37171b61dae21d652c76ab5feb00e83e.ppt
- Количество слайдов: 56



Развитие теории Л-систем Дегерменджи А. Г. Институт биофизики СО РАН 2015

Тезисы: В докладе обобщены собственные теоретические и экспериментальные исследования, связанные с принципами стабильного сосуществования популяций микроорганизмов (бактерий, водорослей, дрожжей), взаимодействующих посредством плотностно-зависимых контролирующих рост факторов (ПКРФ) (в более узком смысле – посредством лимитирующих субстратов) в открытых проточных системах.

В экологии такие регуляторы, которые отвечают, например, за размножение, называются «плотностно-зависимыми контролирующими рост факторами» (ПКРФ), то есть уровень (концентрация) этих факторов зависит от Плотности (биомассы, скорости роста) некоей популяции, и эти Факторы Контролируют Рост данной или другой популяции.
 и особой динамике ПКРФ.")
Доклад посвящен роли ПКРФ в сосуществовании видов (эксперименты, теория, теоремы) и особой динамике ПКРФ.

Структура доклада: - Законы «идеальных» сообществ: -- Принципы сосуществования и стабильности сообществ микроорганизмов в открытых гомогенных экосистемах с учетом ПКРФ [лимитирующих факторов (ЛФ)]. -- Эксперименты по сосуществованию. Управление видовым составом. - Эффект аутостабилизации ЛФ в гомогенных системах. -- Эффект аутостабилизации ЛФ в речных и озерных системах - Множественное лимитирование.

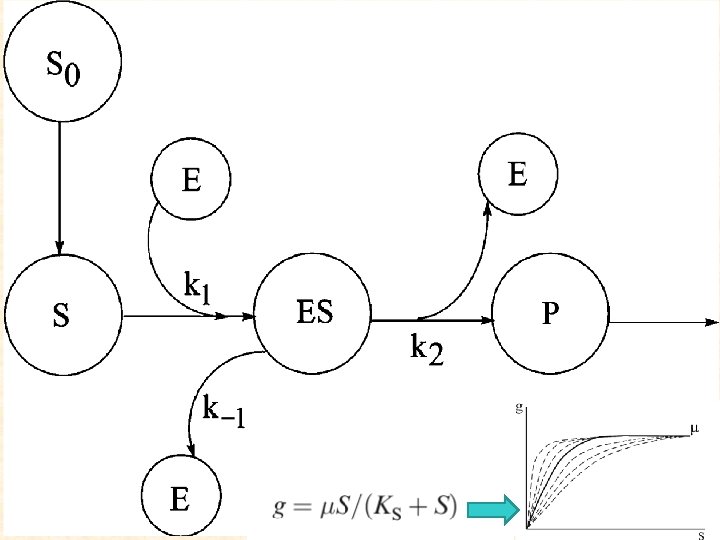
Основы принципа лимитирования Лимитирование роста живых организмов является одной из фундаментальных биологических и экологических концепций и имеет долгую историю обсуждения.

Рост растений Когда рассматривается вопрос лимитирования роста, то чаще всего речь идет о звене продуцентов - растений, водорослей, бактерий, в зависимости от выбранной биологической системы.

Биогенные элементы Но чаще всего при обсуждении лимитировании роста растений идет речь о таких биогенных элементах, как ФОСФОР и АЗОТ, реже упоминая КАЛИЙ, еще реже КАЛЬЦИЙ, МАГНИЙ, НАТРИЙ, ЦИНК и прочие микроэлементы. В первую очередь это связано с тем, какую важную роль биогенные элементы играют в процессах жизнедеятельности организмов и в состав каких органических соединений клеток они входят.
 источник рисунка: https: //ru. wikipedia. org/wiki/Либих,")
Justus von Liebig (1803 – 1897 гг. ) источник рисунка: https: //ru. wikipedia. org/wiki/Либих, _Юстус_фон В 19 столетии Юстус фон Либих сформулировал принцип единственного лимитирующего фактора, который в одном из переводов звучит следующим образом: «Веществом, находящимся в минимуме, управляется урожай и определяются величина и устойчивость последнего во времени» . Часто об этом принципе так же говорят как о «законе минимума» .

