединение фосфатной группы. • Обратимый процесс. • Фосфорилируется остаток серина или тирозина. • Несмотря на большое количество остатков серина или тирозина, фосфорилированию избирательно подвергается их малое (1 – 3) число. • В зависимости от конкретного случая более активным может быть либо фосфо- либо дефосфофермент. • Фосфорилированию подвергаются, кроме ферментов, белки транспортных систем, цитоскелета и др.

ФОСФОРИЛИРОВАНИЕ КИНАЗА ФОСФОРИЛИРУЕТ ФОСФАТАЗА ДЕФОСФОРИЛИРУЕТ

ОГРАНИЧЕННЫЙ ПРОТЕОЛИЗ – избирательное расщепление белка на фрагменты при участии протеаз Многие ферменты производятся в виде протоферментов. Для их активизации происходит отщепление фрагмента полипептидной цепи. Благодаря ограниченному протеолизу клетка в ответ на сигнал может увеличить количество активного фермента без транскрипции и трансляции.

ПРИМЕР: АКТИВАЦИЯ ТРИПСИНОГЕНА

Инсулин синтезируется в виде проинсулина и представляет собой одноцепочечную молекулу. После удаления специфическими протеазами полипептидного сегмента он преобразуется в двухцепочечную молекулу с внутри— и межцепочечными дисульфидными мостиками.

ВНУТРИКЛЕТОЧНАЯ Са 2+ -ЗАВИСИМАЯ ПРОТЕАЗА КАЛЬПАИН Са-АТФаза Нарушение цитоскелета Повреждение мембраны ПОДВЕРГАЕТ ПРОТЕОЛИЗУ МНОГИЕ ВНУТРИКЛЕТОЧНЫЕ БЕЛКИ: БЕЛКИ КАНАЛОВ, ЦИТОСКЕЛЕТА И ДР. Белки ионных каналов

ПРОТЕАЗА КАЛЬПАИН

III. ИЗМЕНЕНИЕ КОМПАРТМЕНТАЛИЗАЦИИ ВЕЩЕСТВ В КЛЕТКЕ Ферменты и субстраты разделены мембраной, изменение ее проницаемости для субстратов, влияет на ход ферментативных реакций.

IV. ТОПОДИНАМИЧЕСКАЯ РЕГУЛЯЦИЯ Основана на динамической неоднородности распределения белков в мембране, их способности образовывать ассоциации.

ПРИМЕР: дыхательная цепь в митохондрии I, III, IV – белковые комплексы II – сукцинатдегидрогеназа V – АТФ-синтаза

V. ЭКСПРЕССИЯ ГЕНОМА РЕГУЛЯЦИЯ СОДЕРЖАНИЯ БЕЛКА В КЛЕТКЕ ПОСРЕДСТВОМ ИНДУКЦИИ И РЕПРЕССИИ СИНТЕЗА, Т. Е. ИЗМЕНЕНИЕМ СКОРОСТИ ТРАНСКРИПЦИИ

РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА У ПРОКАРИОТ ИНДУКТОРОМ СИНТЕЗА СЛУЖИТ СУБСТРАТ

Во всех эукариотических клетках экспрессия генов контролируется регуляторными белками, которые связываются с определенными участками ДНК и стимулируют или подавляют транскрипцию. Регуляторные элементы, стимулирующие транскрипцию, называют ЭНХАНСЕРАМИ (от англ. enchance – усиливать ). Белки, подавляющие транскрипцию, называют САЙЛЕНСЕРАМИ (от англ. silence – заглушать)

