• Размеры ДНК зависят от типа организма. Физическая длина ДНК вирусов составляет десятки микрометров, бактерий – миллиметры, а человека – 2 метра. Общая длина всех ДНК человека составляет 2∙ 1010 км.
• Размеры ДНК зависят от типа организма. Физическая длина ДНК вирусов составляет десятки микрометров, бактерий – миллиметры, а человека – 2 метра. Общая длина всех ДНК человека составляет 2∙ 1010 км.
 Теоретические предпосылки генетической инженерии • Важнейшие достижения молекулярной биологии
Теоретические предпосылки генетической инженерии • Важнейшие достижения молекулярной биологии
 Термины и определения Ген – единица наследственной информации – участок ДНК, кодирующий один белок или РНК. Геном – совокупность всех генов. Экспрессия генов — это процесс, в котором наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Кодон – триплет нуклеотидов, кодирующий отдельную аминокислоту. Генетический код – соответствие кодонов определенным аминокислотам. Генная инженерия или техника рекомбинантных ДНК – конструирование in vitro (в пробирке) функционально активных генетических структур (рекомбинантных ДНК) путем переноса генетического материала из одного организма в другой. Плазмида - кольцевая двухцепочечная внехромосомная ДНК, способная к автономной репликации. Вектор - самореплицирующаяся (автономная) молекула ДНК, например, бактериальная плазмида, используемая в генной инженерии для переноса генов, часто в некотором числе копий) от организма-донора в организм-реципиент, а также для клонирования нуклеотидных последовательностей (клонирующий вектор). Клон – популяция клеток, идентичных родительской клетке. Клонирование в биологии – это получение точных копий организма или другого объекта, например, клетки или гена.
Термины и определения Ген – единица наследственной информации – участок ДНК, кодирующий один белок или РНК. Геном – совокупность всех генов. Экспрессия генов — это процесс, в котором наследственная информация от гена (последовательности нуклеотидов ДНК) преобразуется в функциональный продукт — РНК или белок. Кодон – триплет нуклеотидов, кодирующий отдельную аминокислоту. Генетический код – соответствие кодонов определенным аминокислотам. Генная инженерия или техника рекомбинантных ДНК – конструирование in vitro (в пробирке) функционально активных генетических структур (рекомбинантных ДНК) путем переноса генетического материала из одного организма в другой. Плазмида - кольцевая двухцепочечная внехромосомная ДНК, способная к автономной репликации. Вектор - самореплицирующаяся (автономная) молекула ДНК, например, бактериальная плазмида, используемая в генной инженерии для переноса генов, часто в некотором числе копий) от организма-донора в организм-реципиент, а также для клонирования нуклеотидных последовательностей (клонирующий вектор). Клон – популяция клеток, идентичных родительской клетке. Клонирование в биологии – это получение точных копий организма или другого объекта, например, клетки или гена.
