3.3_РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ.ppt
- Количество слайдов: 70
 РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ
РЕЦЕПТОРЫ, ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ
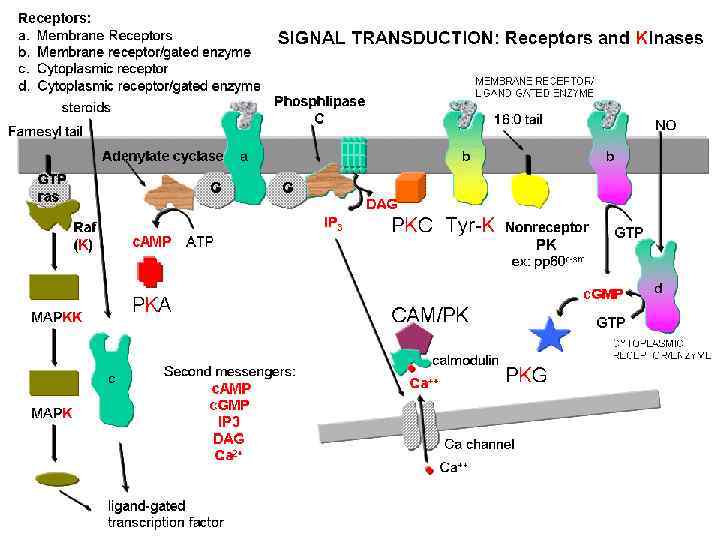
 РЕЦЕПТОРНЫЕ ТИРОЗИНКИНАЗЫ Агонисты: Øфакторы роста Øинсулин
РЕЦЕПТОРНЫЕ ТИРОЗИНКИНАЗЫ Агонисты: Øфакторы роста Øинсулин
 или митогены стимулируют рост клетки и ее прохождение через митоз. GF") Факторы роста (GF) или митогены стимулируют рост клетки и ее прохождение через митоз. GF - полипептиды, состоящие из 50 -100 аминокислот.
Факторы роста (GF) или митогены стимулируют рост клетки и ее прохождение через митоз. GF - полипептиды, состоящие из 50 -100 аминокислот.
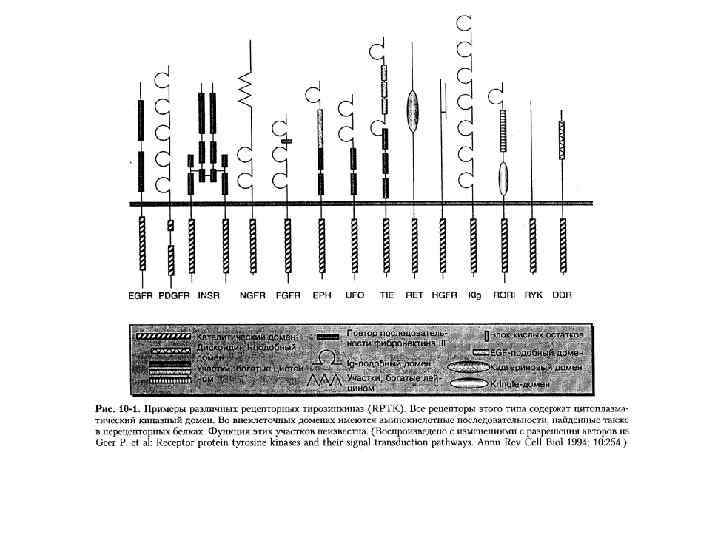
 РЕЦЕПТОРНАЯ ТИРОЗИНКИНАЗА 1 2 3 1 – внеклеточный N-концевой участок, связывающий агонисты; 2 – трансмембранный участок; 3 – внутриклеточный тирозинкиназный домен
РЕЦЕПТОРНАЯ ТИРОЗИНКИНАЗА 1 2 3 1 – внеклеточный N-концевой участок, связывающий агонисты; 2 – трансмембранный участок; 3 – внутриклеточный тирозинкиназный домен
 ØДИМЕРИЗАЦИЯ РЕЦЕПТОРОВ ØТРАНСАВТОФОСФОРИЛИРОВАНИЕ ТИРОЗИНОВЫХ ОСТАТКОВ ØФОРМИРОВАНИЕ УЧАСТКОВ ДЛЯ СВЯЗЫВАНИЯ") МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРНОЙ ТИРОЗИНКИНАЗЫ (РТК) ØДИМЕРИЗАЦИЯ РЕЦЕПТОРОВ ØТРАНСАВТОФОСФОРИЛИРОВАНИЕ ТИРОЗИНОВЫХ ОСТАТКОВ ØФОРМИРОВАНИЕ УЧАСТКОВ ДЛЯ СВЯЗЫВАНИЯ БЕЛКОВ-СУБСТРАТОВ, СОДЕРЖАЩИХ SH 2 и SH 3 ДОМЕНЫ ØФОСФОРИЛИРОВАНИЕ СВЯЗЫВАЮЩИХСЯ С РТК БЕЛКОВ
МЕХАНИЗМ ФУНКЦИОНИРОВАНИЯ РЕЦЕПТОРНОЙ ТИРОЗИНКИНАЗЫ (РТК) ØДИМЕРИЗАЦИЯ РЕЦЕПТОРОВ ØТРАНСАВТОФОСФОРИЛИРОВАНИЕ ТИРОЗИНОВЫХ ОСТАТКОВ ØФОРМИРОВАНИЕ УЧАСТКОВ ДЛЯ СВЯЗЫВАНИЯ БЕЛКОВ-СУБСТРАТОВ, СОДЕРЖАЩИХ SH 2 и SH 3 ДОМЕНЫ ØФОСФОРИЛИРОВАНИЕ СВЯЗЫВАЮЩИХСЯ С РТК БЕЛКОВ
 МЕХАНИЗМ АКТИВАЦИИ Ras. БЕЛКА Связывание лиганда, димеризация рецептора и трансавтофосфорилирование Через специфические белки РТК связывается с неактивным Ras-белком Активизация Ras-белка: замена ГДФ на ГТФ Ключевой компонент сигнального пути, связанного с рецепторной тирозинкиназой – Ras-белок.
МЕХАНИЗМ АКТИВАЦИИ Ras. БЕЛКА Связывание лиганда, димеризация рецептора и трансавтофосфорилирование Через специфические белки РТК связывается с неактивным Ras-белком Активизация Ras-белка: замена ГДФ на ГТФ Ключевой компонент сигнального пути, связанного с рецепторной тирозинкиназой – Ras-белок.