КОНТРОЛЬ НА УРОВНЕ ТРАНСКРИПЦИИ РЕГУЛЯЦИЯ СИНТЕЗА БЕЛКА У ЭУКАРИОТ ИНДУКТОРЫ СИНТЕЗА – СТЕРОИДНЫЕ ГОРМОНЫ

VI. РЕГУЛЯТОРНЫЕ СИСТЕМЫ С УЧАСТИЕМ ВТОРИЧНЫХ ПОСРЕДНИКОВ КОМПОНЕНТЫ СИСТЕМЫ РЕЦЕПТОР ПРЕОБРАЗОВАТЕЛЬ СИГНАЛА – G БЕЛОК ФЕРМЕНТ – УСИЛИТЕЛЬ СИГНАЛА, ОБРАЗУЮЩИЙ ВТОРИЧНЫЕ ПОСРЕДНИКИ ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ Связывание с лигандом Активация рецептора Преобразование сигнала ПРОТЕИНКИНАЗА Активация эффектора БЕЛОК-ЭФФЕКТОР Физиологический эфект

СПОСОБЫ МЕЖКЛЕТОЧНОЙ КОММУНИКАЦИИ СПОСОБЫ ДОСТАВКИ СИГНАЛЬНЫХ МОЛЕКУЛ К КЛЕТКАМ
 1 НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ 2 ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ 3 ГИДРОФИЛЬНЫЕ")
МЕЖКЛЕТОЧНЫЕ СИГНАЛЬНЫЕ МОЛЕКУЛЫ (ПЕРВИЧНЫЕ ПОСРЕДНИКИ) 1 НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ 2 ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ 3 ГИДРОФИЛЬНЫЕ МОЛЕКУЛЫ ДИФФУНДИРУЮТ ЧЕРЕЗ МЕМБРАНУ И СВЯЗЫВАЮТСЯ С ВНУТРИКЛЕТОЧНЫМИ РЕЦЕПТОРАМИ ВЗАИМОДЕЙСТВУЮТ С МЕМБРАННЫМИ РЕЦЕПТОРАМИ

Первичные посредники 1. НЕБОЛЬШИЕ ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ • Стероидные гормоны, вырабатываемые половыми железами и корой надпочечников • тиреоидные гормоны

СХЕМА ДЕЙСТВИЯ СТЕРОИДНЫХ И ТИРЕОИДНЫХ ГОРМОНОВ внутриклеточные рецепторы имеют • гормон-связывающий домен • ДНК-связывающий домен


МЕХАНИЗМ ДЕЙСТВИЯ ЛИПОФИЛЬНЫХ ГОРМОНОВ, ПРОНИКАЮЩИХ ВНУТРЬ КЛЕТКИ

Первичные посредники 2. ЛИПОФИЛЬНЫЕ МОЛЕКУЛЫ Производные арахидоновой кислоты – ЭЙКОЗАНОИДЫ: • ПРОСТАГЛАНДИНЫ • ЛЕЙКОТРИЕНЫ • ПРОСТАЦИКЛИНЫ • ТРОМБОКСАНЫ

Первичные посредники ЭЙКОЗАНОИДЫ ОБРАЗУЮТСЯ ПРАКТИЧЕСКИ ВО ВСЕХ КЛЕТКАХ, ИХ БИОСИНТЕЗ ИНИЦИИРУЕТСЯ ФОСФОЛИПАЗОЙ А 2 ДЕЙСТВУЮТ ЧЕРЕЗ МЕМБРАННЫЕ РЕЦЕПТОРЫ, ОСУЩЕСТВЛЯЮТ ПАРАКРИННОЕ И АУТОКРИННОЕ ДЕЙСТВИЕ ЭЙКОЗАНОИДЫ РЕГУЛИРУЮТ • СОКРАЩЕНИЕ ГЛАДКОМЫШЕЧНЫХ КЛЕТОК • БОЛЕВЫЕ И ВОСПАЛИТЕЛЬНЫЕ РЕАКЦИИ • СЕКРЕЦИЮ ЖЕЛУДОЧНОГО СОКА • АГРЕГАЦИЮ ТРОМБОЦИТОВ И ДР.