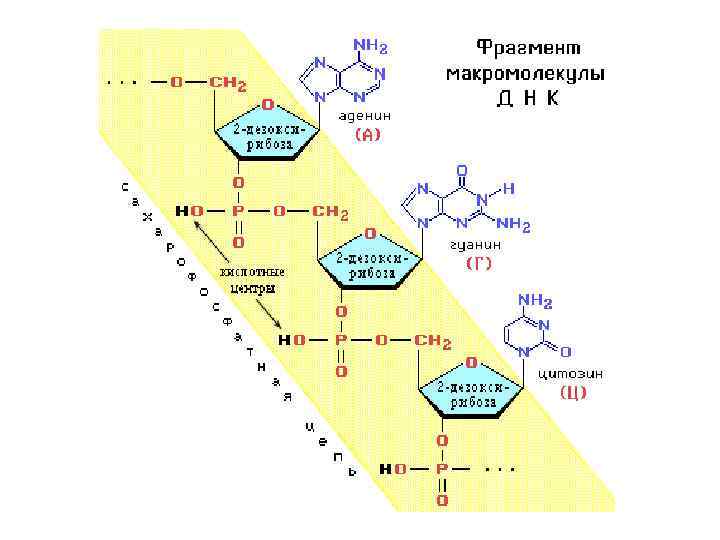
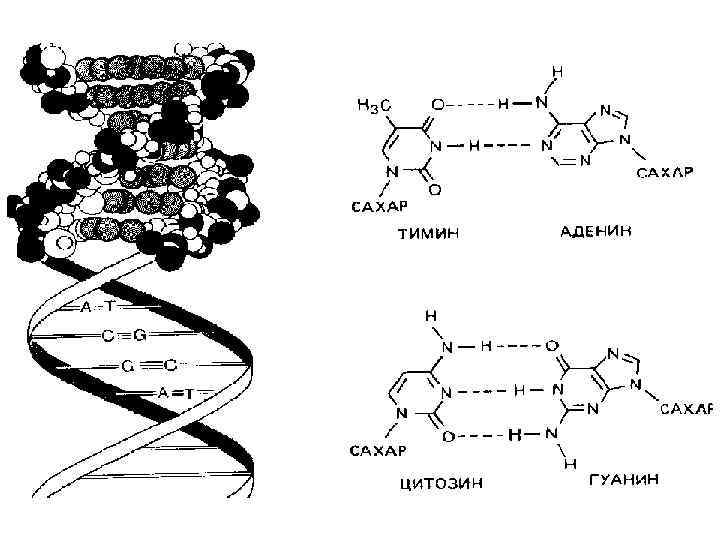
 Молекулярные механизмы матричного синтеза • Процесс удвоения молекулы ДНК называется репликацией ДНК. • Процесс биосинтеза РНК в клетке называется транскрипцией ДНК • Трансляция (м. РНК белок) – многоступенчатый процесс синтеза белка согласно информации, заключенной в последовательности нуклеотидов м. РНК.
Молекулярные механизмы матричного синтеза • Процесс удвоения молекулы ДНК называется репликацией ДНК. • Процесс биосинтеза РНК в клетке называется транскрипцией ДНК • Трансляция (м. РНК белок) – многоступенчатый процесс синтеза белка согласно информации, заключенной в последовательности нуклеотидов м. РНК.
 Репликация ДНК в E. coli Репликация – удвоение ДНК – добавление дезоксинуклеотида к 3’-концу растущей цепи Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований.
Репликация ДНК в E. coli Репликация – удвоение ДНК – добавление дезоксинуклеотида к 3’-концу растущей цепи Хеликаза, топоизомераза и ДНК-связывающие белки расплетают ДНК, удерживают матрицу в разведённом состоянии и вращают молекулу ДНК. Правильность репликации обеспечивается точным соответствием комплементарных пар оснований.
 Репликация – удвоение ДНК – добавление дезоксинуклеотида к 3’-концу растущей цепи
Репликация – удвоение ДНК – добавление дезоксинуклеотида к 3’-концу растущей цепи
 Схема репликации хромосомы E. coli 3, 8 106 п. о. 800 нуклеотидов в сек 40 мин X и Y – две репликативные вилки
Схема репликации хромосомы E. coli 3, 8 106 п. о. 800 нуклеотидов в сек 40 мин X и Y – две репликативные вилки
 Схема репликации хроматина • Хромосомы эукариот представляют собой линейную молекулу ДНК. Эукариотическая ДНК обматывает белковые частицы – гистоны, располагающиеся вдоль ДНК через определённые интервалы, образуя хроматин – волокна, из которых состоят хромосомы. Комплексы участков ДНК и гистонов называются нуклеосомами. Нуклеосомы упорядочены в пространстве, за счёт чего достигается плотная упаковка ДНК в хромосоме.