 Ras-БЕЛОК – МОНОМЕРНАЯ ГТФаза В АКТИВНОМ СОСТОЯНИИ Ras-БЕЛОК КОНТАКТИРУЕТ С ГТФ, В НЕАКТИВНОМ – С ГДФ АКТИВАЦИЯ И ИНАКТИВАЦИЯ Rаs-БЕЛКА ОСУЩЕСТВЛЯЕТСЯ ДВУМЯ СПЕЦИФИЧЕСКИМИ БЕЛКАМИ (GEF и GAP) ПОСЛЕ АКТИВАЦИИ Ras-БЕЛКА В ПРОЦЕСС ВКЛЮЧАЕТСЯ НЕСКОЛЬКО ВНУТРИКЛЕТОЧНЫХ КИНАЗ (КИНАЗНЫЙ КАСКАД) ЭФФЕКТ – ДЕЛЕНИЕ КЛЕТОК
Ras-БЕЛОК – МОНОМЕРНАЯ ГТФаза В АКТИВНОМ СОСТОЯНИИ Ras-БЕЛОК КОНТАКТИРУЕТ С ГТФ, В НЕАКТИВНОМ – С ГДФ АКТИВАЦИЯ И ИНАКТИВАЦИЯ Rаs-БЕЛКА ОСУЩЕСТВЛЯЕТСЯ ДВУМЯ СПЕЦИФИЧЕСКИМИ БЕЛКАМИ (GEF и GAP) ПОСЛЕ АКТИВАЦИИ Ras-БЕЛКА В ПРОЦЕСС ВКЛЮЧАЕТСЯ НЕСКОЛЬКО ВНУТРИКЛЕТОЧНЫХ КИНАЗ (КИНАЗНЫЙ КАСКАД) ЭФФЕКТ – ДЕЛЕНИЕ КЛЕТОК
 Каскад внутриклеточных протеинкиназ, фосфорилирующих многие субстраты, в т. ч. и факторы транскрипции
Каскад внутриклеточных протеинкиназ, фосфорилирующих многие субстраты, в т. ч. и факторы транскрипции
 МЕХАНИЗМ ДЕЙСТВИЯ ИНСУЛИНА
МЕХАНИЗМ ДЕЙСТВИЯ ИНСУЛИНА
 МЕМБРАННОСВЯЗАННАЯ ГУАНИЛАТЦИКЛАЗА АГОНИСТЫ: Na-УРЕТИЧЕСКИЙ ПЕПТИДЫ, СЕКРЕТИРУЕМЫЕ ЯИЧНИКАМИ И СТИМУЛИРУЮЩИЕ ПОДВИЖНОСТЬ СПЕРМАТОЗОИДОВ
МЕМБРАННОСВЯЗАННАЯ ГУАНИЛАТЦИКЛАЗА АГОНИСТЫ: Na-УРЕТИЧЕСКИЙ ПЕПТИДЫ, СЕКРЕТИРУЕМЫЕ ЯИЧНИКАМИ И СТИМУЛИРУЮЩИЕ ПОДВИЖНОСТЬ СПЕРМАТОЗОИДОВ
 РЕЦЕПТОРЫ, НЕ ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ, НО СОПРЯЖЕННЫЕ С ТИРОЗИНКИНАЗОЙ
РЕЦЕПТОРЫ, НЕ ПРОЯВЛЯЮЩИЕ ФЕРМЕНТАТИВНУЮ АКТИВНОСТЬ, НО СОПРЯЖЕННЫЕ С ТИРОЗИНКИНАЗОЙ
 Эти рецепторы при активации связываются с цитоплазматическими тирозинкиназами и образуют сигнальный комплекс. Ø Рецепторы цитокинов Ø Рецепторы антигенов на Т- и Влимфоцитах Ø Fc – рецепторы
Эти рецепторы при активации связываются с цитоплазматическими тирозинкиназами и образуют сигнальный комплекс. Ø Рецепторы цитокинов Ø Рецепторы антигенов на Т- и Влимфоцитах Ø Fc – рецепторы
 ØИнтерфероны (IFNs) ØКолониестимулирующие факторы (CSFs ) ØФактор некроза опухоли") АГОНИСТЫ – ЦИТОКИНЫ ØИнтерлейкины (IL) ØИнтерфероны (IFNs) ØКолониестимулирующие факторы (CSFs ) ØФактор некроза опухоли (TNF)
АГОНИСТЫ – ЦИТОКИНЫ ØИнтерлейкины (IL) ØИнтерфероны (IFNs) ØКолониестимулирующие факторы (CSFs ) ØФактор некроза опухоли (TNF)
 Цитокины — группа гормоноподобных белков и пептидов синтезируются и секретируются клетками иммунной системы и другими типами клеток. ФУНКЦИИ цитокинов: Øуправляют развитием и гомеостазом иммунной системы Øосуществляют контроль за ростом и дифференцировкой клеток крови (системой гемопоэза) Øпринимают участие в неспецифических защитных реакциях организма, оказывая влияние на воспалительные процессы, свертывание крови, кровяное давление Ø принимают участие в регуляции роста, дифференцировки и продолжительности жизни клеток, а также в управлении апоптозом
Цитокины — группа гормоноподобных белков и пептидов синтезируются и секретируются клетками иммунной системы и другими типами клеток. ФУНКЦИИ цитокинов: Øуправляют развитием и гомеостазом иммунной системы Øосуществляют контроль за ростом и дифференцировкой клеток крови (системой гемопоэза) Øпринимают участие в неспецифических защитных реакциях организма, оказывая влияние на воспалительные процессы, свертывание крови, кровяное давление Ø принимают участие в регуляции роста, дифференцировки и продолжительности жизни клеток, а также в управлении апоптозом

 ОСНОВНЫЕ ИСТОЧНИКИ И ЭФФЕКТЫ НЕКОТОРЫХ ЦИТОКИНОВ
ОСНОВНЫЕ ИСТОЧНИКИ И ЭФФЕКТЫ НЕКОТОРЫХ ЦИТОКИНОВ
 ЦИТОКИНОВЫЙ РЕЦЕПТОР
ЦИТОКИНОВЫЙ РЕЦЕПТОР
 Эти рецепторы не имеют киназного участка в своем составе и фосфорилируются особыми цитоплазматическими киназами (JANUS KINASES - JAKs). Расположены на внутренней поверхности мембраны вблизи рецептора. JAKs фосфорилируют остатки тирозина как на рецепторах, так и на сигнальных белках, связывающихся с рецепторами.
Эти рецепторы не имеют киназного участка в своем составе и фосфорилируются особыми цитоплазматическими киназами (JANUS KINASES - JAKs). Расположены на внутренней поверхности мембраны вблизи рецептора. JAKs фосфорилируют остатки тирозина как на рецепторах, так и на сигнальных белках, связывающихся с рецепторами.