Первичные посредники 3. ГИДРОФИЛЬНЫЕ МОЛЕКУЛЫ • ГИДРОФИЛЬНЫЕ ГОРМОНЫ НЕЙРОТРАНСМИТТЕРЫ • ФАКТОРЫ РОСТА (эпидермальный фактор роста, фактор роста нейронов, фактор роста фибробластов и др. ) • ЦИТОКИНЫ (интерлейкины, интерфероны и др. )

Первичные посредники СХЕМА ДЕЙСТВИЯ НЕЙРОМЕДИАТОРОВ

Первичные посредники СХЕМА ДЕЙСТВИЯ ГИДРОФИЛЬНЫХ АГОНИСТОВ Гидрофильные гормоны Инсулин, факторы роста

РЕЦЕПТОРЫ И МЕХАНИЗМЫ ИХ ДЕЙСТВИЯ

РЕЦЕПТОР –СПЕЦИФИЧЕСКИЙ БЕЛОК, КОТОРЫЙ СВЯЗЫВАЕТ СИГНАЛЬНОЕ ВЕЩЕСТВО, ЧТО ПРИВОДИТ В КОНЕЧНОМ ИТОГЕ К РАЗВИТИЮ КЛЕТОЧНОЙ РЕАКЦИИ. АГОНИСТ ПЕРЕВОДИТ РЕЦЕПТОР В АКТИВНОЕ СОСТОЯНИЕ, АНТАГОНИСТ – В НЕАКТИВНОЕ.

ХАРАКТЕРИСТИКИ РЕЦЕПТОРОВ

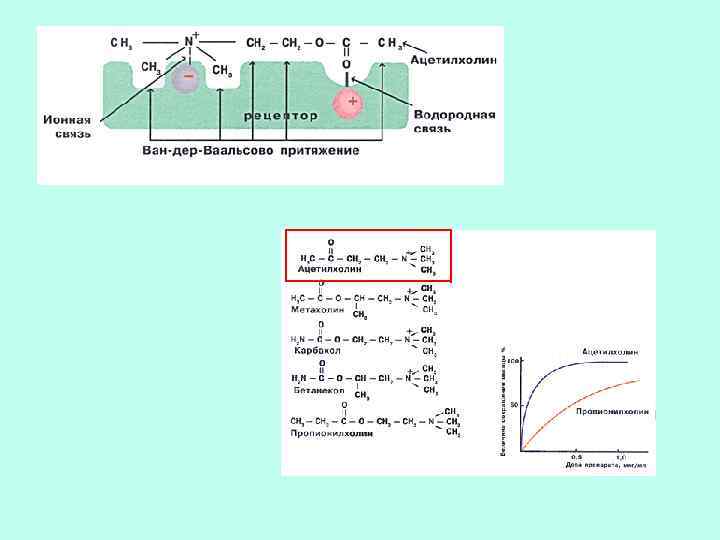
СЕЛЕКТИВНОСТЬ СВОЙСТВО, ОСНОВАННОЕ НА СТРОГОЙ СТРУКТУРНОЙ СПЕЦИФИЧНОСТИ. ДАННЫЙ ЛИГАНД ДОЛЖЕН БЫТЬ ЛИБО ЕДИНСТВЕННЫМ ВЕЩЕСТВОМ, СВЯЗЫВАЮЩИМСЯ С РЕЦЕПТОРОМ, ЛИБО ЭФФЕКТИВНО КОНКУРИРОВАТЬ С ДРУГИМИ.

НАСЫЩАЕМОСТЬ ЧИСЛО МЕСТ СВЯЗЫВАНИЯ С ЛИГАНДОМ ДОЛЖНО БЫТЬ КОНЕЧНЫМ.

