Лекция 2-2012 Развитие нервной системы.ppt
- Количество слайдов: 58
 Развитие нервной системы. Лекция - 2 Александрова М. А. ИБР РАН 2012
Развитие нервной системы. Лекция - 2 Александрова М. А. ИБР РАН 2012
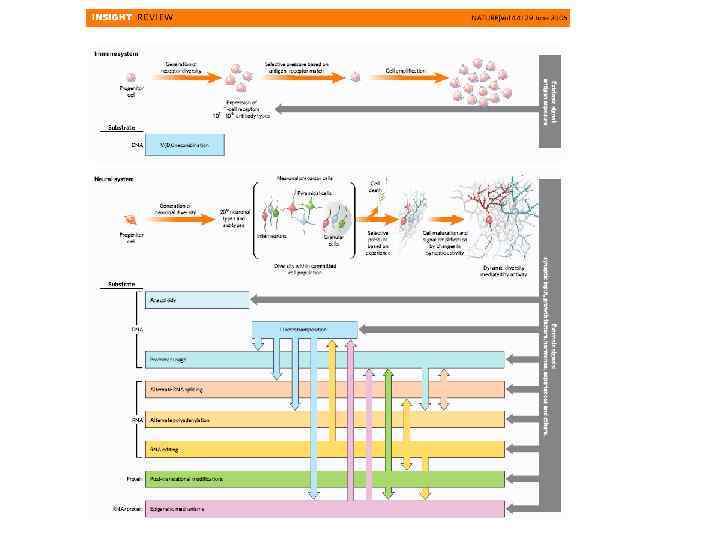
 Целостное поведение животных требует согласованного взаимодействия многих органов и тканей из которых состоит организм. Это согласование осуществляется гуморальными сигналами и нервной системой. Важнейшим отличием действия нервной системы от гуморальной является быстродействие, накопление опыта (научение), способность построения прогноза. Структурно нервная система построена из отдельных блоков (сенсорных и моторных), которые связаны между собой. Нервная система является главной интегрирующей системой организма
Целостное поведение животных требует согласованного взаимодействия многих органов и тканей из которых состоит организм. Это согласование осуществляется гуморальными сигналами и нервной системой. Важнейшим отличием действия нервной системы от гуморальной является быстродействие, накопление опыта (научение), способность построения прогноза. Структурно нервная система построена из отдельных блоков (сенсорных и моторных), которые связаны между собой. Нервная система является главной интегрирующей системой организма
 В основе развития нервной системы лежат два важнейших процесса: Индукция эктодермы Позиционная информация и детерминация Пролиферация клеток Миграция Дифференцировка Апоптоз Рост отростков Формирование связей и синаптогенез Элиминацией избыточных связей Стабилизацией связей В биологии развития нервной системы используют, как модельные объекты: мышь Mus musculus , насекомых Drosophila melanogaster , костистых рыб Danio rerio, лягушек Xenopus laevis , червей Caenorhabditis elegans и др.
В основе развития нервной системы лежат два важнейших процесса: Индукция эктодермы Позиционная информация и детерминация Пролиферация клеток Миграция Дифференцировка Апоптоз Рост отростков Формирование связей и синаптогенез Элиминацией избыточных связей Стабилизацией связей В биологии развития нервной системы используют, как модельные объекты: мышь Mus musculus , насекомых Drosophila melanogaster , костистых рыб Danio rerio, лягушек Xenopus laevis , червей Caenorhabditis elegans и др.
, activin, BMP 2,") Развитие нервной системы позвоночных I. Мезодермальная индукция – Vg 1 (TGF-B), activin, BMP 2, 4, FGF, WNT, Noggin II. Нейральная индукция – noggin, follistatin, chordin
Развитие нервной системы позвоночных I. Мезодермальная индукция – Vg 1 (TGF-B), activin, BMP 2, 4, FGF, WNT, Noggin II. Нейральная индукция – noggin, follistatin, chordin
 Развитие Xenopus Вителлогенин активин
Развитие Xenopus Вителлогенин активин
 BCNE-blastula chordin, noggin expressing center") Сигнальные центры у Xenopus Мозг, базальная пластинка, нотохорд (WNT) BCNE-blastula chordin, noggin expressing center Передняя эндодерма - граница c ЦНС
Сигнальные центры у Xenopus Мозг, базальная пластинка, нотохорд (WNT) BCNE-blastula chordin, noggin expressing center Передняя эндодерма - граница c ЦНС
 Шпемановский организатор источник секретируемых факторов-антагонистов С В области организатора секретируются белки, которые связывают различные факторы роста в межклеточном пространстве и блокируют их нормальный сигналинг через рецепторы.