 МЕХАНИЗМ ДЕЙСТВИЯ ЦИТОКИНОВЫХ РЕЦЕПТОРОВ 1 – СВЯЗЫВАНИЕ РЕЦЕПТОРА С ЛИГАНДОМ 2 – СТИМУЛЯЦИЯ ДИМЕРИЗАЦИИ БЕЛКА-ПЕРЕНОСЧИКА СИГНАЛА. ЭТИ БЕЛКИ ПЕРЕДАЮТ СИГНАЛ НА ТИРОЗИНКИНАЗУ 3 –АКТИВАЦИЯ ТИРОЗИНКИНАЗЫ (ЯНУС-КИНАЗЫ, ИМЕЮЩЕЙ 2 АКТИВНЫХ ЦЕНТРА) 4 – ФОСФОРИЛИРОВАНИЕ ФАКТОРОВ ТРАНСКРИПЦИИ (STAT) 5 – ФАКТОР ПЕРЕХОДИТ В АКТИВНУЮ ФОРМУ И ОБРАЗУЕТ ДИМЕР STAT – фактор транскрипции (signal transducers and activation of transcription) 6 – ТРАНСЛОКАЦИЯ ДИМЕРА В ЯДРО И ИНДУКЦИЯ ТРАНСКРИПЦИИ
МЕХАНИЗМ ДЕЙСТВИЯ ЦИТОКИНОВЫХ РЕЦЕПТОРОВ 1 – СВЯЗЫВАНИЕ РЕЦЕПТОРА С ЛИГАНДОМ 2 – СТИМУЛЯЦИЯ ДИМЕРИЗАЦИИ БЕЛКА-ПЕРЕНОСЧИКА СИГНАЛА. ЭТИ БЕЛКИ ПЕРЕДАЮТ СИГНАЛ НА ТИРОЗИНКИНАЗУ 3 –АКТИВАЦИЯ ТИРОЗИНКИНАЗЫ (ЯНУС-КИНАЗЫ, ИМЕЮЩЕЙ 2 АКТИВНЫХ ЦЕНТРА) 4 – ФОСФОРИЛИРОВАНИЕ ФАКТОРОВ ТРАНСКРИПЦИИ (STAT) 5 – ФАКТОР ПЕРЕХОДИТ В АКТИВНУЮ ФОРМУ И ОБРАЗУЕТ ДИМЕР STAT – фактор транскрипции (signal transducers and activation of transcription) 6 – ТРАНСЛОКАЦИЯ ДИМЕРА В ЯДРО И ИНДУКЦИЯ ТРАНСКРИПЦИИ
 СВЯЗЫВАНИЕ С ЛИГАНДОМ ВЫЗЫВАЕТ ДИМЕРИЗАЦИЮ РЕЦЕПТОРОВ b) АССОЦИАЦИЯ С ЦИТОЗОЛЬНОЙ ТИРОЗИНКИНАЗОЙ") ЦИТОКИНОВЫЕ РЕЦЕПТОРЫ а) СВЯЗЫВАНИЕ С ЛИГАНДОМ ВЫЗЫВАЕТ ДИМЕРИЗАЦИЮ РЕЦЕПТОРОВ b) АССОЦИАЦИЯ С ЦИТОЗОЛЬНОЙ ТИРОЗИНКИНАЗОЙ с) ЕЕ АКТИВАЦИЯ d)ВЗАИМОДЕЙСТВИЕ С ЦИТОЗОЛЬНЫМИ СУБСТРАТАМИ
ЦИТОКИНОВЫЕ РЕЦЕПТОРЫ а) СВЯЗЫВАНИЕ С ЛИГАНДОМ ВЫЗЫВАЕТ ДИМЕРИЗАЦИЮ РЕЦЕПТОРОВ b) АССОЦИАЦИЯ С ЦИТОЗОЛЬНОЙ ТИРОЗИНКИНАЗОЙ с) ЕЕ АКТИВАЦИЯ d)ВЗАИМОДЕЙСТВИЕ С ЦИТОЗОЛЬНЫМИ СУБСТРАТАМИ
 РЕЦЕПТОРЫ КЛЕТОЧНОЙ АДГЕЗИИ ØИНТЕГРИНЫ ØКАДГЕРИНЫ ØСЕЛЕКТИНЫ
РЕЦЕПТОРЫ КЛЕТОЧНОЙ АДГЕЗИИ ØИНТЕГРИНЫ ØКАДГЕРИНЫ ØСЕЛЕКТИНЫ
, так и с") ИНТЕГРИНЫ взаимодействуют как с внеклеточными молекулами (коллаген, фибронектин и др. ), так и с внутриклеточными белками цитоскелета. участвуют в передаче информации из внеклеточной среды в клетку, а также от внутриклеточных белков через рецептор во внеклеточный матрикс определяют направление дифференцировки клетки, ее форму, митотическую активность, способность к миграции
ИНТЕГРИНЫ взаимодействуют как с внеклеточными молекулами (коллаген, фибронектин и др. ), так и с внутриклеточными белками цитоскелета. участвуют в передаче информации из внеклеточной среды в клетку, а также от внутриклеточных белков через рецептор во внеклеточный матрикс определяют направление дифференцировки клетки, ее форму, митотическую активность, способность к миграции
 Примеры некоторых интегринов: Ø рецепторы для белков внеклеточного матрикса. Они связываются с гликопротеиновыми компонентами внеклеточного матрикса, в частности с фибронектином, ламинином и витронектином ; рецепторы нативного фибронектина имеются на многих клетках, например на фагоцитах, играют важную роль в механизме фиксации клеток на нерастворимых субстратах. Ø интегрины тромбоцитов (IIb и IIIa) участвуют в агрегации тромбоцитов, происходящей при свёртывании крови; Ø лейкоцитарные белки адгезии. Для того чтобы мигрировать к месту инфекции и воспаления, лейкоциты должны вступить во взаимодействие с эндотелиальными клетками сосудов. Это взаимодействие может опосредовать связывание Т-лимфоцитов с фибробластами при воспалении.
Примеры некоторых интегринов: Ø рецепторы для белков внеклеточного матрикса. Они связываются с гликопротеиновыми компонентами внеклеточного матрикса, в частности с фибронектином, ламинином и витронектином ; рецепторы нативного фибронектина имеются на многих клетках, например на фагоцитах, играют важную роль в механизме фиксации клеток на нерастворимых субстратах. Ø интегрины тромбоцитов (IIb и IIIa) участвуют в агрегации тромбоцитов, происходящей при свёртывании крови; Ø лейкоцитарные белки адгезии. Для того чтобы мигрировать к месту инфекции и воспаления, лейкоциты должны вступить во взаимодействие с эндотелиальными клетками сосудов. Это взаимодействие может опосредовать связывание Т-лимфоцитов с фибробластами при воспалении.