СРОДСТВО К ЛИГАНДУ НАСЫЩЕНИЕ РЕЦЕПТОРА ПРОИСХОДИТ ПРИ ФИЗИОЛОГИЧЕСКИХ КОНЦЕНТРАЦИЯХ ЛИГАНДА АКТГ — 0 -50 пг/мл Тироксин общий (Т 4) — 62 -141 нмоль/л Кальцитонин — 5, 5 -28 пмоль/л. Данные приведены для плазмы крови

ТКАНЕВАЯ СПЕЦИФИЧНОСТЬ СВЯЗЫВАНИЕ ЛИГАНДА С РЕЦЕПТОРОМ ПРОИСХОДИТ В ТОЙ ТКАНИ, ГДЕ НАБЛЮДАЕТСЯ ЕГО БИОЛОГИЧЕСКОЕ ДЕЙСТВИЕ.

ОБРАТИМОСТЬ ДЕЙСТВИЯ БЛОКАДА СВЯЗЫВАНИЯ АГОНИСТА С РЕЦЕПТОРОМ ДОЛЖНА ПРИВОДИТЬ К ОБРАТИМОСТИ БИОЛОГИЧЕСКОГО ЭФФЕКТА

КЛАССИФИКАЦИЯ РЕЦЕПТОРОВ

РЕЦЕПТОРЫ РАЗЛИЧАЮТ ПО ИХ ЛОКАЛИЗАЦИИ МЕМБРАННЫЕ ЦИТОПЛАЗМАТИЧЕСКИЕ

ЦИТОПЛАЗМАТИЧЕСКИЕ РЕЦЕПТОРЫ ПРЕДСТАВЛЕНЫ ГЛОБУЛЯРНЫМИ БЕЛКАМИ

МЕМБРАННЫЕ РЕЦЕПТОРЫ ЯВЛЯЮТСЯ ИНТЕГРАЛЬНЫМИ БЕЛКАМИ ОБЩИЙ ПЛАН СТРОЕНИЯ: • ВНЕКЛЕТОЧНЫЙ ДОМЕН СПЕЦИФИЧЕН К ВНЕШНЕМУ СИГНАЛУ • ТРАНСМЕМБРАННЫЙ УЧАСТОК • УЧАСТОК, ПОГРУЖЕННЫЙ В ЦИТОПЛАЗМУ, СПЕЦИФИЧЕН К АССОЦИИРОВАННОМУ С РЕЦЕПТОРОМ ВНУТРИКЛЕТОЧНОМУ БЕЛКУ

Другой подход основан на видах агонистов, с которым взаимодействует рецептор (применяют, как правило, для мембранных рецепторов). НАПРИМЕР • Адренэргические • Холинэргические • Серотонинэргические

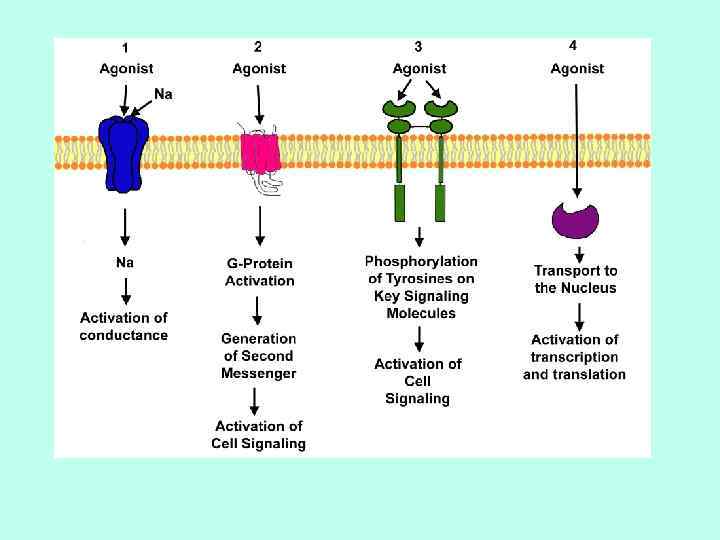
Третий ПОДХОД ОСНОВАН НА МЕХАНИЗМЕ ПЕРЕДАЧИ ВНЕКЛЕТОЧНОГО СИГНАЛА РЕЦЕПТОРЫКАНАЛЫ РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С G-БЕЛКАМИ РЕЦЕПТОРЫ, НЕ ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ, НО СОПРЯЖЕННЫЕ С ТИРОЗИНКИНАЗОЙ