Бочка Либиха Всем известно графическое представление этой концепции в виде бочки с водой, у которой клёпки разной длины представляют лимитирующие факторы источник рисунка: https: //ru. wikipedia. org/wiki/Закон_ограничивающего_фактора



ПРИНЦИПЫ СОСУЩЕСТВОВАНИЯ ПОПУЛЯЦИЙ В ОТКРЫТЫХ СИСТЕМАХ Система непрерывной культуры нескольких популяций микроорганизмов с биомассой Хi взаимодействующих через метаболиты Aj.
 сосуществования популяций,")
1. Биофизические законы организации «идеальных» сообществ 1. 1. Установлен общий принцип (теорема) сосуществования популяций, взаимодействующих сколь угодно сложной сетью (априори неизвестной) в смешанной культуре: в стационарной перемешиваемой системе число сосуществующих популяций (Хi) не превышает числа Плотностно-Зависимых Контролирующих Рост Факторов. ПКРФ (Aj), через которые виды взаимодействуют: Принцип строго доказан на основе анализа общего вида системы уравнений, описывающей динамику смеси видов (микроорганизмов): удельная скорость роста (УСР), D – скорость протока, - входной уровень jго ПКРФ, af – скорость трансформации ПКРФ. . m ≤ n. . i=1…m; j=1…n
 с различными механизмами сосуществования")
Проанализировав многочисленные эксперименты (проведенные в ИБФ и описанные в литературе) с различными механизмами сосуществования микробных популяций (без хищников) в гомогенной непрерывной культуре, мы пришли к выводу, что сосуществование основывается на конкуренции, метаболических взаимодействиях и искусственном контроле за счет автоматизированных систем обратной связи (турбидостат, p. H-стат, хлорофилостат и т. п. ).

1. 2. 1. Один из многочисленных экспериментальных примеров сосуществования двух видов дрожжей в хемостате: эксперимент (точки) и теория (расчетные линии).

A Б B

СОСУЩЕСТВОВАНИЕ МИКРОВОДОРОСЛЕЙ Chlorella И Spirulina В РЕЖИМЕ ПРОТОЧНОЙ КУЛЬТУРЫ «ХЛОРОФИЛЛОСТАТ» И ЛИМИТИРОВАНИЯ СВЕТОМ Spirulina Chlorella Spirulina Зависимость УСР (g) монокультур Chlorella and Spirulina от средней облученности клеток E. Динамика доли Spirulina (теория линии) и эксперименты (х, • ), рассчитанные по хлорофиллу “a”, при E 0 = 40 вт/м 2 и поддержании различной оптической плотности C = 1. 6 (a), C = 0. 52 -0. 7 (b), C = 0. 36 (c) по хлорофиллу “a” (680 nm).

Если термин «регулятор» включает также «биотехнологический» фактор, т. е. контроль, осуществляемый при помощи стабилизацию внешнего устройства определенной и функции направленный плотности на вида (турбидостат, хлорофилостат, p. H-стат и т. п. ), то учет количества биотехнологических факторов, t, не меняет критерий сосуществования: m n+t. Критерий остается неизменным и при химическом взаимодействии между факторами или пространственной неоднородности, даже при относительно низкой скорости обменных потоков. Следствием критерия сосуществования является то, что любая подсистема, состоящая из меньшего количества видов (например, из двух), должна контролироваться не менее чем двумя регуляторами и т. д.

1. 2. Известные различные схемы взаимодействий, подтверждающие выведенный принцип сосуществования в открытой системе Sl Nos ПКРФ Сосуществующие виды Граф взаимодействий Ref. 1 2 3 4 5 1 S, glucose, Р, lactate X 1 — Lactobacillus plantarum Х 2 — Proptonibacterium shermanii 2 S, lactate P, acetate X 1 — Desulfovibrio desulfuricans X 2 — Methanobacterium sp. [12] 3 S, manitol P, fructose X 1 — Acetobacter Suboyxdans X 2 — Saccharomyces carlsbergensis [13] 4 S, 5 -methyl-recorcin (orcin) P 1 , unknown product Р 2, unknown product X 1 — Pseudomonas sp. Х 2 — Grampositive bacilli Х 3 — Grampositive bacilli [14] [11]