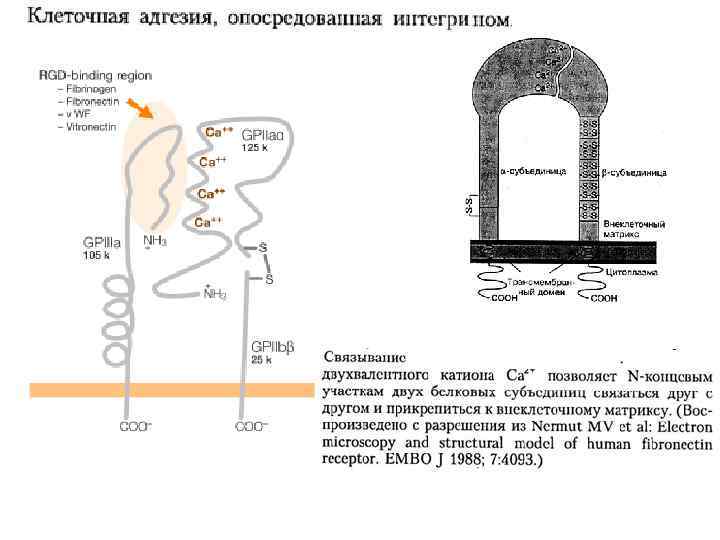
 Рецептор фибронектина. Каждая субъединица имеет Øединственный трансмембранный домен, Øкороткий цитоплазматический Øпротяжённый N-внеклеточный домены. Обе субъединицы (α, β) интегрина гликозилированы и удерживаются вместе нековалентными связями, α-Субъединица синтезируется в виде одной полипептидной цепи, затем расщепляемой на малую трансмембранную цепь и большую внеклеточную цепь, соединённые дисульфидными мостиками. β-Субъединица содержит 4 повтора из 40 аминокислотных остатков каждый. Связываясь с фибронектином снаружи и с цитоскелетом внутри клетки, интегрин действует как трансмембранный линкер.
Рецептор фибронектина. Каждая субъединица имеет Øединственный трансмембранный домен, Øкороткий цитоплазматический Øпротяжённый N-внеклеточный домены. Обе субъединицы (α, β) интегрина гликозилированы и удерживаются вместе нековалентными связями, α-Субъединица синтезируется в виде одной полипептидной цепи, затем расщепляемой на малую трансмембранную цепь и большую внеклеточную цепь, соединённые дисульфидными мостиками. β-Субъединица содержит 4 повтора из 40 аминокислотных остатков каждый. Связываясь с фибронектином снаружи и с цитоскелетом внутри клетки, интегрин действует как трансмембранный линкер.
 с соавт. (2007) разработали концепцию, описывающая участие различных") Британские химики Мартин Хамфриз (Martin Humphries) с соавт. (2007) разработали концепцию, описывающая участие различных типов белков в заживлении ран. Движение новых клеток и заполнение ими полости, вызванной ранением, управляется тем, что клетки прикрепляются и организуются вокруг белковой матрицы фибронектина – внеклеточного белка.
Британские химики Мартин Хамфриз (Martin Humphries) с соавт. (2007) разработали концепцию, описывающая участие различных типов белков в заживлении ран. Движение новых клеток и заполнение ими полости, вызванной ранением, управляется тем, что клетки прикрепляются и организуются вокруг белковой матрицы фибронектина – внеклеточного белка.
, Øплацентарные (Р), Øэпителиальные (Е).") КАДГЕРИНЫ Кадгерины – кальций-зависимые адгезивные молекулы. Три типа: Øнейрональные (N), Øплацентарные (Р), Øэпителиальные (Е). Участвуют в структурной организации тканей (включая лимфоидную), в том числе и эмбриона. Внеклеточные домены этих белков (N, P, E) связывают ионы кальция.
КАДГЕРИНЫ Кадгерины – кальций-зависимые адгезивные молекулы. Три типа: Øнейрональные (N), Øплацентарные (Р), Øэпителиальные (Е). Участвуют в структурной организации тканей (включая лимфоидную), в том числе и эмбриона. Внеклеточные домены этих белков (N, P, E) связывают ионы кальция.
 обеспечивает узнавание клеточных партнёров") Молекулы адгезии в межклеточной коммуникации. Взаимодействие трансмембранных молекул адгезии (кадгерины) обеспечивает узнавание клеточных партнёров и их прикрепление друг к другу (адгезию), что позволяет клеткам-партнёрам формировать щелевые контакты, а также передавать сигналы от клетки к клетке не только при помощи диффундирующих молекул, но и путём взаимодействия встроенных в мембрану лигандов со своими рецепторами в мембране клетки-партнёра
Молекулы адгезии в межклеточной коммуникации. Взаимодействие трансмембранных молекул адгезии (кадгерины) обеспечивает узнавание клеточных партнёров и их прикрепление друг к другу (адгезию), что позволяет клеткам-партнёрам формировать щелевые контакты, а также передавать сигналы от клетки к клетке не только при помощи диффундирующих молекул, но и путём взаимодействия встроенных в мембрану лигандов со своими рецепторами в мембране клетки-партнёра
, участвующих в различных временных") СЕЛЕКТИНЫ: L-селектин Р-селектин Е-селектин семейство углеводосвязывающих белков клеточной поверхности (лектинов), участвующих в различных временных межклеточных адгезионных взаимодействиях в кровяном русле. Внеклеточная часть селектинов состоит из 3 доменов: 1 представлен 2 -9 блоками повторяющихся аминокислотных остатков (комплементрегуляторный белок), 2 Домен эпидермального фактора роста (ЭФР), 3 - N-концевой лектиновый домен
СЕЛЕКТИНЫ: L-селектин Р-селектин Е-селектин семейство углеводосвязывающих белков клеточной поверхности (лектинов), участвующих в различных временных межклеточных адгезионных взаимодействиях в кровяном русле. Внеклеточная часть селектинов состоит из 3 доменов: 1 представлен 2 -9 блоками повторяющихся аминокислотных остатков (комплементрегуляторный белок), 2 Домен эпидермального фактора роста (ЭФР), 3 - N-концевой лектиновый домен
 Взаимодействие клеток крови с эндотелиальными клетками малых кровеносных сосудов. Лейкоцит формирует временные адгезионные контакты с эндотелиальной клеткой. В образовании контактов участвуют белки семейства селектинов: E-селектин на поверхности эндотелиальной клетки, P-селектин на поверхности эндотелиальной клетки и тромбоцита, Lселектин на поверхности многих лейкоцитов.