Схема репликации хроматина • Хромосомы эукариот представляют собой линейную молекулу ДНК. Эукариотическая ДНК обматывает белковые частицы – гистоны, располагающиеся вдоль ДНК через определённые интервалы, образуя хроматин – волокна, из которых состоят хромосомы. Комплексы участков ДНК и гистонов называются нуклеосомами. Нуклеосомы упорядочены в пространстве, за счёт чего достигается плотная упаковка ДНК в хромосоме.
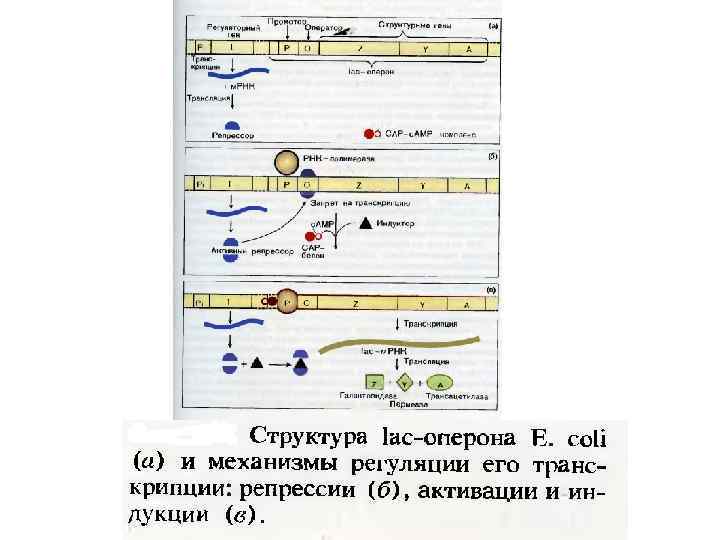
. Транскриптон – участок ДНК,") Транскрипция ДНК-зависимая РНК-полимераза – основной фермент транскрипции (присоединяется к промотору). Транскриптон – участок ДНК, подвергающийся транскрипции. Первичный транскрипт – РНК, образованная в результате транскрипции – комплементарная копия транскриптона от промотора до терминатора (5’→ 3’). Оперон (транскриптон прокариот) – комплекс функционально связанных структурных генов (кодируемые ими белки участвуют в одной биохимической цепи реакций) и оператора.
Транскрипция ДНК-зависимая РНК-полимераза – основной фермент транскрипции (присоединяется к промотору). Транскриптон – участок ДНК, подвергающийся транскрипции. Первичный транскрипт – РНК, образованная в результате транскрипции – комплементарная копия транскриптона от промотора до терминатора (5’→ 3’). Оперон (транскриптон прокариот) – комплекс функционально связанных структурных генов (кодируемые ими белки участвуют в одной биохимической цепи реакций) и оператора.
 Строение транскриптона Прокариоты Промотор - участок оперона, служащий для узнавания ферментом РНК-полимеразой. Последовательность оснований по ходу цепи ДНК ниже сайта промотора с направлением 3’ 5’ используется в качестве матрицы для синтеза РНК. После промотора в опероне находится акцепторная зона (у эукариот) или оператор (у прокариот), которая служит для связывания с регуляторами транскрипции (например, усилителями – энхансерами или репрессорами). Оператор разрешает или запрещает транскрипцию. Цистрон - последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев - белок) или одну т. РНК, или одну р. РНК
Строение транскриптона Прокариоты Промотор - участок оперона, служащий для узнавания ферментом РНК-полимеразой. Последовательность оснований по ходу цепи ДНК ниже сайта промотора с направлением 3’ 5’ используется в качестве матрицы для синтеза РНК. После промотора в опероне находится акцепторная зона (у эукариот) или оператор (у прокариот), которая служит для связывания с регуляторами транскрипции (например, усилителями – энхансерами или репрессорами). Оператор разрешает или запрещает транскрипцию. Цистрон - последовательность нуклеотидов ДНК, кодирующая один полипептид (в большинстве случаев - белок) или одну т. РНК, или одну р. РНК
