Биология_6_Размножение организмов.pptx
- Количество слайдов: 32



Размножение организмов

Общие положения. Бесполое размножение • • Размножение или репродукция – присущая всем живым существам функция воспроизведения себе подобных. В отличие от всех других жизненно важных функций организма, размножение направлено не на поддержание жизни отдельной особи, а на сохранение ее генов в потомстве и продолжение рода. В филогенезе у разных групп организмов сформировались, во многих случаях независимо, разные способы размножения, и тот факт, что эти группы выжили и существуют, доказывает эффективность этих способов. Все разнообразие способов размножения можно разделить на два основных типа: бесполое (его вариант – вегетативное) размножение и половое размножение. Бесполое размножение одноклеточных организмов осуществляется путем простого деления клетки надвое. У одноклеточных или колониальных эвкариот, имеющих гетеротрофный тип питания, деление не во всех случаях приводит к образованию клеток полностью идентичных исходной. Нередко при делении они сокращают уровень политении, уменьшая в каждой клетке число копий ДНК. Крайняя простота деления, связанная с относительной простотой организации одноклеточных организмов, позволяет размножаться очень быстро. Так, в благоприятных условиях число бактерий может удваиваться каждые 30 -60 минут; почти что, такую же скорость размножения можно наблюдать и у одноклеточных эукариот (например, у почвенных Actinomiceta).

Размножающийся бесполым путем организм способен бесконечно воспроизводить себя, пока резко не изменятся условия среды обитания или вредная для организма мутация не оборвет данную линию. Нередко бесполому размножению бактерий предшествует образование спор. Бактериальные споры – это покоящиеся клетки с замедленным метаболизмом, окруженные многослойной оболочкой, устойчивые к высыханию и другим неблагоприятным условиям, вызывающим гибель обычных клеток. Спорообразование служит как для переживания таких условий, так и для расселения бактерий: попав в подходящую среду, спора прорастает, превращаясь в вегетативную (делящуюся) клетку. Бесполое размножение с помощью одноклеточных спор свойственно также некоторым грибам и водорослям. Споры в этом случае образуются путем эндомитоза, причем иногда (особенно у грибов) в огромных количествах; при прорастании они воспроизводят организм с уровнем политении кратным материнскому. Некоторые грибы, например фитофтора, образуют подвижные, снабженные жгутиками споры, называемые зооспорами или бродяжками. Проплавав в капельках влаги некоторое время, такая бродяжка "успокаивается", теряет жгутики, покрывается плотной оболочкой и затем, в благоприятных условиях, прорастает. Помимо таких спор, у многих из указанных организмов, а также у всех высших растений формируются споры иного рода, а именно споры, образующиеся путем мейоза. Они содержат гаплоидный набор хромосом и дают начало поколению, обычно не похожему на материнское. (Почкование, например, у губок или вегетативное размножение растений. )

Половое размножение • • Ник Лейн. «Лестница жизни» . Бернард Шоу. “Нобелевским банком спермы” Биохимик Джордж Уолд (гитаристы) Н. Лейн перечисляет многочисленные отрицательные черты полового размножения по сравнению с бесполым, например, с почкованием, свойственным прокариотам, или клонированием, часто наблюдаемым и у прокариот, и у растений. Среди недостатков полового размножения он упоминает: болезни, передащюимися при половых контактах; затрата сил и прочих ресурсов на поиски полового партнёра; вероятность столкнуться с паразитичными генами – есть множество примеров конфликтов между генами, когда паразитические гены убивают все сперматозоиды или даже всех потомков, не получивших их ( «паразитические гены» (ВК)) в наследство, и тому подобное. Добавим еще аргумент, который по мнению остроумного Лейна, демонстрирует преимущество бесполого размножения: «Многие из нас согласятся, что главное волшебство секса (то есть полового размножения) состоит как раз в его способности генерировать изменчивость, всякий раз извлекая из небытия уникальных существ, будто кроликов из шляпы. Но если взглянуть на этот процесс с точки зрения специалистов по математической генетике, окажется, что изменчивость ради изменчивости – это отнюдь не всегда хорошо. Зачем ломать удачную комбинацию? Почему бы просто ее не клонировать?