Взаимодействие клеток крови с эндотелиальными клетками малых кровеносных сосудов. Лейкоцит формирует временные адгезионные контакты с эндотелиальной клеткой. В образовании контактов участвуют белки семейства селектинов: E-селектин на поверхности эндотелиальной клетки, P-селектин на поверхности эндотелиальной клетки и тромбоцита, Lселектин на поверхности многих лейкоцитов.
 Адгезия клеток крови и эндотелия и последующая трансмиграция клеток крови через эндотелий. В адгезии участвуют со стороны эндотелиальных клеток представители надсемейства Ig молекулы ICAM-1 и VCAM-1, со стороны лейкоцитов интегрины VLA-4, LFA-1. В диапедезе лейкоцитов через стенку венул участвует молекула PECAM-1 (CD 31), также принадлежащая надсемейству Ig.
Адгезия клеток крови и эндотелия и последующая трансмиграция клеток крови через эндотелий. В адгезии участвуют со стороны эндотелиальных клеток представители надсемейства Ig молекулы ICAM-1 и VCAM-1, со стороны лейкоцитов интегрины VLA-4, LFA-1. В диапедезе лейкоцитов через стенку венул участвует молекула PECAM-1 (CD 31), также принадлежащая надсемейству Ig.
 КАЛЬЦИЙ-ЗАВИСИМЫЕ ПУТИ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ
КАЛЬЦИЙ-ЗАВИСИМЫЕ ПУТИ РЕГУЛЯЦИИ КЛЕТОЧНОЙ АКТИВНОСТИ
 • Сокращение мышц Роль ионов кальция • Выделение нейромедиаторов • Вторичный посредник Расмуссен, 1970
• Сокращение мышц Роль ионов кальция • Выделение нейромедиаторов • Вторичный посредник Расмуссен, 1970
, действие которых опосредовано ионами кальция.") Известно более 30 БАВ (гормоны, факторы роста, простагландины, цитокины), действие которых опосредовано ионами кальция. Большинство агонистов вызывает повышение внутриклеточной концентрации ионов кальция (например, ангиотензин II, эндотелин I, тромбоксан, брадикинин, гистамин, серотонин и др. ) Гораздо меньше агентов, приводящих к снижению внутриклеточной концентрации ионов кальция (например, простациклин, простагландин Е 1, Na-уретический пептид)
Известно более 30 БАВ (гормоны, факторы роста, простагландины, цитокины), действие которых опосредовано ионами кальция. Большинство агонистов вызывает повышение внутриклеточной концентрации ионов кальция (например, ангиотензин II, эндотелин I, тромбоксан, брадикинин, гистамин, серотонин и др. ) Гораздо меньше агентов, приводящих к снижению внутриклеточной концентрации ионов кальция (например, простациклин, простагландин Е 1, Na-уретический пептид)
 Роль ионов кальция как вторичного посредника • Низкая внутриклеточная концентрация ионов кальция ( 10 -7 М) • Возможность быстро повысить внутриклеточную концентрацию (до 0, 6 – 2 мк. М) ионов кальция
Роль ионов кальция как вторичного посредника • Низкая внутриклеточная концентрация ионов кальция ( 10 -7 М) • Возможность быстро повысить внутриклеточную концентрацию (до 0, 6 – 2 мк. М) ионов кальция
 Характеристики Ca 2+как вторичного посредника 1. В ответ на действие БАВ активируются структуры, которые поставляют ионы кальция в клетку. 2. В ответ на БАВ увеличивается [Ca 2+]in. 3. Снижение активности Ca 2+-АТФазы приводит к усилению эффекта ионов Ca 2+. 4. Эффект БАВ воспроизводится с помощью Ca 2+ионофора. 5. Повышение [Ca 2+]in предшествует биологическому эффекту.
Характеристики Ca 2+как вторичного посредника 1. В ответ на действие БАВ активируются структуры, которые поставляют ионы кальция в клетку. 2. В ответ на БАВ увеличивается [Ca 2+]in. 3. Снижение активности Ca 2+-АТФазы приводит к усилению эффекта ионов Ca 2+. 4. Эффект БАВ воспроизводится с помощью Ca 2+ионофора. 5. Повышение [Ca 2+]in предшествует биологическому эффекту.
 Кальций-связывающие белки I. Кальций-связывающие белки с ферментативной активностью II. Кальций-фосфолипид-связывающие белки III. Кальций-запасающие белки IV. Кальций-связывающие белки в составе ионных каналов и др. ион-транспортных систем V. Регуляторные кальций-связывающие белки, не проявляющие ферментативной активности
Кальций-связывающие белки I. Кальций-связывающие белки с ферментативной активностью II. Кальций-фосфолипид-связывающие белки III. Кальций-запасающие белки IV. Кальций-связывающие белки в составе ионных каналов и др. ион-транспортных систем V. Регуляторные кальций-связывающие белки, не проявляющие ферментативной активности
 I. Кальций-связывающие белки с ферментативной активностью кальпаин Мультисубстратная протеаза кальцинейрин Протеинфосфатаза
I. Кальций-связывающие белки с ферментативной активностью кальпаин Мультисубстратная протеаза кальцинейрин Протеинфосфатаза
 КАЛЬПАИН Гетеродимер: большая каталитическая и малая регуляторная субъединицы. гидролизует белки цитоскелета, ядерные белки, мембранные рецепторы, многие ферменты, транспортные белки
КАЛЬПАИН Гетеродимер: большая каталитическая и малая регуляторная субъединицы. гидролизует белки цитоскелета, ядерные белки, мембранные рецепторы, многие ферменты, транспортные белки
 КАЛЬЦИНЕЙРИН Гетеродимер, состоит из регуляторной и каталитической субъединиц, которые, в свою очередь, имеют несколько доменов. Широко представлена в мозге Дефосфорилирует различные функционально важные фосфобелки по серину и треонину
КАЛЬЦИНЕЙРИН Гетеродимер, состоит из регуляторной и каталитической субъединиц, которые, в свою очередь, имеют несколько доменов. Широко представлена в мозге Дефосфорилирует различные функционально важные фосфобелки по серину и треонину
 Содержат 4 Са 2+-связывающих домена В ПРИСУТСТВИИ") II. Кальций-фосфолипид-связывающие белки АННЕКСИНЫ (известно более 10) Содержат 4 Са 2+-связывающих домена В ПРИСУТСТВИИ ИОНОВ КАЛЬЦИЯ СВЯЗЫВАЮТ ФОСФОЛИПИДЫ МЕМБРАН, ТЕМ САМЫМ ØУЧАСТВУЮТ В ПРОЦЕССАХ СЛИЯНИЯ И АГРЕГАЦИИ МЕМБРАН ØИНГИБИРУЮТ ФОСФОЛИПАЗУ А 2 ØКОНТРОЛИРУЮТ ПРОЛИФЕРАЦИЮ И ДИФФЕРЕНЦИРОВКУ КЛЕТОК
II. Кальций-фосфолипид-связывающие белки АННЕКСИНЫ (известно более 10) Содержат 4 Са 2+-связывающих домена В ПРИСУТСТВИИ ИОНОВ КАЛЬЦИЯ СВЯЗЫВАЮТ ФОСФОЛИПИДЫ МЕМБРАН, ТЕМ САМЫМ ØУЧАСТВУЮТ В ПРОЦЕССАХ СЛИЯНИЯ И АГРЕГАЦИИ МЕМБРАН ØИНГИБИРУЮТ ФОСФОЛИПАЗУ А 2 ØКОНТРОЛИРУЮТ ПРОЛИФЕРАЦИЮ И ДИФФЕРЕНЦИРОВКУ КЛЕТОК
 Во время апоптоза клетки высвобождают фосфатидилсерин на клеточной поверхности. Аннексин V, являющийся фосфолипид-связывающим протеином, в присутствии ионов кальция селективно, с высокой аффинностью, связывает фосфатидилсерин.