5 Si —РО 4 ions S 2 - Si. O 2 ions Diatoms X 1 — Cyclotello Х 2 —Asterionella [15] 6 S — parathion (phosphoorganic insecticide) Р 1 — paranitrophenol Р 2 — diethyl phosphate Х 1 — Ps. Stutzeri Х 2 — Ps. Aeruginosa [16] 7 S — oxygen Р 1 — methanol Р 2 — organic matter Р 3 — organic matter X 1 — Pseudomonas sp. Х 2 — Hyphomicrobiит X 3 — Flavobacterium sp. X 4 — Acinetobacter sp. [17] 8 S 1 — n - oxibenzoate S 2 — glucose Хг — Klebsiella aerogenes X 2 — Pseudomonas aeruginosa [18] 9 S — glucose P — riboflavin Xi — Saccharomyces Cerevisiqe X 2 — Lactobacillus casei [19] 10 S — glucose P — vitamin X 1 — Candida mycoderma X 2 - C. tropicalis [20]

ДИАГРАММЫ КОНТРОЛЯ СОСТАВА СООБЩЕСТВА ПОСРЕДСТВОМ ВХОДНЫХ ПОТОКОВ РЕГУЛЯТОРОВ Эксперименты доказали теоретический вывод, что биохимические ПКРФ – метаболиты и лимитирующие рост субстраты – в проточном сообществе являются эффективными «рычагами» контроля видового состава сообщества.
")
1. 2. 3. Управление составом сообществ путем изменения входных концентраций ПКРФ: теория (выделенные области) и опыты ( ) Сосуществование 2 х видов дрожжей и доминирование при изменении ПКРФ: входной глюкозы и автолизата (=витамин В 12). Пунктир – линии равного состава. Принцип сосуществования выполняется: m=2=2=n.
")
1. 2. 3. Управление составом сообществ путем изменения входных концентраций ПКРФ: теория (окрашенные области) и опыты ( ) Сосуществование 2 х видов микроводорослей и доминирование при изменении ПКРФ: суммарной оптической плотности и световой поток (принцип сосуществования выполнен: m=2=2=n). Вывод: а) теория позволяет «создать» сообщества с наперед заданным видовым составом и б) формы областей достаточно сложны и без теории не могут быть построены.
 Критерий классификации взаимодействий (по Одуму) GR 2")
КРИТЕРИИ ВЗАИМОДЕЙСТВИЙ Критерий классификации взаимодействий (по Дегерменджи) Критерий классификации взаимодействий (по Одуму) GR 2 + X X 1 + X 2 + GR + A + + GR 1 0 ? !

1. 4. НОВЫЙ КРИТЕРИЙ МЕЖМИКРОБНЫХ ВЗАИМОДЕЙСТВИЙ – УСКОРЕНИЕ РОСТА В связи с запаздыванием отклика популяции на возмущение был введен новый критерий межмикробного взаимодействия: изменение ускорения роста одной из популяций в ответ на изменение численности другой ( А ). Тангенс угла наклона ( Б ) между касательной к контрольной (невозмущенной) кривой удельной скорости роста и к экспериментальной (возмущенной) – ВЭ и будет характеризовать влияние популяции. Для вывода формулы (Д) воздействия i – популяции на I-ю использовался отклик возмущения ПКРФ ( В ) и контрольный его уровень ( Г ). Итоговая формула удельного (на единицу возмущения) воздействия выглядит по формуле ( Д ): или Б A Д . . В Г

Теоретически, в целом, коэффициент межвидового взаимодействия определяется следующим образом: Сумма справа фактически определяет положительные и отрицательные компоненты общего воздействия l популяции на i популяцию. На основе Bil можно вычислить все коэффициенты межвидового взаимодействия без необходимости решать систему уравнений. Сравнив экспериментальный коэффициент обратной связи (рисунок, Bexp) и теоретический Bii, можно оценить «пробел в знаниях» о регуляции роста монокультур.