Шпемановский организатор источник секретируемых факторов-антагонистов С В области организатора секретируются белки, которые связывают различные факторы роста в межклеточном пространстве и блокируют их нормальный сигналинг через рецепторы.
 Ганс Шпеман и Хильда Мангольд – Нобелевская премия по эмбриологии 1935 г.
Ганс Шпеман и Хильда Мангольд – Нобелевская премия по эмбриологии 1935 г.
 Нервная индукция «по умолчанию» - блокада BMP 4 в части клеток эктодермы приводит их к нейрализации Молекулы адгезии E-CAM; E-cadherin Молекулы адгезии N-CAM, N-cadherin
Нервная индукция «по умолчанию» - блокада BMP 4 в части клеток эктодермы приводит их к нейрализации Молекулы адгезии E-CAM; E-cadherin Молекулы адгезии N-CAM, N-cadherin
 Способность мезодермы превращать вышележающую эктодерму в нейральную ткань называется Нейральной индукцией Вдоль передне-задней оси зародыша в нотохорде распределены паракринные факторы, участвующие в индукции. передняя ось задняя ось +FGF Результатом взаимодействия BMP 4 и chordin является активация пронейральных basic. HLH транскрипционных факторов- MASH-1, 2 у млекопитающих.
Способность мезодермы превращать вышележающую эктодерму в нейральную ткань называется Нейральной индукцией Вдоль передне-задней оси зародыша в нотохорде распределены паракринные факторы, участвующие в индукции. передняя ось задняя ось +FGF Результатом взаимодействия BMP 4 и chordin является активация пронейральных basic. HLH транскрипционных факторов- MASH-1, 2 у млекопитающих.
 Сигнальные белки, участвующие в процессе нейральной индукции. TGF-β- семейство трансформирующих факторов роста, в него входят BMP, activin, GDF.
Сигнальные белки, участвующие в процессе нейральной индукции. TGF-β- семейство трансформирующих факторов роста, в него входят BMP, activin, GDF.
 Нейральная индукция.
Нейральная индукция.
 Бластула-гаструла Нервная пластинка Нейрула Нервные валики Нервная трубка
Бластула-гаструла Нервная пластинка Нейрула Нервные валики Нервная трубка
 Экспрессия Chordin в прегаструле ксенопуса Экспрессия Noggin") Паттерны экспрессий ингибиторов BMP (гибридизация in situ) Экспрессия Chordin в прегаструле ксенопуса Экспрессия Noggin в гаструле ксенопуса нотохорд Development - (2000) Regulation of the neural patterning activity of sonic hedgehog by secreted BMP inhibitors expressed by notochord and somites. Karel F. Liem Jr, Thomas M. Jessell* and James Briscoe‡
Паттерны экспрессий ингибиторов BMP (гибридизация in situ) Экспрессия Chordin в прегаструле ксенопуса Экспрессия Noggin в гаструле ксенопуса нотохорд Development - (2000) Regulation of the neural patterning activity of sonic hedgehog by secreted BMP inhibitors expressed by notochord and somites. Karel F. Liem Jr, Thomas M. Jessell* and James Briscoe‡
 и по noggin+chordin (С) возникают разные дефекты в") У нокаутных мышей по noggin (В) и по noggin+chordin (С) возникают разные дефекты в развитии головы. (From Bachiller et al. , 2000)
У нокаутных мышей по noggin (В) и по noggin+chordin (С) возникают разные дефекты в развитии головы. (From Bachiller et al. , 2000)
 Позиционная информация и детерминация
Позиционная информация и детерминация
. Регуляционная") Модели развития нервной системы: Мозаичная модель – полное детерминирование судьбы отдельной клетки (беспозвоночные). Регуляционная модель – случайное и изменяемое развитие отдельных клеток, при детерминированности только нейрального направления (нейрон, глия). Клональная модель – детерминирована судьба отдельных групп клеток, которые являются потомками одного бластомера.
Модели развития нервной системы: Мозаичная модель – полное детерминирование судьбы отдельной клетки (беспозвоночные). Регуляционная модель – случайное и изменяемое развитие отдельных клеток, при детерминированности только нейрального направления (нейрон, глия). Клональная модель – детерминирована судьба отдельных групп клеток, которые являются потомками одного бластомера.