Во время апоптоза клетки высвобождают фосфатидилсерин на клеточной поверхности. Аннексин V, являющийся фосфолипид-связывающим протеином, в присутствии ионов кальция селективно, с высокой аффинностью, связывает фосфатидилсерин.
 КАЛЬСЕКВЕСТРИН В поперечно-полосатых мышцах КАЛЬРЕТИКУЛИН") III. Кальций-запасающие белки СИСТЕМА ПАССИВНОГО ЗАБУФЕРИВАНИЯ ИОНОВ КАЛЬЦИЯ (примеры) КАЛЬСЕКВЕСТРИН В поперечно-полосатых мышцах КАЛЬРЕТИКУЛИН В СПР
III. Кальций-запасающие белки СИСТЕМА ПАССИВНОГО ЗАБУФЕРИВАНИЯ ИОНОВ КАЛЬЦИЯ (примеры) КАЛЬСЕКВЕСТРИН В поперечно-полосатых мышцах КАЛЬРЕТИКУЛИН В СПР
 IV. Кальций-связывающие белки в составе ион-транспортных систем Са 2+зависимые калиевые каналы Са 2+зависимые хлорные каналы Na+/Ca 2+ обменник Са 2+ -АТФаза Са 2+
IV. Кальций-связывающие белки в составе ион-транспортных систем Са 2+зависимые калиевые каналы Са 2+зависимые хлорные каналы Na+/Ca 2+ обменник Са 2+ -АТФаза Са 2+
 V. Регуляторные кальций-связывающие белки, не проявляющие ферментативной активности Центральное место среди этих белков занимает КАЛЬМОДУЛИН Впервые обнаружен Ченгом в мозге КРС в качестве Са 2+-зависимого активатора фосфодиэстеразы циклических нуклеотидов. Впоследствии обнаружен во всех эукариотических клетках.
V. Регуляторные кальций-связывающие белки, не проявляющие ферментативной активности Центральное место среди этих белков занимает КАЛЬМОДУЛИН Впервые обнаружен Ченгом в мозге КРС в качестве Са 2+-зависимого активатора фосфодиэстеразы циклических нуклеотидов. Впоследствии обнаружен во всех эукариотических клетках.
 Является “E – F hand”протеином. В структуре белка было обнаружено 6 -спиралей (A, B, C, D, E, F). В петлях полипептидной цепи между участками C и D, E и F находятся функциональные группы с высоким сродством к кальцию. Спирали E и F расположены как вытянутые указательный и большой пальцы - ион Са 2+
Является “E – F hand”протеином. В структуре белка было обнаружено 6 -спиралей (A, B, C, D, E, F). В петлях полипептидной цепи между участками C и D, E и F находятся функциональные группы с высоким сродством к кальцию. Спирали E и F расположены как вытянутые указательный и большой пальцы - ион Са 2+
 Ион кальция
Ион кальция
 Связывающие Са 2+ участки содержат остатки кислых аминокислот и обогащены атомами кислорода. 6 – 8 атомов кислорода образуют ячейку, соответствующую размеру иону кальция (r 0, 099 нм)
Связывающие Са 2+ участки содержат остатки кислых аминокислот и обогащены атомами кислорода. 6 – 8 атомов кислорода образуют ячейку, соответствующую размеру иону кальция (r 0, 099 нм)
 Са-АТФаза Сокращение ГМК Клеточная") Са 2+-кальмодулин-зависимые ферменты процессы Киназа легких цепей миозина (в ГМК) Са-АТФаза Сокращение ГМК Клеточная пролиферация Аденилатциклаза ФДЭ Са-Са. М-зависимая протеинкиназа Диссоциация микротрубочек Высвобождение нейромедиаторов