Многим людям идея клонировать Моцарта или Шоу покажется попыткой “поиграть в Бога”, опасным проявлением непомерной человеческой гордыни, но генетиков смущает не это. Они обращают внимание на несколько более приземленную вещь: бесконечная из-менчивость, порождаемая сексом, может приводить к страданиям, болезням и безвремен-ной смерти, в то время как простое клонирование позволяет от всего этого застраховаться. Благодаря обеспечиваемому клонированием сохранению сочетаний генов, выкованных в горниле отбора, оно часто будет самым надежным выбором» . Однако после всего, выше цитиророванного, Ник Лейн пишет: «Если секс можно считать странной причудой, экзистенциальной нелепостью, то отказ от секса оказывается ещё хуже, ведь он в большинстве случаев приводит к вымиранию. А значит, у секса должны быть и серьёзные преимущества, с лихвой перевешивающие безрассудство этого занятия» . Дальше речь и пойдет об этих преимуществах. Принципиальное отличие полового размножения от бесполого состоит в том, что в нем участвуют обычно два родительских организма, признаки которых перекомбиниру-ются у потомства. Половое размножение свойственно всем эвкариотам, но преобладает оно у животных и высших растений. Переход к этому типу размножения имел огромное значение для эволюции жизни на Земле.

, иллюстрирующая основное преимущество полового размножения. В частности,")
Весьма убедительна аргументация В. Н. Стегния (2013), иллюстрирующая основное преимущество полового размножения. В частности, он пишет: «Очевидно, что эффективность бесполого размножения значительно выше, почему же тогда эукариоты, как правило, размножаются половым путём? Именно потому, что при половом размножении возможна рекомбинация. Но если организмы, размножающиеся половым путём, значительно проигрывают бесполым в эффективности размножения, то рекомбинация должна давать им преимущества, покрывающие этот гигантский проигрыш. В чём же они заключаются? Мёллер показал, что генофонд клонально размножающихся организмов должен медленно, но неуклонно деградировать за счёт последовательного накопления вредных мутаций. Сейчас в научно литературе этот процесс называется храповиком Мёллера. … Мёллер показал, что бесполые популяции, несмотря на давление мутационного процесса, могут поддерживать свое существование за счёт очень высокой численности и сильного давления стабилизирующего отбора, благодаря которому носители даже не очень вредных мутаций быстро погибают, а их место занимают клоны, свободные от мутаций. Однако чем больше у организмов генов, тем больше они накапливают мутаций. Отсюда третье условие, позволяющее виду жить с храповиком Мёллера: малый размер генома и как следствие относительная простота организации. Мощное и радикальное средство борьбы с храповиком Мёллера – рекомбинация. Комбинируя аллели генов при образовании гамет, организм, размножающийся половым путём, может «перегрузить» мутациями одни гаметы и одновременно «недогрузить» другие. В итоге особи, возникшие из перегруженных мутациями гамет, погибают, а продукты гамет, очищенных от мутаций, процветают.

Половой процесс • Многие организмы, размножающиеся бесполым путем, все же «изобрели» ряд спо-собов, с помощью которых они время от времени совершают обмен генетическим матери-алом между двумя клетками одного вида. Такой обмен получил название полового про-цесса. У большинства форм он осуществляется путем конъюгации (соединения). Класси-ческий пример конъюгации демонстрируют инфузории. Две их особи временно соединя-ются ротовыми аппаратами, и между ними образуется цитоплазматический мостик, по которому происходит обмен ядерным материалом. Этому обмену предшествует мейотическое деление нуклеоида. По завершении обмена клетки расходятся и затем размножаются путем деления (митоза, образования гамет). У некоторых бактерий при конъюгации происходит однонаправленный перенос линейной последовательности генов хромосомы от "мужской" клетки (донора) к "женской" (реципиенту), причем величина переносимого фрагмента обычно зависит от времени контакта клеток. Таким образом, половой процесс сводится не к размножению, а к созданию в клетке новых комбинаций генов; собственно размножение происходит бесполым путем.

Половое размножение животных • Многие организмы, размножающиеся бесполым путем, все же «изобрели» ряд спо-собов, с помощью которых они время от времени совершают обмен генетическим матери-алом между двумя клетками одного вида. Такой обмен получил название полового про-цесса. У большинства форм он осуществляется путем конъюгации (соединения). Класси-ческий пример конъюгации демонстрируют инфузории. Две их особи временно соединя-ются ротовыми аппаратами, и между ними образуется цитоплазматический мостик, по которому происходит обмен ядерным материалом. Этому обмену предшествует мейотическое деление нуклеоида. По завершении обмена клетки расходятся и затем размножаются путем деления (митоза, образования гамет). У некоторых бактерий при конъюгации происходит однонаправленный перенос линейной последовательности генов хромосомы от "мужской" клетки (донора) к "женской" (реципиенту), причем величина переносимого фрагмента обычно зависит от времени контакта клеток. Таким образом, половой процесс сводится не к размножению, а к созданию в клетке новых комбинаций генов; собственно размножение происходит бесполым путем.