Это обеспечило основу для новой классификации взаимодействий в экспериментальных сообществах и для подтверждения отношений между характером взаимодействий (знак и интенсивность) и уровнем входных потоков.
 сезонная динамика коэффициента обратной связи в регуляции бактериопланктона (B) (Дегерменджи, 2007)")
h-2 Теоретическая (расчетная) сезонная динамика коэффициента обратной связи в регуляции бактериопланктона (B) (Дегерменджи, 2007)
 В РЕГУЛЯЦИИ")
1. 4. 1. ПРИМЕР СОПОСТАВЛЕНИЯ ЭКСПЕРИМЕНТАЛЬНЫХ И ТЕОРЕТИЧЕСКИХ КОЭФФИЦИЕНТОВ ОБРАТНОЙ СВЯЗИ (КВ) В РЕГУЛЯЦИИ РОСТА ФИТОПЛАНКТОНА ОЗ. ШИРА (лето, 1998) Дата Вывод: как минимум знак обратной связи (минус) совпадает с теоретическим, но значения теории и опыта пока еще сильно разнятся. Это связано вообще с большой ошибкой дифференциальных показателей.

1. 4. 2. Связь введенных коэффициентов взаимодействий с точностью расчетов динамики численности популяций в сообществе Вопрос: как незнание регуляторов влияет на точность расчетов? Реальный (опыт) с учетом всех ПКРФ (числом n) коэффициент обратной связи (монокультура) как регуляторах (n-k)? как , а теоретический, исходя из знаний о некоторых , то степень незнания регуляторов имеет вид: , а степень знания Доказано, что относительная погрешность расчета биомассы ПРЯМО ПРОПОРЦИОНАЛЬНА СТЕПЕНИ ЗНАНИЯ РЕГУЛЯТОРОВ:
 вне зависимости от физической или биохимической природы")
Выводы : • принцип сосуществования верен: а) вне зависимости от физической или биохимической природы ПКРФ; б) если между ПКРФ есть любые химические реакции; в) если есть две связанные пространственно неоднородные области при достаточном уровне обменных потоков между ними; • если в опытах обнаружено равновесное сосуществование, например, 2 х видов микроорганизмов и один ПКРФ, то с необходимостью надо искать, по крайней мере, еще один ПКРФ; • создание смешанной проточной культуры из заданных m видов с необходимостью требует присутствия в системе не менее чем m ПКРФ, входные потоки которых определяют процентный состав видов (от 0 % до 100%); • добавление в систему параметров (числом t) автоматического регулирования (различные «-статы» ) не меняет принцип: m ≤ n+t; наличие в системе хищников (числом k) также не меняет принцип сосуществования: m ≤ n+k.

1. 5. СУТЬ ЯВЛЕНИЯ АУТОСТАБИЛИЗАЦИИ ЛИМИТИРУЮЩЕГО ФАКТОРА . . при и для не контролирующих факторов при

1. 5. 1. АУТОСТАБИЛИЗАЦИЯ ПКРФ В ОТКРЫТЫХ СИСТЕМАХ Ks = S/ So = 0 В процессе поиска критерия сосуществования видов было обнаружено, что ПКРФ обладают важным парадоксальным свойством – аутостабилизацией, т. е. изменчивость входных потоков ПКРФ не оказывает никакого действия (или очень незначительное) на «фоновые» значения ПКРФ. Мерой такой независимости являются коэффициенты чувствительности (Kj≡∂Aj/∂A 0 j): изменение стационарного уровня каждого ПКРФ на изменение единицы входного уровня ПКРФ. На рисунке показано, что в области, где бактерия Methilomonas sp. лимитируется азотом (область I), значение Ks равно нулю, и действительно, фоновая концентрация азота не зависит от входной (S 0).

1. 5. 2. АУТОСТАБИЛИЗАЦИЯ ПКРФ В СМЕШАННОЙ 2 -Х ВИДОВОЙ ХЕМОСТАТНОЙ КУЛЬТУРЕ Дрожжи – Saccharomyces cerevisiae, бактерии – Lactobacillus casei. ПКРФ: V – рибофлавин, S – глюкоза. Зоны аутостабилизации выделены красным (Megee et al. 1972).