Са 2+-кальмодулин-зависимые ферменты процессы Киназа легких цепей миозина (в ГМК) Са-АТФаза Сокращение ГМК Клеточная пролиферация Аденилатциклаза ФДЭ Са-Са. М-зависимая протеинкиназа Диссоциация микротрубочек Высвобождение нейромедиаторов
 КОМПЛЕКС КАЛЬЦИЙ-КАЛЬМОДУЛИН ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ С ФЕРМЕНТОМ-МИШЕНЬЮ ЧЕРЕЗ Са 2+-КАЛЬМОДУЛИНЗАВИСИМУЮ ПРОТЕИНКИНАЗУ
КОМПЛЕКС КАЛЬЦИЙ-КАЛЬМОДУЛИН ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ С ФЕРМЕНТОМ-МИШЕНЬЮ ЧЕРЕЗ Са 2+-КАЛЬМОДУЛИНЗАВИСИМУЮ ПРОТЕИНКИНАЗУ
 ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ КОМПЛЕКСА Са 2+ – КАЛЬМОДУЛИН С ФЕРМЕНТОМ-МИШЕНЬЮ
ПРЯМОЕ ВЗАИМОДЕЙСТВИЕ КОМПЛЕКСА Са 2+ – КАЛЬМОДУЛИН С ФЕРМЕНТОМ-МИШЕНЬЮ
 АКТИВАЦИЯ ФЕРМЕНТОВ – МИШЕНЕЙ ЧЕРЕЗ Са 2+-КАЛЬМОДУЛИН-ЗАВИСИМУЮ ПРОТЕИНКИНАЗУ
АКТИВАЦИЯ ФЕРМЕНТОВ – МИШЕНЕЙ ЧЕРЕЗ Са 2+-КАЛЬМОДУЛИН-ЗАВИСИМУЮ ПРОТЕИНКИНАЗУ
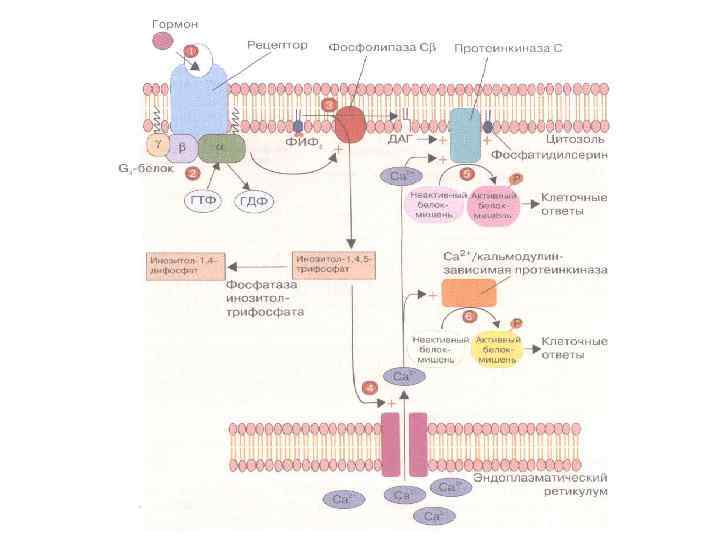
 СИГНАЛЬНАЯ СИСТЕМА, ОПОСРЕДОВАННАЯ ИОНАМИ КАЛЬЦИЯ И ПРОДУКТАМИ ГИДРОЛИЗА МЕМБРАННЫХ ФОСФОИНОЗИТИДОВ
СИГНАЛЬНАЯ СИСТЕМА, ОПОСРЕДОВАННАЯ ИОНАМИ КАЛЬЦИЯ И ПРОДУКТАМИ ГИДРОЛИЗА МЕМБРАННЫХ ФОСФОИНОЗИТИДОВ
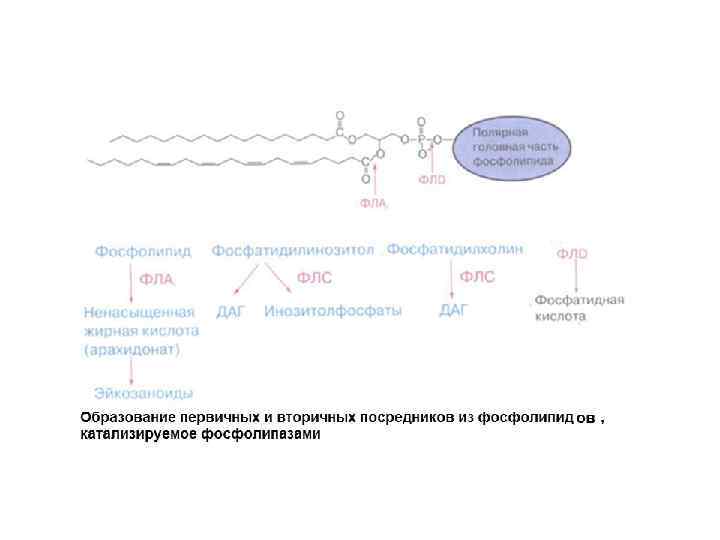
 ВИДЫ ФОСФОЛИПАЗ А PL 1 – гидролиз эфирной связи в положении 1 (отщепляет SN-1 ацильную цепь). А PLС – гидролиз эфирной связи в положении 3 (гидролизует связь между глицериновым остатком фосфолипида и полярной фосфатной группой) PLD - гидролизует связь между фосфатной группой и спиртовой группой PL 2 – гидролиз эфирной связи в положении 2 (отщепляет SN-2 ацильную цепь, образование свободных жирных кислот)
ВИДЫ ФОСФОЛИПАЗ А PL 1 – гидролиз эфирной связи в положении 1 (отщепляет SN-1 ацильную цепь). А PLС – гидролиз эфирной связи в положении 3 (гидролизует связь между глицериновым остатком фосфолипида и полярной фосфатной группой) PLD - гидролизует связь между фосфатной группой и спиртовой группой PL 2 – гидролиз эфирной связи в положении 2 (отщепляет SN-2 ацильную цепь, образование свободных жирных кислот)
 PI-4 -фосфат (PIP) PI-4, 5 -бифосфат (PIP 2)") МЕТАБОЛИЗМ МЕМБРАННЫХ ФОСФОИНОЗИТИДОВ Фосфатидилинозитол(PI) PI-4 -фосфат (PIP) PI-4, 5 -бифосфат (PIP 2)
МЕТАБОЛИЗМ МЕМБРАННЫХ ФОСФОИНОЗИТИДОВ Фосфатидилинозитол(PI) PI-4 -фосфат (PIP) PI-4, 5 -бифосфат (PIP 2)
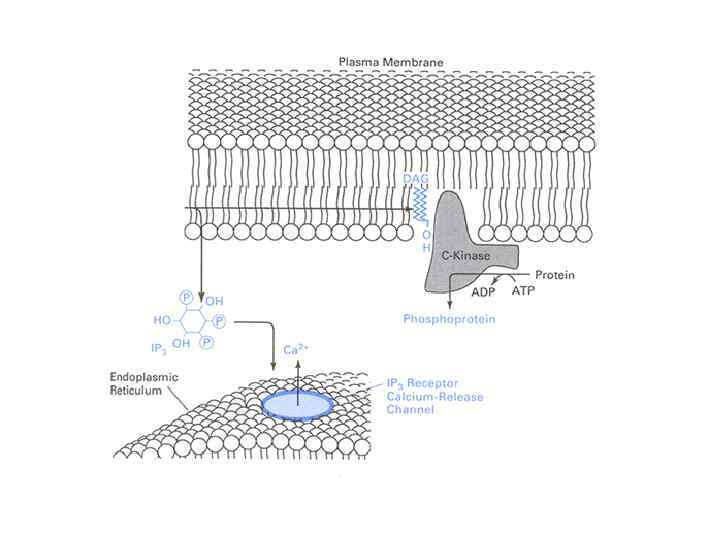
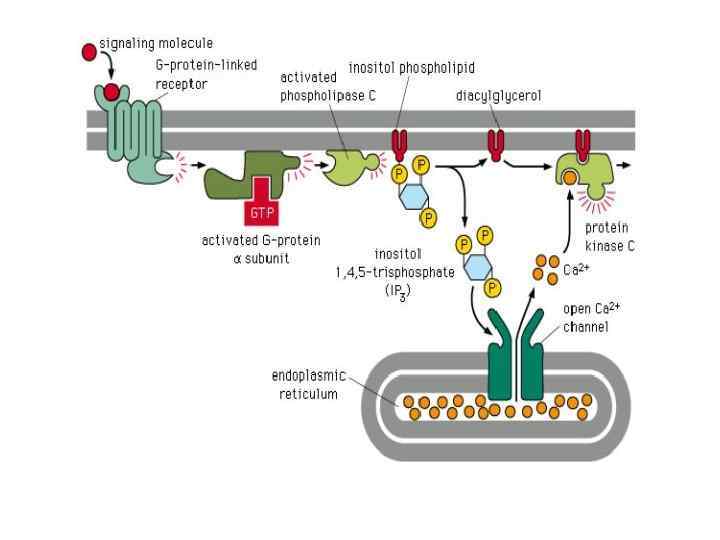
 ПРОДУКТЫ ГИДРОЛИЗА ФОСФОИНОЗИТИДОВ: ВТОРИЧНЫЕ ПОСРЕДНИКИ Инозитолтрифосфат – гидрофильная молекула, которая диффундирует в цитозоль и взаимодействует с рецепторами ретикулума Диацилглицерол – гидрофобная молекула, которая остается в мембране. Под действием фосфолипазы А 2 дает арахидоновую кислоту, которая является предшественником эйкозаноидов.