,") Эукариоты Процессинг включает кэпирование (присоединение к 5’концевому метилированному звену предшественника м. РНК 7 метилгуанозина), полиаденилирование (присоединение к 3’-концу сегмента поли(А)), сплайсинг РНК (вырезание интронов и соединение экзонов – кодирующих последовательностей – в непрерывную последовательность)
Эукариоты Процессинг включает кэпирование (присоединение к 5’концевому метилированному звену предшественника м. РНК 7 метилгуанозина), полиаденилирование (присоединение к 3’-концу сегмента поли(А)), сплайсинг РНК (вырезание интронов и соединение экзонов – кодирующих последовательностей – в непрерывную последовательность)
 Транскрипция РНК
Транскрипция РНК
 Роль малой ядерной РНК в процессе транскрипции
Роль малой ядерной РНК в процессе транскрипции
 Схема процессинга пре-м. РНК в ядре Процессинг – совокупность ферментативных Процессов, в результате которых синтезируемая РНК превращается в функционально полноценную молекулу
Схема процессинга пре-м. РНК в ядре Процессинг – совокупность ферментативных Процессов, в результате которых синтезируемая РНК превращается в функционально полноценную молекулу
 Стадия элонгации трансляции.
Стадия элонгации трансляции.
 Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК.
Генетический код - это система записи информации о последовательности расположения аминокислот в белках с помощью последовательности расположения нуклеотидов в ДНК.
 • - специфичность (один кодон -") Свойства генетического кода • - триплетность (43=64 кодона) • - специфичность (один кодон - одна аминокислота? Исключение составляет кодон AUG. У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин. ) • - вырожденность (dсе аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом. Всего 61 триплет кодирует 20 аминокислот) • - универсальность • - однонаправленность (5’ 3’) • - непрерывность • - неперекрываемость • - линейность
Свойства генетического кода • - триплетность (43=64 кодона) • - специфичность (один кодон - одна аминокислота? Исключение составляет кодон AUG. У прокариот в первой позиции (заглавная буква) он кодирует формилметионин, а в любой другой - метионин. ) • - вырожденность (dсе аминокислоты, за исключением метионина и триптофана, кодируются более чем одним триплетом. Всего 61 триплет кодирует 20 аминокислот) • - универсальность • - однонаправленность (5’ 3’) • - непрерывность • - неперекрываемость • - линейность
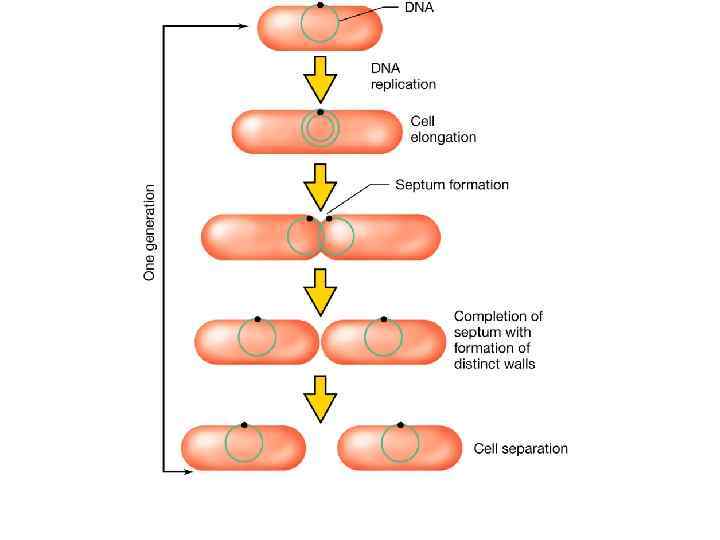
 Механизм распределения бактериальных хромосом А – клетка содержит частично реплицированную хромосому, прикрепленную к ЦМ в точке репликации; Б – репликация хромосомы завершена; В – разделение дочерних хромосом. 1 – ДНК; 2 – прикрепление хромосомы к ЦМ; 3 – ЦМ; 4 – клеточная стенка; 5 – синтезированный участок ЦМ; 6 – новый материал клеточной стенки.