Переход к половому размножению связан с появлением специализированных половых клеток – мужских и женских гамет, в результате слияния которых (оплодотворения) образуется зигота – клетка, из которой развивается новый организм, обладающий новой комбинацией исходных генетических признаков. Половое размножение впервые появилось у простейших, но переход к нему не был связан с немедленной утратой способности к репродукции бесполым путем: ряд животных сохранили ее, обычно чередуя бесполое размножение с половым. Такое чередование поколений наблюдается у некоторых простейших, кишечнополостных и оболочников. Гаметы и гонады. Основой образования гамет (гаметогенеза) служит мейоз археспоры вследствие чего гаметы, в отличие от других (соматических) клеток организма, гаплоидны. Хотя бесполое размножение клеток посредством деления надвое сохранилось в эволюции как основной механизм роста и развития организма, но репродукция (половое размножение) особей, осуществляется только с участием мейоза. Слияние гамет восстанавливает у зиготы число хромосом до диплоидного. Отметим, что у всех многоклеточных организмов дальнейшие деления клеток зиготы происходят или путем редукции исходной политении, или путем митоза.


У многих простейших в половом размножении участвуют морфологически одинаковые мужские и женские гаметы. Такое явление называется изогамией, и свойственно только одноклеточным. Однако уже у некоторых простейших, например споровиков, и у всех многоклеточных организмов произошла дифференциация гамет: они стали различаться по форме и функции – возникла гетерогамия, т. е. разделение половых клеток на яйца (женские гаметы) и сперматозоиды (мужские гаметы). Большинству животных свойственна оогамия: крупная неподвижная политенная яйцеклетка (яйцо) и мелкий подвижный сперматозоид, за счет активных движений которого происходит его контакт с яйцом, ведущий к оплодотворению. Политения сперматозоида обычно редуцируется еще в семенниках (мужских гонадах) на стадии сперматогенных стволовых клеток. Редукция числа копий ДНК в крупных политенных хромосомах яйцеклеток завершается уже после слияния половых клеток, в эмбриогенезе. Истинное оплодотворение происходит по окончании раннего эмбриогенеза после редукции числа политенных копий молекулы ДНК. У губок и некоторых ресничных червей половые клетки рассеяны в теле и выводятся через разрывы стенки тела или через ротовое отверстие, но у многих плоских червей (а в зачаточной форме – и у гидры) появились гонады – специальные железы, продуцирующие гаметы. Мужские гонады – это семенники, женские – яичники. Правда, у таких гермафродитных животных, как брюхоногие моллюски, мужские и женские половые клетки созревают в одной и той же гонаде, но обычно в разное время, так что гонада функционирует, то как семенник, то как яичник, и самооплодотворения не происходит.

Осеменение. Половые органы обеспечивают продукцию и выделение половых клеток, а тем самым – осеменение, т. е. сближение яиц и сперматозоидов двух особей. Процесс осеменения предшествует оплодотворению – слиянию гамет. Различают два способа осеменения (и соответственно оплодотворения): наружное и внутреннее. При наружном осеменении яйца и сперматозоиды выделяются в воду, где сперматозоиды, активно плавая, могут соединиться с яйцом и произвести оплодотворение. Понятно, что этот способ может быть свойствен только водным (или, как земноводные, сохранившим связь с вод-ной средой) животным, и действительно, он наблюдается у большинства из них. Наружное осеменение обычно не связано со сложным устройством половой системы, хотя у некоторых животных развиваются дополнительные приспособления, например, для сцепления двух особей во время выделения ими половых продуктов. Большую независимость от внешних факторов (в частности, от водной среды) и более экономную продукцию гамет обеспечивает другой способ осеменения – внутреннее, при котором сперматозоиды вводятся непосредственно в женские половые пути. Известен также вариант внутреннего осеменения с помощью сперматофоров – капсул, наполненных сперматозоидами. Такое осеменение называют иногда наружновнутренним. Внутреннее осеменение свойственно ряду водных животных и всем наземным. Оно по-явилось уже на очень ранней ступени эволюции, а именно у плоских червей.