Рецептор-зависимые каналы Неактивное состояние Открыток состояние Рецепторы, связанные с Gбелками R G E R G активированный. G-белок Фермент-связанные рецепторы E инактивированный энзим E

РЕЦЕПТОРЫ-КАНАЛЫ

МЕХАНИЗМ РАБОТЫ РЕЦЕПТОРА-КАНАЛА СВЯЗЫВАНИЕ С ЛИГАНДОМ ПРИВОДИТ К КОНФОРМАЦИОННЫМ ПЕРЕСТРОЙКАМ РЕЦЕПТОРА, ЧТО ВЫЗЫВАЕТ ОТКРЫВАНИЕ ИОННОГО КАНАЛА

Электрический скат, из электрического органа которого были выделены и очищены никотиновые холинэргические рецепторы, расшифрована их аминокислотная последовательность
 ИМЕЕТ 2 УЧАСТКА ДЛЯ СВЯЗЫВАНИЯ")
СОСТОИТ ИЗ 5 СУБЪЕДИНИЦ (2 , , , ) ИМЕЕТ 2 УЧАСТКА ДЛЯ СВЯЗЫВАНИЯ АЦЕТИЛХОЛИНА ПРОНИЦАЕМ ДЛЯ ИОНОВ НАТРИЯ, КАЛЬЦИЯ БЛОКАТОР БУНГАРОТОКСИН

ДРУГИЕ ПРИМЕРЫ: 1. РЕЦЕПТОРЫ К ГЛУТАМАТУ 2. РЕЦЕПТОРЫ К АСПАРТАТУ 3. ГЛИЦИНОВЫЕ РЕЦЕПТОРЫ 4. ГАМК-рецепторы 1, 2 проницаемы для катионов 3, 4 проницаемы для ионов хлора

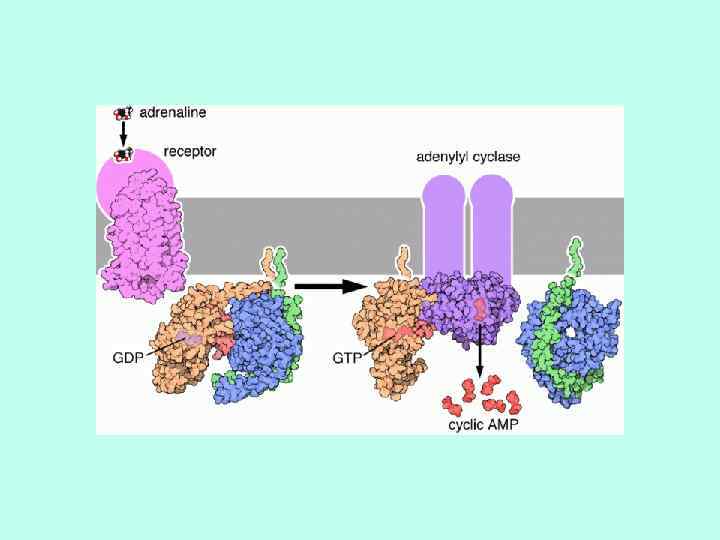
РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С БЕЛКАМИ G-

СТРУКТУРА РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С БЕЛКОМ Участки гликозилирования N-концевой домен С-концевой домен G-

ПРИМЕРЫ АГОНИСТОВ, ДЕЙСТВУЮЩИХ НА РЕЦЕПТОРЫ, СОПРЯЖЕННЫЕ С GБЕЛКАМИ АДРЕНАЛИН, НОРАДРЕНАЛИН АЦЕТИЛХОЛИН СЕРОТОНИН ГИСТАМИН ОПИОИДЫ, КАННАБИНОИДЫ ГОРМОНЫ ПЕПТИДНОЙ ПРИРОДЫ (АКТГ, ВАЗОПРЕССИН, АНГИОТЕНЗИН)

ПРИМЕРЫ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С G-БЕЛКОМ АДРЕНОРЕЦЕПТОР 2 АДРЕНОРЕЦЕПТОР адреналин 2 адренорецептор

Калиевый канал

Кроме ферментов, G -белки могут взаимодействовать с ионтранспортными системами, например, каналами, ионообменниками.