Механизм распределения бактериальных хромосом А – клетка содержит частично реплицированную хромосому, прикрепленную к ЦМ в точке репликации; Б – репликация хромосомы завершена; В – разделение дочерних хромосом. 1 – ДНК; 2 – прикрепление хромосомы к ЦМ; 3 – ЦМ; 4 – клеточная стенка; 5 – синтезированный участок ЦМ; 6 – новый материал клеточной стенки.
 Конъюгация у бактерий Конъюгация – перенос генетического материала путем прямого контакта между клетками Клетки Escherichia coli, связанные между собой F-пилями. Фактор F – плазмида, содержит гены, необходимые для конъюгации.
Конъюгация у бактерий Конъюгация – перенос генетического материала путем прямого контакта между клетками Клетки Escherichia coli, связанные между собой F-пилями. Фактор F – плазмида, содержит гены, необходимые для конъюгации.
 Трансдукция Умеренный бактериофаг • Трансдукция – передача В+ участков бактериальной ДНК от клетки-донора клетке реципиенту при участии лизис бактериофагов. инкубация В- В капсиды вместо фаговой ДНК могут включаться фрагменты бактериальной ДНК – образуются дефектные фаги. В+
Трансдукция Умеренный бактериофаг • Трансдукция – передача В+ участков бактериальной ДНК от клетки-донора клетке реципиенту при участии лизис бактериофагов. инкубация В- В капсиды вместо фаговой ДНК могут включаться фрагменты бактериальной ДНК – образуются дефектные фаги. В+
 Трансформация • Трансформация – передача генов при помощи свободной растворимой ДНК, выделенной из клеток-доноров Трансформация протекает в три стадии: 1) адсорбция двуцепочечной ДНК на участках клеточной стенки компетентных клеток; 2) ферментативное расщепление связавшейся ДНК в некоторых случайно расположенных местах с образованием фрагментов 4 -5*106 Dа; 3) проникновение фрагментов ДНК с молекулярной массой не менее 5*106 Dа, сопровождающееся разрушением одной из цепей ДНК (последний этап энергозависим). Проникшая цепь ДНК рекомбинирует с генетическим материалом реципиентной клетки.
Трансформация • Трансформация – передача генов при помощи свободной растворимой ДНК, выделенной из клеток-доноров Трансформация протекает в три стадии: 1) адсорбция двуцепочечной ДНК на участках клеточной стенки компетентных клеток; 2) ферментативное расщепление связавшейся ДНК в некоторых случайно расположенных местах с образованием фрагментов 4 -5*106 Dа; 3) проникновение фрагментов ДНК с молекулярной массой не менее 5*106 Dа, сопровождающееся разрушением одной из цепей ДНК (последний этап энергозависим). Проникшая цепь ДНК рекомбинирует с генетическим материалом реципиентной клетки.
, трансформация с использованием отдельной молекулы ДНК (Б), трансдукция") Механизмы переноса бактериальной ДНК Конъюгация (А), трансформация с использованием отдельной молекулы ДНК (Б), трансдукция с помощью фагов (В).
Механизмы переноса бактериальной ДНК Конъюгация (А), трансформация с использованием отдельной молекулы ДНК (Б), трансдукция с помощью фагов (В).

 ПОЛУЧЕНИЕ ПРОДУЦЕНТОВ МЕТОДАМИ СЕЛЕКЦИИ Селекция – это направленный отбор мутантов – организмов, наследственность которых приобрела скачкообразное изменение в результате структурной модификации в нуклеотидной последовательности ДНК. Основные этапы селекции • Получение мутантов • Отбор мутантов: а) Отбор случайных (непредсказуемых) мутаций по количественному признаку; б) Отбор по количественному признаку среди мутантов с определенным фенотипом. Фенотип - совокупность всех признаков и свойств организма, сформировавшихся в процессе его индивидуального развития.