Усложнение половой системы. Переход к внутреннему осеменению и оплодотворению сопровождался усложнением половой системы и формированием дополнительных половых органов. Так, образовались железы, например выделяющие жидкость, в которой находятся сперматозоиды и которая необходима им для движения, или – у самок – формирующие наружную оболочку яиц. У плоских червей и ряда других животных, особенно у насекомых, развились семяприемники для хранения поступающей при осеменении спермы. Поскольку сперматозоиды могут длительно сохранять в них жизнеспособность, наличие семяприемников делает оплодотворение менее зависимым от встречи партнеров: многие насекомые успешно размножаются, спариваясь единственный раз в жизни. Соответственно и время между спариванием и откладкой яиц может варьировать в широких пределах. У самок ряда насекомых (стрекоз, цикад, кузнечиков, наездников и др. ) образовался такой дополнительный половой орган, как яйцеклад, служащий для откладки яиц в ячейки, землю либо в ткани растений или животных. Возникли также копулятивные (совокупительные) органы как приспособление для внутреннего осеменения. У разных групп животных они формировались разным путем. Однако у ряда животных, например многих птиц, специальные копулятивные органы отсутствуют. Если у некоторых яйцекладущих совершенствовался аппарат для откладки яиц, то у животных, перешедших к живорождению, прежде всего у млекопитающих, произошли иные изменения половой системы; самое значительное из них – преобразование среднего отдела яйцевода в матку, где развивается зародыш.

Спаривание. Одним из условий успешного размножения служит одновременное созревание гамет у мужских и женских особей. Некоторые животные способны размножаться круглый год, но у многих, особенно у обитателей средних и высоких широт, размножение сезонное. В этом случае наступление периода размножения зависит от внешних факторов: длины светового дня, температуры воздуха, наличия пищи и т. д. Действие этих факторов на репродуктивную систему, как правило, не прямое, а опосредованное – чаще всего гормонами, регулирующими функциональную активность гонад. Так, у позвоночных с сезонным размножением изменение освещенности влияет на секрецию гормонов гипофиза, "включающих" функцию гонад, а тем самым и определяющих сроки полового размножения. Однако этих физиологических механизмов может быть недостаточно для обеспечения спаривания. В действие часто вступает половой отбор наиболее сильных и приспособленных особей, обычно самцов, способных привлечь самку и отстоять свое право на размножение. Турнирные бои между самцами, ухаживание перед спариванием, охрана своей территории для размножения, так же как, по-видимому, и брачный наряд самцов, – все это средства достижения успеха в размножении самых жизнеспособных особей. Половое поведение достигает большой сложности у высокоорганизованных животных с их развитой нейроэндокринной системой.

Способы воспроизведения потомства. Разные группы животных выработали не только разные способы оплодотворения; у них по-разному появляется на свет потомство. В зависимости от того, как это происходит, различают три способа размножения. Яйцерождение. Подавляющее большинство видов животных откладывают яйца, из которых выводится молодь. Таких животных называют яйцеродящими или яйцекладущими. К ним относятся почти все морские беспозвоночные, насекомые, многие рыбы, земноводные, пресмыкающиеся, птицы и однопроходные млекопитающие. Живорождение. У живородящих животных оплодотворенное яйцо развивается в теле самки, получая от нее питание до самого рождения детеныша на свет. К живородящим относятся все млекопитающие за исключением однопроходных – утконоса и ехидны. Живорождение встречается и в других группах, например у некоторых пресмыкающихся и у более примитивных животных. Яйцеживорождение. Существует и промежуточная форма воспроизведения потомства: яйцо развивается, продолжая оставаться в теле самки, но питание зародыша обеспечивается желтком яйца, а не организмом матери. Яйцеживорождение свойственно некоторым акулам и другим рыбам, ряду земноводных, многим ящерицам и змеям.

Стратегии размножения. С этими способами размножения связаны и разные его стратегии. На одном полюсе – стратегия экономного размножения, характеризующегося медленным воспроизведением малочисленного потомства и заботливым его выращиванием (выкармливанием, уходом, обереганием, научением); на другом – расточительное, избыточное размножение с производством очень большого числа яиц и воспроизведением многочисленного потомства при отсутствии заботы о нем. Если в первом случае вероятность выживания потомства весьма значительна, то во втором шансы на сохранение яиц и выживание каждого отдельного потомка крайне малы, так что только интенсивное размножение может компенсировать высокий процент гибели яиц и молоди на всех стадиях развития. Первая стратегия в целом свойственна высокоорганизованным животным – млекопитающим и птицам. Однако в пределах этих групп основная стратегия может быть выражена в разной степени. Например, у приматов она проявляется в наибольшей мере: они длительно вынашивают плод и производят на свет обычно по одному, еще очень беспомощному, детенышу, которого долго вскармливают и растят. С другой стороны, мыши, крысы или кролики могут плодиться несколько раз в год, рождая каждый раз до десятка детенышей, которые быстро приобретают самостоятельность. В результате такого интенсивного размножения повышается вероятность гибели потомков из-за недостатка пищи или – в связи с быстрым ростом популяции – из-за распространения болезней и размножения хищников.