 Развитие клеток бластулы в культуре
Развитие клеток бластулы в культуре
. Am, Al") Клональная модель развития мозга - М. Джекобсон, Г. Хирозе (мечение бластомеров HRP). Am, Al – anterio medial; … lateral Am – v. mesencephalon; v. diencephalon; v. retina; v. telencephalon. Pl Pl Al Pm Аm Al Pm Al - d. mesencephalon; d. diencephalon; d. retina; d. telencephalon. Pm, Pl – posterior medial; . . . lateral. Pm – v. spinal cord; v. rhombencephalon. Pl – d. rhombencephalon; d. spinal cord. Бластула, состоящая из 512 бластомеров. В области анимального полюса 7 клональных компартментов (по 20 клеток) определяют нервную систему еще до момента индукции. В ходе формирования нервной пластинки клетки мигрируют, но не переходят в другие компартменты.
Клональная модель развития мозга - М. Джекобсон, Г. Хирозе (мечение бластомеров HRP). Am, Al – anterio medial; … lateral Am – v. mesencephalon; v. diencephalon; v. retina; v. telencephalon. Pl Pl Al Pm Аm Al Pm Al - d. mesencephalon; d. diencephalon; d. retina; d. telencephalon. Pm, Pl – posterior medial; . . . lateral. Pm – v. spinal cord; v. rhombencephalon. Pl – d. rhombencephalon; d. spinal cord. Бластула, состоящая из 512 бластомеров. В области анимального полюса 7 клональных компартментов (по 20 клеток) определяют нервную систему еще до момента индукции. В ходе формирования нервной пластинки клетки мигрируют, но не переходят в другие компартменты.
 Бластула Гаструла Нейрула Вероятно, процесс детерминации начинается в бластуле, а закрепляется при появление осевых структур и приурочен к стадии поздней гаструлы. Д е т е р м и н а ц и я
Бластула Гаструла Нейрула Вероятно, процесс детерминации начинается в бластуле, а закрепляется при появление осевых структур и приурочен к стадии поздней гаструлы. Д е т е р м и н а ц и я
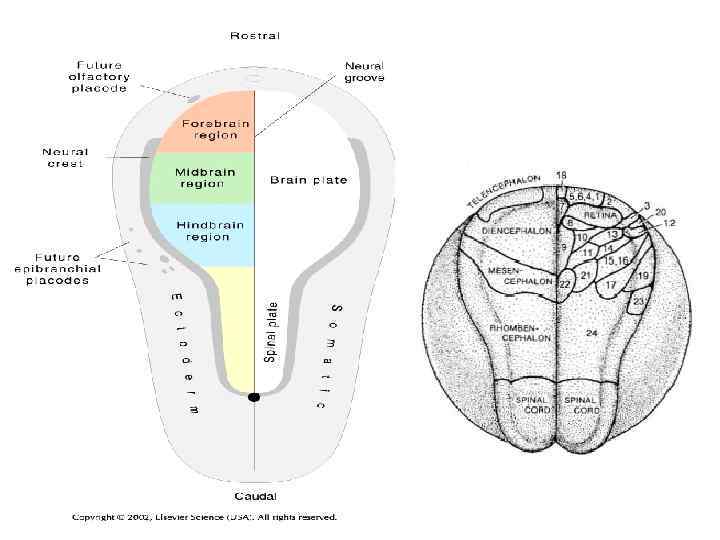
 гены, которые формируют план тела, устанавливая разграничение головного и") Позиционной информацией руководят HOX (гомеобоксные) гены, которые формируют план тела, устанавливая разграничение головного и хвостового отделов.
Позиционной информацией руководят HOX (гомеобоксные) гены, которые формируют план тела, устанавливая разграничение головного и хвостового отделов.
 Нох гены консервативны в эволюционном ряду. Они имеют сходную пространственную организацию и сходный порядок вдоль хромосом
Нох гены консервативны в эволюционном ряду. Они имеют сходную пространственную организацию и сходный порядок вдоль хромосом
 Позиционирование дорзо-вентрального паттерна в нервной трубке индуцируют BMPs, WNT со стороны эпидермиса и Shh (Sonic hedgehog), который экспрессируется в нотохорде. Гомеобоксные гены, участвующие в этих процессах: Nkx 2. 2, Nkx 6. 1, Pax 6, Pax 7, Dbx 1, Dbx 2 и др.