ПОЛУЧЕНИЕ ПРОДУЦЕНТОВ МЕТОДАМИ СЕЛЕКЦИИ Селекция – это направленный отбор мутантов – организмов, наследственность которых приобрела скачкообразное изменение в результате структурной модификации в нуклеотидной последовательности ДНК. Основные этапы селекции • Получение мутантов • Отбор мутантов: а) Отбор случайных (непредсказуемых) мутаций по количественному признаку; б) Отбор по количественному признаку среди мутантов с определенным фенотипом. Фенотип - совокупность всех признаков и свойств организма, сформировавшихся в процессе его индивидуального развития.
 Основные классы алкилирующих агентов Класс Представитель Структурная формула Иприты сернист ые Иприт S(CH 2 Cl)2 азотистые Азотистый иприт HN(CH 2 Cl )2 Эпоксиды Этиленоксид (CH 2)О Этиленими ны Этиленимин (CH 2)NH Алкилалкан Этилметансульф сульонат фонаты C 2 H 5 OSO 2 CH 3 -Лактоны -Пропиолактон Диазосоеди нения Диазометан CH 2 N ═ N
Основные классы алкилирующих агентов Класс Представитель Структурная формула Иприты сернист ые Иприт S(CH 2 Cl)2 азотистые Азотистый иприт HN(CH 2 Cl )2 Эпоксиды Этиленоксид (CH 2)О Этиленими ны Этиленимин (CH 2)NH Алкилалкан Этилметансульф сульонат фонаты C 2 H 5 OSO 2 CH 3 -Лактоны -Пропиолактон Диазосоеди нения Диазометан CH 2 N ═ N
 Селекция на устойчивость к структурному аналогу целевого продукта Основа метода: отбор клонов с нарушенной негативной регуляцией собственного биосинтеза E 1 E 2 E 3 A B Lys C Если в клетке в результате мутации ферменты, участвующие в биосинтезе, станут нечувствительны к негативной регуляции лизином, эта клетка может стать сверхпродуцентом S-(2 -аминоэтил)цистеин (AEC) вызывает аллостерическое ингибирование бета-аспараткиназы
Селекция на устойчивость к структурному аналогу целевого продукта Основа метода: отбор клонов с нарушенной негативной регуляцией собственного биосинтеза E 1 E 2 E 3 A B Lys C Если в клетке в результате мутации ферменты, участвующие в биосинтезе, станут нечувствительны к негативной регуляции лизином, эта клетка может стать сверхпродуцентом S-(2 -аминоэтил)цистеин (AEC) вызывает аллостерическое ингибирование бета-аспараткиназы
 Селективная среда На среде с AEC вырастают только клетки, в которых нарушена негативная регуляция лизином, и поэтому они не чувствительны и к AEC. Интактные клетки на этой среде не делятся, так как у них заблокирован биосинтез лизина AEC Использование селективной среды позволяет снизить количество исследуемых клонов до десятков
Селективная среда На среде с AEC вырастают только клетки, в которых нарушена негативная регуляция лизином, и поэтому они не чувствительны и к AEC. Интактные клетки на этой среде не делятся, так как у них заблокирован биосинтез лизина AEC Использование селективной среды позволяет снизить количество исследуемых клонов до десятков
 Ступенчатый отбор При конструировании сверхпродуцента Trp использовали следующие его структурные аналоги N селекции Дикий штамм Выход (г/л) 0. 15 1 2 4. 9 5. 7 3 4 5 7. 1 10. 0 12. 0