Партеногенез. Яйца некоторых организмов способны развиваться без оплодотворения, т. е. без участия сперматозоида. Такой процесс однополого размножения называют партеногенезом, или девственным размножением. Его рассматривают как редуцированную форму полового размножения. Примеры естественного партеногенеза у млекопитающих неизвестны; они изредка встречаются у низших позвоночных и весьма обычны у беспозвоночных, особенно у насекомых. Существует два типа партеногенеза: облигатный (т. е. обязательный) и факультативный. Первый свойствен видам, у которых самцов либо нет совсем, либо они редки и не способны функционировать. К таким видам относятся некоторые тли, палочники, сверчки, бабочки; популяции без самцов изредка встречаются у рыб, например у серебряного карася. При факультативном партеногенезе яйца могут развиваться как партеногенетически, так и в результате оплодотворения, причем партеногенетическое размножение может преобладать в условиях, когда слишком редки контакты разнополых особей, например на границе ареала распространения вида. Известен также циклический партеногенез, при котором размножение с участием обоих полов чередуется с партеногенетическим. Например, многие виды тлей дают несколько партеногенетических поколений в течение короткого теплого периода лета, а на зиму откладывают оплодотворенные яйца, которые покрыты плотной оболочкой и способны перезимовывать; весной из них выходят только самки, но осенью появляется поколение с некоторым количеством самцов – и цикл возобновляется.

Половое размножение растений • • • Особенности гаметогенеза у растений. Процесс формирования половых клеток у растений подразделяется на два этапа: 1 -й этап – спорогенез, он завершается образованием гаплоидных клеток – спор; 2 -й этап – гаметогенез, в результате которого образуются зрелые, готовые к оплодотворению, гаметы. Процесс образования микроспор, или пыльцевых зерен, у растений называют микроспорогенезом, а процесс образования мегаспор (или макроспор) – мегаили макроспорогенезом. Процесс гаметогенеза у растений в принципе сходен с таковым у животных, но протекает несколько отличным путем. У растений отсутствует зачатковый путь, т. е. раннее обособление половых клеток. (Пояснение). Микроспорогенез и микрогаметогенез. Рассмотрим микроспорогенез и микрогаметогенез на наиболее общем примере покрытосеменных растений. В субэпидермальной ткани молодого микроспорофилла, называемой археспорием, каждая клетка после ряда делений становится материнской клеткой пыльцы, которая проходит все фазы мейоза.

В результате мейоза клетка археспория даёт четыре гаплоидные микроспоры. Микроспоры лежат четверками, их называют тетрадами спор. При созревании тетрады распадаются на отдельные микроспоры. Этим заканчивается микроспорогенез. Вслед за образованием одноядерной микроспоры начинается микрогаметогенез. Первое митотическое деление микроспоры приводит к образованию вегетативной и генеративной клеток. В дальнейшем вегетативная клетка и ее ядро не делятся. В ней накапливаются запасные питательные вещества, которые в последующем обеспечивают деление генеративной клетки и рост пыльцевой трубки в столбике пестика. Генеративная клетка, содержащая меньшее количество цитоплазмы, вновь делится. Это деление может осуществляться еще в пыльцевом зерне или в пыльцевой трубке. В результате образуются две мужские половые клетки, которые, в отличие от сперматозоидов животных, не способны к движению, так как не имеют жгутиков, и называются спермиями. Таким образом, из одной споры с гаплоидным набором хромосом в результате двух митотических делений образуются три клетки. Две из них спермии и одна вегетативная. Именно после этого микрогаметогенез завершён.
; Б – Схема развития")
Гаметогенез цветковых растений. А – Схема развития мужского гаметофита (микроспорогенез); Б – Схема развития женского гаметофита (макроспорогенез): 1 – микроспора, или материнская клетка, пыльцевого зерна, одна из тетрады; 2 – двуклеточное пыльцевое зерно, вегетативная (крупная) и генеративная (мелкая) клетки; 3 – деление генеративной клетки (образование 2 -х спермиев); 4 – зрелое трехклеточное пыльцевое зерно (спермии-клетки свободно лежат в цитоплазме вегетативной клетки (клетки-трубки)); 5 – прорастание пыльцевого зерна;
; 7 -8 – первое деление")
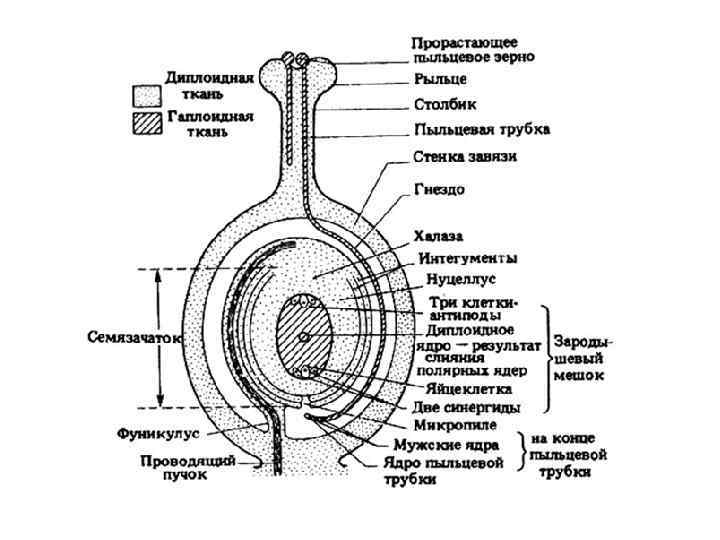
6 – мейоз и образование макроспоры (заштрихованные клетки гибнут); 7 -8 – первое деление гаплоидного ядра макроспоры и начало редукции политении; 9 – второе деление (митоз), четырехъядерная стадия развития женского гаметофита; 10 – третье деление (митоз), восьмиядерная стация; 11 – зрелый восьмиклеточный женский гаметофит (в нем различаются яйцеклетка (более крупная), две синергиды (рядом с яйцеклеткой), две клетки в центре с двумя полярными ядрами и три антиподы); 12 – двойное оплодотворение (слияние спермиев с ядром яйцеклетки и с объединившимися ядрами центральной клетки). Одна из синергид дегенерирует (заштрихована), в ней видны остатки содержимого пыльцевой трубки. Из четырех клеток, располагающихся у микропиле, три клетки – яйцеклетка и две так называемые синергиды образуют яйцевой аппарат. Синергиды играют вспомогательную роль при оплодотворении, они скоро разрушаются. Четвертое ядро отходит к центру зародышевого мешка, где сливается с одним из ядер, отошедшим от халазального конца. Слившиеся в центральной части два гаплоидных ядра образуют одно диплоидное, вторичное, или центральное, ядро зародышевого мешка. Оставшиеся у халазального конца зародышевого мешка три ядра обособляются в клетки; они называются антиподами. Антиподы, так же как и синергиды, играют вспомогательную роль при развитии зиготы и вскоре разрушаются. Таким образом, в результате трех митотических делений в зародышевом мешке об-разуется 8 одинаковых гаплоидных ядер, из которых только одно образует яйцеклетку.