Позиционирование дорзо-вентрального паттерна в нервной трубке индуцируют BMPs, WNT со стороны эпидермиса и Shh (Sonic hedgehog), который экспрессируется в нотохорде. Гомеобоксные гены, участвующие в этих процессах: Nkx 2. 2, Nkx 6. 1, Pax 6, Pax 7, Dbx 1, Dbx 2 и др.
 Вентро-дорзальный градиент Shh определяет тип возникающих нейронов Базальная пластинка Shh Хорда
Вентро-дорзальный градиент Shh определяет тип возникающих нейронов Базальная пластинка Shh Хорда
 Распределение транскрипционных факторов семейства Pax, Mash, Lim, Isl и сигнальных молекул Delta, Serrate, Notch в развивающейся нервной трубке.
Распределение транскрипционных факторов семейства Pax, Mash, Lim, Isl и сигнальных молекул Delta, Serrate, Notch в развивающейся нервной трубке.
 Важную роль в развитии и формировании структур переднего мозга играют регуляторные гены кодирующие транскрипционные факторы Dlx 1, 2, 5 - Distal-less family genes Pax 6 – Paired box gene 6 Otx 1, 2 - Orthodenticle family genes Emx 1, 2 - Empty-spiracles family gene
Важную роль в развитии и формировании структур переднего мозга играют регуляторные гены кодирующие транскрипционные факторы Dlx 1, 2, 5 - Distal-less family genes Pax 6 – Paired box gene 6 Otx 1, 2 - Orthodenticle family genes Emx 1, 2 - Empty-spiracles family gene
 Dlx 1, 2, 5 - Distal-less family genes Dlx гены необходимы для обеспечения тангенциальной миграции интернейронов из субпаллиум в паллиум при развитии мозга позвоночных. Мыши без Dlx проявляют электрофизиологические и гистологические доказательства эпилепсии. В ростральном переднем мозге Dlx ген связан с тормозными ГАМК нейронами. паллиум субпаллиум
Dlx 1, 2, 5 - Distal-less family genes Dlx гены необходимы для обеспечения тангенциальной миграции интернейронов из субпаллиум в паллиум при развитии мозга позвоночных. Мыши без Dlx проявляют электрофизиологические и гистологические доказательства эпилепсии. В ростральном переднем мозге Dlx ген связан с тормозными ГАМК нейронами. паллиум субпаллиум
 Экспрессия гена otx 2 в развивающемся мозгу цыпленка. Otx 2 игает важную роль в развитии мозга и сенсорных органов. Otx 2 последовательно маркирует структуры переднего мозга, но не выявляется в задних отделах мозга.
Экспрессия гена otx 2 в развивающемся мозгу цыпленка. Otx 2 игает важную роль в развитии мозга и сенсорных органов. Otx 2 последовательно маркирует структуры переднего мозга, но не выявляется в задних отделах мозга.
 Otx 2 ген в развитии переднего мозга и передней части головы. Otx 2 уже экспрессируется в эпибласте у Е 5. 5 мышиных эмбрионов. Позднее его экспрессия распределяется в большинстве передних областей нейроэктодермы, в той части, которая предопределяет передний и средний мозг. Делеция гена otx 2 у мышей приводит к утрате передних отделов, хотя сохраняются структуры заднего мозга (анэнцефалия)
Otx 2 ген в развитии переднего мозга и передней части головы. Otx 2 уже экспрессируется в эпибласте у Е 5. 5 мышиных эмбрионов. Позднее его экспрессия распределяется в большинстве передних областей нейроэктодермы, в той части, которая предопределяет передний и средний мозг. Делеция гена otx 2 у мышей приводит к утрате передних отделов, хотя сохраняются структуры заднего мозга (анэнцефалия)
,") Emx 2 экспрессия появляется в вентрикулярной герминальной зоне, ранний маркер неокортекса (Е 8. 5), играет роль в кортикальном нейрогенезе. Emx 2 экспрессируется в основном во всех кортикальных нейронах, которые мигрируют, пролиферируют, дифференцируются и уже дифференцированных и затем исчезает.
Emx 2 экспрессия появляется в вентрикулярной герминальной зоне, ранний маркер неокортекса (Е 8. 5), играет роль в кортикальном нейрогенезе. Emx 2 экспрессируется в основном во всех кортикальных нейронах, которые мигрируют, пролиферируют, дифференцируются и уже дифференцированных и затем исчезает.