1. 5. 3. ТЕОРЕМА КВАНТОВАНИЯ, ИЛИ СВЯЗЬ ЭФФЕКТА АУТОСТАБИЛИЗАЦИИ ПКРФ С КОЭФФИЦИЕНТАМИ ВЗАИМОДЕЙСТВИЯ Bil или Эта формула получила название теорема «квантования» , поскольку сумма коэффициентов чувствительностей – целое число и может меняться лишь на целое число единиц (если число видов и число ПКРФ «сильно» изменяться). Как следствие теоремы была выведена формула для суммарного изменения биомасс, которая также полезна при анализе целостности сообщества.

Мы назвали эти две корреляции «корреляциями квантования» ; помимо фундаментальной важности, они имеют и прикладное значение. Они могут использоваться для определения полноты списка принимаемых в расчет регуляторов. Теоремы привели к экспериментальному определению количества морфологически неразличимых микробных популяций.

Проявление эффекта аутостабилизации минерального фосфора в соответствии с данными мониторинга состояния 43 озер мира (Janse 2005)
 образует пик на")
АУТОСТАБИЛИЗАЦИЯ В СИСТЕМЕ СЕРНОГО ЦИКЛА ОЗЕРА ШИРА Поток осаждаемой органики (детрита) образует пик на 8 -9 м, который к 14 м формирует пик биомассы сульфатредукторов. С 14 м и глубже наступает лимит по органике и биомасса падает. Теория (равенство УСР и смертности) дает оценку лимита в 0. 25 мг/л, а численный расчет для глубины 14 -21 м дает значение от 0. 15 -0. 17 мг/л, которые близки к 0. 25.
 минерального фосфора за счет окисления")
АУТОСТАБИЛИЗАЦИЯ МИНЕРАЛЬНОГО ФОСФОРА теория наблюдение Апвелинг (поток из глубины) минерального фосфора за счет окисления органики в гиполимнионе не лимитирует фотосинтез вблизи хемоклина (здесь лимит по свету), но, начиная с 9 -8 м, создает лимитирующие условия в фотической зоне и аутостабилизируется.

Вывод: • открыт первый строго формальный закон аутостабилизации, функционально связывающий открытое по обмену веществ стационарное сообщество (микробных популяций), взаимодействующих через физико-химические плотностные факторы; • формула дает экспериментатору точный критерий полноты знаний, как о числе сосуществующих видов, так и числе факторов взаимодействий; • хотя каждый Kj может изменяться «от стационара к стационару» , быть не целым числом с любым знаком (оставаясь, как правило, по модулю меньше единицы) их сумма всегда будет ЦЕЛОЕ число, равное разности числа факторов и числа популяций. Это определяет степень полноты наших знаний о графе взаимодействий в сообществе: если Формула не выполняется для данного сообщества, то значит часть ПКРФ не определена и/или не все популяции обнаружены.
 Несмотря на большое число")
Лимитирование роста – гипотезы, факторы, открытые вопросы (обзор современной литературы) Несмотря на большое число работ, которые продолжают появляться и в настоящее время (поиск в Wo. S показывает около 25000 публикаций, в которых упоминается лимитирование роста), ученые расходятся в понимании механизмов лимитирования, предлагая в многочисленных статьях данные экспериментов и наблюдений, которые подтверждают или опровергают различные точки зрения.

Современная терминология CO-LIMITATION – лимитирование роста организмов несколькими факторами источник : Price N. M. and Morel M. M. , 1991. Colimitation of phytoplankton growth by nickel and nitrogen. Limnol. Oceanogr. 36(6). 1071 -1077.