Рассмотренная схема образования восьмиядерного зародышевого мешка из одной мегаспоры является наиболее типичной для покрытосеменных растений. Образование в зародышевом мешке диплоидного центрального ядра, в которое в процессе оплодотворения проникнет один из двух спермиев, образовав триплоидную клетку, которая образует у покрытосеменных растений триплоидный эндосперм, и является одной из основных особенностей, отличающих покрытосеменные растения от голосеменных. У голосеменных – эндосперм гаплоидный. Мегаспоры (или макроспоры) образуются при мега- или макроспорогенезе, и за время от 1 до 4 лет непрерывного роста мегагаметофитов, проходят путь от политенной мегаспоры до зрелых яйцеклеток, готовых к оплодотворению. Микро- и мегаспорогенез у сосны При спорогенезе происходит синтез политенных копий ДНК. У хвойных на образование политенной археспоры затрачивается сезон, при этом возрастает степень политении. У других растений уровень политении меньше, но во всех случаях создание политенных спор требует основных энергетических затрат. Дальнейшее развитие микро- и макрогамет идет в режиме автоматических редукционно-распределительных делений, не требуя дополнительной энергии. Наблюдения М. Г. Романовского, изучавшего политению у сосновых, показали, что выделение археспориальной клетки и образование 1248 копий молекул ДНК в каждой женской хромосоме происходит в первую вегетацию.