 Pax 6 – Paired box gene 6 играет важную роль в развитии НС, «мастер ген» для развития глаза и др. Участие транскрипционного фактора Pax 6 в индукции оптических и назальных структур у зародыша крысы WT PAX-/-
Pax 6 – Paired box gene 6 играет важную роль в развитии НС, «мастер ген» для развития глаза и др. Участие транскрипционного фактора Pax 6 в индукции оптических и назальных структур у зародыша крысы WT PAX-/-
 Pax 6 в развитии разных структур мозга
Pax 6 в развитии разных структур мозга
 Транскрипционные факторы Emx 2 и Pax 6 определяют специфичность представительства сенсорных модальностей по коре мозга. Градиенты распределения факторов Дикий тип
Транскрипционные факторы Emx 2 и Pax 6 определяют специфичность представительства сенсорных модальностей по коре мозга. Градиенты распределения факторов Дикий тип
 Сигнальная система Notch/Delta неоходима для установления самоподдержания в стволовых и прогенитрных клетках. На эктодерме дрозофилы установлено, что Notch/Delta сигнальная система определяет дифференцировку нейроэпителиальных клеток в нейробласты. Судьба клетки зависит от уровня активности Notch рецепторов. У мутантов Notch-/- все пронейральные клетки развиваются в нейробласты.
Сигнальная система Notch/Delta неоходима для установления самоподдержания в стволовых и прогенитрных клетках. На эктодерме дрозофилы установлено, что Notch/Delta сигнальная система определяет дифференцировку нейроэпителиальных клеток в нейробласты. Судьба клетки зависит от уровня активности Notch рецепторов. У мутантов Notch-/- все пронейральные клетки развиваются в нейробласты.
 Numb ингибитор Notch. Ассиметричное распределение Numb определяет судьбу клетки при делении стволовых нейроэпителиальных клеток и нейробластов неокортекса. Q. Shen et al. , Dev. 2002.
Numb ингибитор Notch. Ассиметричное распределение Numb определяет судьбу клетки при делении стволовых нейроэпителиальных клеток и нейробластов неокортекса. Q. Shen et al. , Dev. 2002.
 Соответствующие по времени воздействия комплексов специфических молекулярных факторов, являются критичными для правильного развития структур мозга и дифференцировки отдельных нервных клеток.
Соответствующие по времени воздействия комплексов специфических молекулярных факторов, являются критичными для правильного развития структур мозга и дифференцировки отдельных нервных клеток.
 У млекопитающих нервные клетки формируются из двух источников: 1. Нервной трубки 2. Ганглиозной пластинки (нервного гребня)
У млекопитающих нервные клетки формируются из двух источников: 1. Нервной трубки 2. Ганглиозной пластинки (нервного гребня)
 Из нервной трубки формируется головной и спинной мозг.
Из нервной трубки формируется головной и спинной мозг.
 Ганглиозная пластинка—нервный гребень. Формируются клетки периферической нервной системы, вегетативные ганглии, клетки мягкой мозговой оболочки, пигментные клетки, хрящевые клетки, мозговое в-во надпочечников
Ганглиозная пластинка—нервный гребень. Формируются клетки периферической нервной системы, вегетативные ганглии, клетки мягкой мозговой оболочки, пигментные клетки, хрящевые клетки, мозговое в-во надпочечников
 Миграция клеток нервного гребня
Миграция клеток нервного гребня
 Развитие центральной нервной системы. Формирование коры головного мозга.
Развитие центральной нервной системы. Формирование коры головного мозга.