Ступенчатый отбор При конструировании сверхпродуцента Trp использовали следующие его структурные аналоги N селекции Дикий штамм Выход (г/л) 0. 15 1 2 4. 9 5. 7 3 4 5 7. 1 10. 0 12. 0
 Две возможности конструирования сверхпродуцентов Дикий штамм: Сверхпродуцент: Lys Увеличение") Отбор по биохимической недостаточности (ауксотрофности) Две возможности конструирования сверхпродуцентов Дикий штамм: Сверхпродуцент: Lys Увеличение интенсивности биосинтеза Lys Уменьшение интенсивности катаболизма Х АУКСОТРОФНЫЕ (БИОХИМИЧЕСКИЕ) МУТАНТЫ -мутанты бактерий с дополнительными питательными потребностями из-за потери способности синтезировать те или иные
Отбор по биохимической недостаточности (ауксотрофности) Две возможности конструирования сверхпродуцентов Дикий штамм: Сверхпродуцент: Lys Увеличение интенсивности биосинтеза Lys Уменьшение интенсивности катаболизма Х АУКСОТРОФНЫЕ (БИОХИМИЧЕСКИЕ) МУТАНТЫ -мутанты бактерий с дополнительными питательными потребностями из-за потери способности синтезировать те или иные
 Селекция в два этапа: 1. После мутагенеза клетки высевают") Отбор по биохимической недостаточности (ауксотрофности) Селекция в два этапа: 1. После мутагенеза клетки высевают на полной среде 2. Клетки всех клонов высевают на среде без вещества Х Те клоны, которые не размножаются на этой среде ауксотрофы по Х, следовательно могут продуцировать Lys в больших количествах Полная среда Среда без Х
Отбор по биохимической недостаточности (ауксотрофности) Селекция в два этапа: 1. После мутагенеза клетки высевают на полной среде 2. Клетки всех клонов высевают на среде без вещества Х Те клоны, которые не размножаются на этой среде ауксотрофы по Х, следовательно могут продуцировать Lys в больших количествах Полная среда Среда без Х
 Реверсия ауксотрофности: 1. Мутагенез 2. Отбор на селективной среде") Отбор по биохимической недостаточности (ауксотрофности) Реверсия ауксотрофности: 1. Мутагенез 2. Отбор на селективной среде (без Х) Выживают (размножаются) только те клетки, в которых в результате мутации вещество Х стало вновь синтезироваться
Отбор по биохимической недостаточности (ауксотрофности) Реверсия ауксотрофности: 1. Мутагенез 2. Отбор на селективной среде (без Х) Выживают (размножаются) только те клетки, в которых в результате мутации вещество Х стало вновь синтезироваться
 нельзя скрещивать неродственные") ПОЛУЧЕНИЕ ПРОДУЦЕНТОВ МЕТОДАМИ ГЕНЕТИЧЕСКОЙ ИНЖЕНЕРИИ Ограничения использования метода селекции: • 1) нельзя скрещивать неродственные виды; • 2) нельзя извне управлять процессом рекомбинации в организме; • 3) нельзя предугадать, какое получится потомство.
ПОЛУЧЕНИЕ ПРОДУЦЕНТОВ МЕТОДАМИ ГЕНЕТИЧЕСКОЙ ИНЖЕНЕРИИ Ограничения использования метода селекции: • 1) нельзя скрещивать неродственные виды; • 2) нельзя извне управлять процессом рекомбинации в организме; • 3) нельзя предугадать, какое получится потомство.