Во второй вегетационный период формируются пыльцевые зерна и ценоцитные женские гаметофиты (у Pinus 64 ядерные). Происходит опыление. Пыльца, попав на нуцеллус, прорастает, пыльцевая трубка, достигнув длины 0. 5 -0. 8 мм, закрепляет пыльцевое зерно на нуцеллусе и на год приостанавливает развитие. В третью вегетацию макрогаметофит (женский гаметофит) продолжает деления и переходит к клеточному состоянию; закладываются архегонии и в них выделяются яйцеклетки. Пыльцевая трубка у Pinus sylvestris, начиная с апреля, продолжает рост и в июне происходит оплодотворение. На следующем этапе осуществляется мегагаметогенез. Оставшаяся функционировать мегаспора продолжает расти, и затем ее ядро претерпевает ряд митозов. При этом сама клетка не делится, она образует зародышевый мешок. Дальнейшее развитие эмбриона и редукция политении женской части новообразованной зиготы растягивается до четвертого года, года созревания семян. В северных заполярных районах естественного и интродукционного ареалов видов сосны эмбриогенез не завершается, и семена так и остаются недоразвитыми (Козубов, Данилова, 2006). Сравнение процессов созревания половых клеток у животных и растений показывает почти полный параллелизм их, несмотря на то, что расхождение (дивергенция) или независимое возникновение растений и животных в филогенезе (последнее более вероятно) произошло на очень раннем этапе возникновения клеточной организации.
 и женскими (2,")
Семенное размножение сосны обыкновенной: А – ветка сосны с мужскими (1) и женскими (2, 3) стробилами: 2 – 1 -го года жизни; 3 – 2 -го года жизни после опыления; 4 – зрелая шишка с высыпавшимися семенами; Б – мужская шишка; В – микроспорофилл (5) с 2 пыльцевыми мешками (6); Г – пыльца: 7 – антеридиальная клетка; 8 – вегетативная клетка; 9 – воздушные мешки; 10 – экзина; 11 – интина; Д – прорастание пыльцы: 12 – клетка-ножка; 13 – спермагенная клетка; 14 – пыльцевая трубка; Е – женская шишка: 15 – семенная чешуя с 2 семязачатками; Ж – семязачаток с мегаспорами: 16 – интегумент; 17 – микропиле; 18 – нуцеллус; 19 – мегаспоры; З – семязачаток после развития женского гаметофита: 20 – эндосперм; 21 – архегоний; И – семя: 22 – семенная кожура деревянистая (бывший интегумент); 23 – семенная кожура пленчатая (бывший ну-целлус); 24 – зародыш; 25 – эндосперм; К – семенная чешуя с семенами.

Вегетативное размножение растений • • Вегетати вное размноже ние – образование новой особи из многоклеточной части тела родительской особи, один из способов бесполого размножения, свойственный многоклеточным организмам. У водорослей и грибов происходит путём отделения неспециализированных участков таллома или посредством образования специализированных участков (выводковые почки водоросли). У высших растений происходит либо как распадение материнской особи на две и более дочерние особи (например, при отмирании ползучих побегов или корневищ, отделении корневых отпрысков), либо как отделение от материнской особи зачатков дочерних особей (например, клубни, луковицы, выводковые почки). У некоторых растений могут укореняться отделившиеся от материнского растения побеги (у ивовых) или листья. В основе вегетативного размножения животных лежат процессы, сходные с процессами регенерации; как правило, при отсутствии способности к регенерации у данной группы организмов (например, коловратки, нематоды, пиявки) отсутствует и вегетативное размножение, а при наличии развитой регенерационной способности (кольчатые черви, гидроидные, плоские черви, иглокожие) встречается и вегетативное размножение.

У растений ярко выраженная способность к вегетативному размножению связана с тотипотентностью растительных клеток. Тотипотентность (лат. totus – весь, potentia – сила) – это свойство клетки реализовать генетическую информацию, обеспечивающую ее дифференцировку и развитие до целого организма. Тотипотентностью обладают оплодотворенные яйцеклетка растений и яйцо животных организмов. Что касается дифференцированных клеток, то у животных тотипотентность присуща только некоторым клеткам кишечнополостных. Так, соматические клетки гидры дают начало новому организму. У высших животных с ранних этапов эмбриогенеза, с началом специализации клеток, тотипотентность не реализуется. Однако клетки, изолированные из эмбрионов млекопитающих, в условиях культивирования могут сохранять плюрипотентность – способность дифференцироваться во все типы клеток как собственно зародыша, так и экстраэмбриональных тканей. Такие клетки получили название эмбриональных стволовых клеток. У растений в природных условиях (in vivo) тотипотентность могут проявлять и специализированные клетки. Пример тому – вегетативное размножение, в том числе наблюдаемое в результате развития растений из клеток листьев бегонии, узумбарской фиалки или каланхое. Тотипотентность у растений реализуется и при заживлении ран. В этом случае на раневой поверхности растений в результате неорганизованной пролиферации клеток происходит развитие каллуса (лат. callus – толстая кожа, мозоль). Образование каллуса можно наблюдать прививках в местах срастания привоя и подвоя.