ПРОДУКТЫ ГИДРОЛИЗА ФОСФОИНОЗИТИДОВ: ВТОРИЧНЫЕ ПОСРЕДНИКИ Инозитолтрифосфат – гидрофильная молекула, которая диффундирует в цитозоль и взаимодействует с рецепторами ретикулума Диацилглицерол – гидрофобная молекула, которая остается в мембране. Под действием фосфолипазы А 2 дает арахидоновую кислоту, которая является предшественником эйкозаноидов.
 ИЗОФОРМЫ ФОСФОЛИПАЗЫ С
ИЗОФОРМЫ ФОСФОЛИПАЗЫ С
 ФОСФОЛИПАЗА") ФОСФОЛИПАЗА С: изоформы и С ФОСФОЛИПАЗА активируется через рецепторы, связанные с G-белками (Gqбелок) ФОСФОЛИПАЗА С активируется благодаря фосфорилированию ее тирозинкиназой.
ФОСФОЛИПАЗА С: изоформы и С ФОСФОЛИПАЗА активируется через рецепторы, связанные с G-белками (Gqбелок) ФОСФОЛИПАЗА С активируется благодаря фосфорилированию ее тирозинкиназой.
 ФОСФОЛИПАЗА С 1 -адренорецепторы, мускариновые рецепторы
ФОСФОЛИПАЗА С 1 -адренорецепторы, мускариновые рецепторы
 ФОСФОЛИПАЗА С Факторы роста
ФОСФОЛИПАЗА С Факторы роста
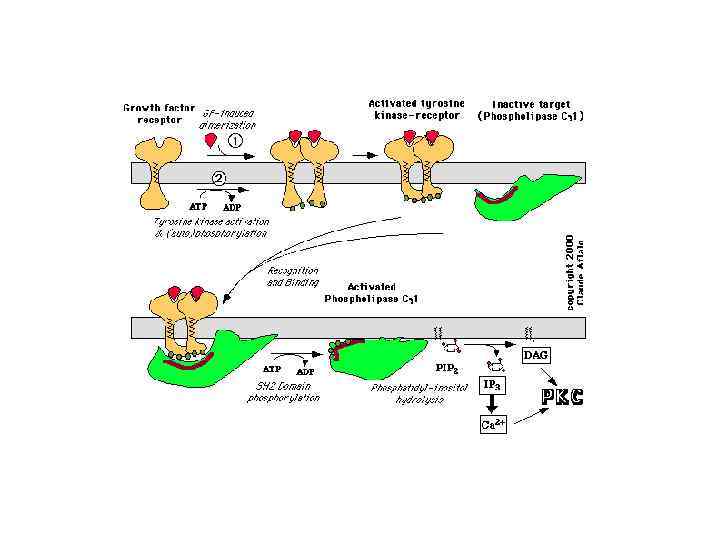
 ПРОЦЕССЫ, В КОТОРЫХ УЧАСТВУЕТ ПРОТЕИНКИНАЗА С Секреция Сокращения ГМК Агрегация тромбоцитов Фосфорилирование рецепторов
ПРОЦЕССЫ, В КОТОРЫХ УЧАСТВУЕТ ПРОТЕИНКИНАЗА С Секреция Сокращения ГМК Агрегация тромбоцитов Фосфорилирование рецепторов
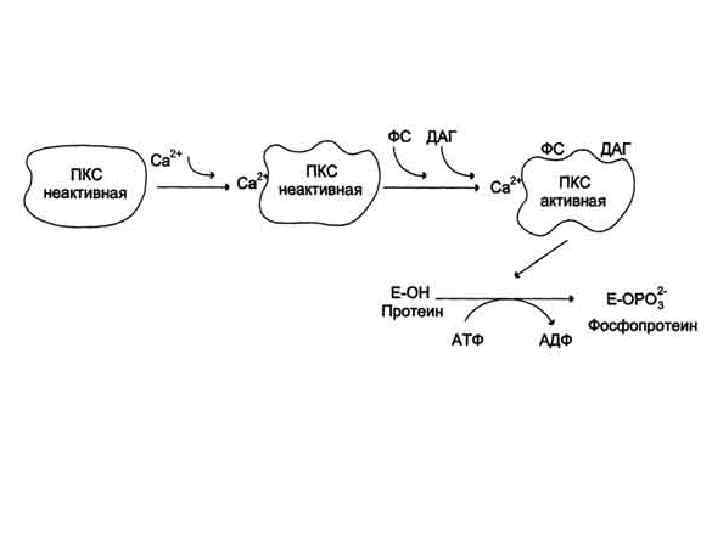
 и каталитического (C) С") Протеинкиназа С R состоит из двух доменов – регуляторного (R) и каталитического (C) С 2 Zn 2+ Каталитический домен: центр, связывающий АТФ и белок-субстрат. Са 2+ АТФ Белоксубстрат Неактивная протеинкиназа С – цитозольная, при активации становится мембранносвязанной ДАГ Регуляторный домен: мотив цинковые пальцы и участок с высоким сродством к Са 2+
Протеинкиназа С R состоит из двух доменов – регуляторного (R) и каталитического (C) С 2 Zn 2+ Каталитический домен: центр, связывающий АТФ и белок-субстрат. Са 2+ АТФ Белоксубстрат Неактивная протеинкиназа С – цитозольная, при активации становится мембранносвязанной ДАГ Регуляторный домен: мотив цинковые пальцы и участок с высоким сродством к Са 2+