Elser J. J. et al. , 2007. Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecology Letters. 10. 1135 -1142. В базе анализа находились 653, 243 и 173 натурных эксперимента в пресноводных, морских и наземных экосистемах, соответственно. Один из выводов этой работы заключается в обнаружении синергического эффекта от одновременного добавления фосфора и азота. То есть биомасса или продукция автотрофов увеличиваются сильнее, чем при отдельном добавлении биогенов.
 Одновременное добавление фосфора и азота приводит к тому, что в")
Бочка Либиха (современный вариант) Одновременное добавление фосфора и азота приводит к тому, что в системе происходит частая смена лимита, частое переключение между факторами. То есть фактически речь идет об индивидуальном влиянии каждого биогенного элемента, последовательно сменяющих друга. источник рисунка: Davidson E. A. & Howarth R. W. , 2007. Nutrients in synergy. Nature. 449. 1000– 1001.
 Основное положение MLH: -рост может быть лимитирован несколькими факторами одновременно")
Multiple Limitation Hypothesis (MLH) Основное положение MLH: -рост может быть лимитирован несколькими факторами одновременно источник : Bloom A. J. et al. , 1985. Resource limitation in plants – an economic analogy. Ann. Rev. Ecol. Syst. 16. 363– 392.

Liebig’s law and MLH: equation of growth rate

Liebig’s law and MLH: chart of growth rate источник рисунка: Sperfeld E. et al. , 2012. Multiple resource limitation theory applied to herbivorous consumers: Liebigs minimum rule vs. interactive co-limitation. Ecology Letters. 15. 142– 150.

Sperfeld E. et al. , 2012. Multiple resource limitation theory applied to herbivorous consumers: Liebigs minimum rule vs. interactive co-limitation. Ecology Letters. 15. 142– 150. - изучали зависимость скорости роста Daphnia magna от двух лимитирующих рост факторов: концентрации в корме холестерина и ЕРА (эйкозапентаеновая кислота). - вывод: все модели и подходы одинаково хорошо описывают лабораторный эксперимент на концах осей факторов, но в области «сопоставимых» значений лимитирующих факторов модели построенные на принципе Либиха грубы и переоценивают/преувеличивают значение скорости роста. Наиболее точное совпадение расчетов и лабораторных результатов в этой области получается в моделях, предполагающих одновременное лимитирование роста двумя факторами (MLH).
 1. Переоценка/преувеличение значение скорости роста в области")
Недостатки закона минимума Либиха (из современной литературы) 1. Переоценка/преувеличение значение скорости роста в области «сопоставимых» концентраций лимитирующих факторов. 2. Резкий переход/переключение между лимитирующими факторами.

Egli T. , Zinn M. , 2003. The concept of multiple-nutrient-limited growth of microorganisms and its application in biotechnological processes. Biotechnology Advances. 22. 35 -43. В работе Томаса Эгли и Манфреда Зинна суммируется опыт по хемостатному культивированию микроорганизмов и отмечается режима с двойным или более лимитированием роста биогенными элементами. Подобный режим как правило может существовать в ограниченном диапазоне соотношений биогенных элементов, как показано на рисунке для отношения C: N, и ширина этой зоны двойного лимитирования зависит от условий культивирования, на рисунке - от значения скорости разбавления.

Danger M. et al. , 2008. Does Liebig’s law of the minimum scale up from species to communities? Oikos. 117. 1741 -1751. - применимость закона минимума Либиха на уровне сообществ организмов (Либих сформулировал свою концепцию для индивидуальных растений и только в последствии принцип лимитирующего фактора стали применять шире). - авторы разработали и исследовали модель бактериального сообщества в хемостате, представленную на слайде, и показали, что для сообщества из нескольких видов существует зона одновременного ко-лимитирования роста, что нарушает закон Либиха.

Harpole W. S. et al. , 2011. Nutrient co-limitation of primary producer communities. Ecology Letters. 14. 852 -862. В работе Стенли Харпола с соавторами объединены результаты 641 исследования по ко-лимитированию автотрофного звена в пресноводных, морских и наземных экосистемах. На рисунке представлен результат их систематизации. Площадь каждого прямоугольника соответствует доле в общем числе рассмотренных работ.
 Применение принципа единственного лимитирующего фактора для биологических систем разного уровня организации.")
Открытые вопросы: 1) Применение принципа единственного лимитирующего фактора для биологических систем разного уровня организации. 2) Временной характер действия лимитирующих факторов – последовательно или одновременно? 3) Области единственного и ко- лимитирования в пространстве значений лимитирующих факторов. 4) Множественность типов реакции лимитируемого звена на изменение содержаний биогенных элементов.

Спасибо за внимание!
37171b61dae21d652c76ab5feb00e83e.ppt