СТРУКТУРА G -БЕЛКА

МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРОВ, СОПРЯЖЕННЫХ С GБЕЛКОМ

ВИДЫ G-БЕЛКОВ ИДЕНТИФИЦИРОВАНО ОКОЛО 20 ВИДОВ G-БЕЛКОВ GS И GI БЕЛКИ ДЕЙСТВУЮТ НА АДЕНИЛАТЦИКЛАЗУ GO ИНГИБИРУЕТ Са-КАНАЛЫ В НЕЙРОНАХ Gq АКТИВИРУЕТ ФОСФОЛИПАЗУ C Gk ОТКРЫВАЕТ КАЛИЕВЫЕ КАНАЛЫ В СЕРДЕЧНЫХ И ДРУГИХ КЛЕТКАХ G-БЕЛКИ СЕНСОРНЫХ КЛЕТОК (ТРАНСДУЦИН ИЛИ Gt В ПАЛОЧКАХ И ДР. ) G-белки, отмеченные цветом, выделены и очищены

ИСПОЛЬЗОВАНИЕ БАКТЕРИАЛЬНЫХ ТОКСИНОВ ПОЗВОЛИЛО ОБНАРУЖИТЬ ДВА ВИДА G-БЕЛКОВ ПО ОТНОШЕНИЮ К АДЕНИЛАТЦИКЛАЗЕ ингибирующий Коклюшный токсин стимулирующий Холерный токсин
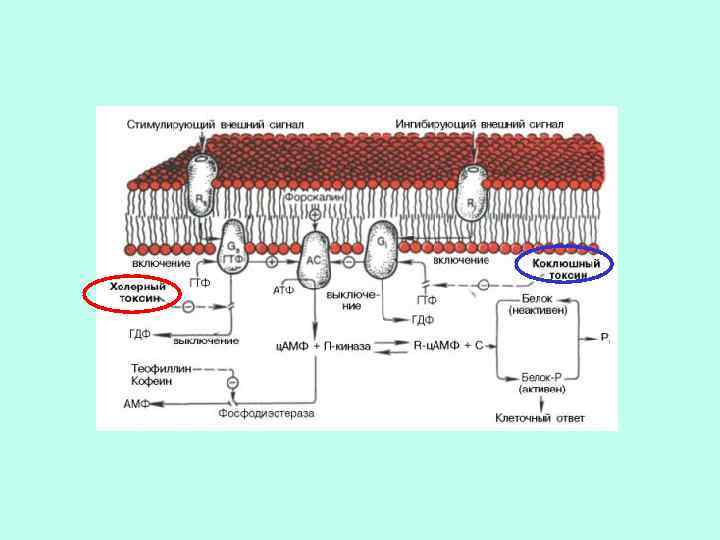

Холерный токсин, также как и коклюшный, содержит фермент АДФ-рибозилазу. АДФрибозилирование -субъединицы GS белка приводит к ее необратимому активированию, т. е. препятствует гидролизу ГТФ. Итог – активация аденилатциклазы и непрекращающаяся наработка ц. АМФ. АДФрибозилирование -субъединицы GI белка препятствует его диссоциации на субъединицы. Результат – устранение ингибирующего действия на аденилатциклазу, и она остается активной.

ВИДЫ G-БЕЛКОВ ПО ОТНОШЕНИЮ К АДЕНИЛАТЦИКЛАЗЕ
1 Regulation1.ppt