 Нервная пластинка и формирование нервных валиков
Нервная пластинка и формирование нервных валиков
 Смыкание нервных валиков и выделение нервной трубки
Смыкание нервных валиков и выделение нервной трубки
 Формирование нервной трубки у эмбриона человека Larsen’s Human Embryology. Смыкание нервных валиков в трубку идет примерно от середины к краям
Формирование нервной трубки у эмбриона человека Larsen’s Human Embryology. Смыкание нервных валиков в трубку идет примерно от середины к краям
 Нервная трубка сложена из нейроэпителия - это псевдостратифицированные эмбриональные клетки, которые дают радиальную глию. basal apical
Нервная трубка сложена из нейроэпителия - это псевдостратифицированные эмбриональные клетки, которые дают радиальную глию. basal apical
 В нервной трубке все митозы происходят в прижелудочковой зоне. Сначала родятся нейроны. Включение Ph. H H 3 (фосфогистон метит только G 2 и М) Включение BRDU (метит S, G 2, M фазы цикла) желудочек мозга Митозы в клетках нервной трубки только в вентрикулярной зоне Транслокация ядер в клетках нервной трубки
В нервной трубке все митозы происходят в прижелудочковой зоне. Сначала родятся нейроны. Включение Ph. H H 3 (фосфогистон метит только G 2 и М) Включение BRDU (метит S, G 2, M фазы цикла) желудочек мозга Митозы в клетках нервной трубки только в вентрикулярной зоне Транслокация ядер в клетках нервной трубки
 Развитие нейронов коры мозга скординоровано в пространстве и времени
Развитие нейронов коры мозга скординоровано в пространстве и времени
") Нейробласты мигрируют по отросткам клеток радиальной глии. Радиальная миграция. Sidman and Rakic (1973)
Нейробласты мигрируют по отросткам клеток радиальной глии. Радиальная миграция. Sidman and Rakic (1973)
. Рилин- гликопротеин, регулирует") Белок Reelin регулирует организацию кортикальной пластинки и миграцию клеток. (Reeler mice). Рилин- гликопротеин, регулирует миграцию и позиционирование нервных клеток в период фетального и раннего постнатального развития. Апое. Е опосредует липопротеиновый метаболизм с его рецептором связывается рилин.
Белок Reelin регулирует организацию кортикальной пластинки и миграцию клеток. (Reeler mice). Рилин- гликопротеин, регулирует миграцию и позиционирование нервных клеток в период фетального и раннего постнатального развития. Апое. Е опосредует липопротеиновый метаболизм с его рецептором связывается рилин.
 Нарушение слоистой организации в ламинарных структурах мозга у мышей Reeler Нейроны Кахала-Ретциуса
Нарушение слоистой организации в ламинарных структурах мозга у мышей Reeler Нейроны Кахала-Ретциуса
 Распределение клеток Кахала-Ретциуса в развивающейся коре мозга
Распределение клеток Кахала-Ретциуса в развивающейся коре мозга
 Латеральная миграция нейробластов из латерального и медиального срединного возвышения. Формирование интернейронов.
Латеральная миграция нейробластов из латерального и медиального срединного возвышения. Формирование интернейронов.
 Э-22 Э-18 МЗ Э-17 КП КП МЗ Э-15 КП МЗ ПЗ ПЗ ПЗ КП СВЗ СВЗ ВЗ
Э-22 Э-18 МЗ Э-17 КП КП МЗ Э-15 КП МЗ ПЗ ПЗ ПЗ КП СВЗ СВЗ ВЗ
 Гетерохронное развитие нейронов коры головного мозга • • • Слой I - E 11 -15 Слой II-III - E 18 -21 Слой IV - E 18 Слой V - E 15 -18 Слой VI - E 12 -16 Развитие клеток в ЦНС подчиняется пространственно-временным градиентам. Первыми дифференцируются нейроны, а затем астроциты и постнатально олигодендроциты.
Гетерохронное развитие нейронов коры головного мозга • • • Слой I - E 11 -15 Слой II-III - E 18 -21 Слой IV - E 18 Слой V - E 15 -18 Слой VI - E 12 -16 Развитие клеток в ЦНС подчиняется пространственно-временным градиентам. Первыми дифференцируются нейроны, а затем астроциты и постнатально олигодендроциты.
 Нервная система развивается проходя ряд последовательных этапов с точной временной последовательностью, которая специфична для каждой нервной структуры.
Нервная система развивается проходя ряд последовательных этапов с точной временной последовательностью, которая специфична для каждой нервной структуры.
 Тотальная генетическая информация в геноме млекопитающих имеет около 105 генов, она не достаточна что бы кодировать межнейрональные связи, которых 1015. В развитие нервной системы вовлечены эпигенетические факторы. Влияния от эмбриона- межклеточные сигналы. Влияния от внешней среды- питание, сенсорный и социальный опыт, обучение и т. д. Эпигенетические факторы могут ИМПРИНТИРОВАТЬСЯ (запечатлеваться) при развитии ЦНС.
Тотальная генетическая информация в геноме млекопитающих имеет около 105 генов, она не достаточна что бы кодировать межнейрональные связи, которых 1015. В развитие нервной системы вовлечены эпигенетические факторы. Влияния от эмбриона- межклеточные сигналы. Влияния от внешней среды- питание, сенсорный и социальный опыт, обучение и т. д. Эпигенетические факторы могут ИМПРИНТИРОВАТЬСЯ (запечатлеваться) при развитии ЦНС.