Вегетативное размножение или продление существования особи? • Частичным ответом на поставленный в подзаголовке вопрос служит статья А. Бе-кетова в Энциклопедическом словаре Ф. А. Брокгауза и И. А. Ефрона: «Особи, или так называемые индивиды, в царстве растений далеко не столь резко обособлены друг от друга, как то замечается у высших животных. Поэтому и понятие о долговечности растений не совпадает с тем же понятием в приложении к высшим животным. Дерево, представляющее собой наиболее сложное растение, есть собственно не одна особь, а собрание весьма многих, часто почти бесчисленных, особей, соединенных между собой общим стволом, значительная часть которого уже не живет, а служит лишь общей подпорой. Каждый побег, произошедший из одной единственной почки, может с полным правом считаться особью, что явствует из присущей ему возможности отделяться естественно или искусственно от несущего ствола и давать начало новому растению. Наконец, и сам побег не представляет полной простоты, ибо он состоит из колен, которые нередко могут друг от друга быть отделены (естественно или искусственно) и разрастаться в новую особь. (Упоминаемые в тексте «колена» называются метамерами, то есть отрезками побега, включающими в себя узлы – места прикрепления листа с пазушной почкой, и отрезков стебля между двумя соседними узлами - междоузлиями. У растений с супротивным листорасположением в узле прикрепляются две противостоящие другу почки. ВК)
,")
Таким образом, в царстве растений следует принимать особи разных порядков: сложные (прим. ветвистое дерево), простые (прим. неветвистый побег) и простейшие (колено, или междоузлие). Сказанное должно постоянно иметь в виду при оценке долговечности растений, …. В описательной ботанике, а также и в практике речь идет постоянно о сложной особи. Это получает свой логический смысл в том отношении, что и сама сложная особь имеет, подобно простейшей, свое начало и конец, рождение и смерть. Тут опять замечается глубокое различие между сложнейшими формами растительной и животной жизни. Дело в том, что жизнь сложного животного, протекающая даже при самых благоприятных условиях, заканчивается через определенный период времени по причине постепенного ослабления его физиологических отправлений; напротив того, дерево, при благоприятных условиях, может продолжать свое существование безраздельно, именно вследствие того, что оно представляет собой целую колонию органически связанных между собой простых особей. Так, громадный оротавский драконник (Драцена драконовая Dracaena draco – растение рода драцена. ВК) произрастает в тропиках и субтропиках Африки и на островах Юго-Восточной Азии, которому насчитывалось до 6000 лет, погиб в 1868 г. не от старости, а от сломившей его бури; он до самого конца цвел и приносил плоды. Итак, во внутреннем строении растения наиболее сложного не заключается еще причины смерти. Это свойство ослабляется с упрощением самой особи, причем усиливается ее живучесть. Так, низшие растения могут нередко раздробляться без вреда для продолжения жизни на незначительные участки. Другие, как, например, лишайники, могут сохранять жизненность на долгие годы в совершенно сухом виде.

В приведённой выше длинной цитате, несмотря на некоторую наивность, много правильно подмеченного и, по современным представлениям, несомненного. Предположения о теоретической бессмертности древесных растений в ботанической литературе высказывались неоднократно. В качестве примера можно привести нисколько не устаревшую монографию Ю. В. Гамалея (2004). И на самом деле, древесные растения регулярно самовозобновляюся за счет прироста новых метамеров. Казалось бы, ни какая объективная причина не может приостановить этого процесса. Однако фактически бессмертных растений не бывает, хотя единая конкретная причина, ограничивающая долголетие этих организмов нигде не называется. Среди множества часто упоминаемых причин смерти древесных растений – засуха, вымокание, инвазия патогенных организмов, ураганные ветры и тому подобное. Всё это из области случайных обстоятельств, которые хоть и имеют место когда-то и где-то, но не всегда, не везде и не обязательно. А вот биологически строго обоснованной причины смерти и обоснованных пределов длительности жизни для тех или иных видов древесных растений в литературе не приводится – это остаётся открытым вопросом. На типичных примерах вегетативного размножения растений, в том числе и древесных, мы здесь не останавливаемся – они в избытке приводятся в научнопопулярной и садоводческой литературе. Однако на одном примере, относящемся преимущественно к лесообразующим древесным растениям, считаем нужным остановиться. Речь идет о так называемых капах – шаровидных разрастаниях участков стволов и ветвей деревьев, служащих своеобразными «органами» вегетативного размножения.

Краткое заключение • До настоящего времени отсутствуют даже более или менее убедительные предположения о путях возникновения полового размножения. Не ясно, почему в царстве животных по мере усложнения организации ослабевает способность к бесполому размножению, а в царстве растений наблюдается обратное. Совершенно непонятно, каким образом возник цветок покрытосеменных растений. Число такого рода вопросов и неясностей по мере развития науки о размножении организмов, впрочем, как и других научных направлений, не сокращается, а возрастает. И это вполне закономерно – по мере углубления в какую-либо научную проблему появляются новые, обосабливающиеся ответвления в направлении исследований, и каждое из них привносит свои вопросы и временно неразешимые недоумения.
Биология_6_Размножение организмов.pptx