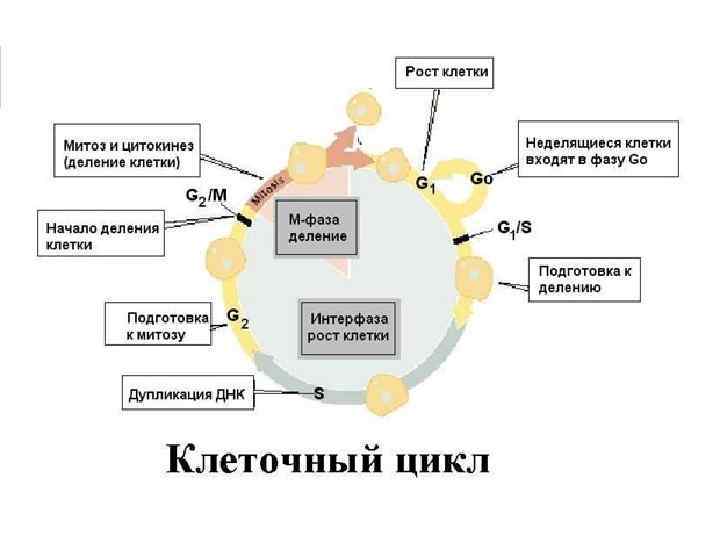
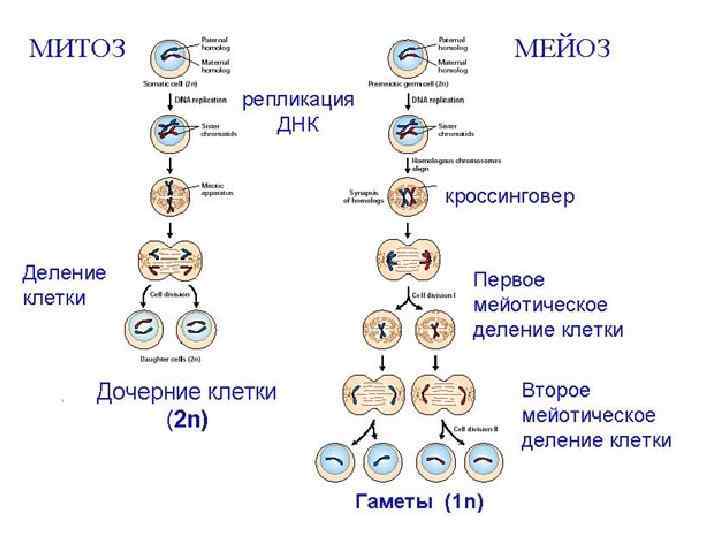
 Теоретические предпосылки генной инженерии 1. Молекулярные механизмы матричного синтеза: ДНК Репликация ДНК РНК Транскрипция м. РНК белок Трансляция Обмен генами у гомологичных хромосом при половом процессе Рекомбинация 2. Кольцевые двуспиральные малые молекулы ДНК, автономно размножающиеся в бактериальной клетке и несущие маркерный ген Плазмиды 3. Ферменты, способные расщеплять ДНК в строго определенном месте с образованием липких концов у образуемых фрагментов Рестриктазы
Теоретические предпосылки генной инженерии 1. Молекулярные механизмы матричного синтеза: ДНК Репликация ДНК РНК Транскрипция м. РНК белок Трансляция Обмен генами у гомологичных хромосом при половом процессе Рекомбинация 2. Кольцевые двуспиральные малые молекулы ДНК, автономно размножающиеся в бактериальной клетке и несущие маркерный ген Плазмиды 3. Ферменты, способные расщеплять ДНК в строго определенном месте с образованием липких концов у образуемых фрагментов Рестриктазы
 Возможности генной инженерии 1. Можно скрещивать индивидуальные гены видов, стоящих на разных ступенях эволюции. В основе рекомбинации гетерологичных ДНК in vitro лежит прием, позволяющий с помощью рестриктаз подготовить молекулы для скрещивания, то есть разрезать разные ДНК с образованием одинаковых липких концов. 2. Можно управлять процессом рекомбинации, так как он происходит в пробирке и не защищен запрещающими механизмами организма. 3. Можно предсказать результат, т. к. отбирается потомство одной молекулы ДНК (молекулярное клонирование).
Возможности генной инженерии 1. Можно скрещивать индивидуальные гены видов, стоящих на разных ступенях эволюции. В основе рекомбинации гетерологичных ДНК in vitro лежит прием, позволяющий с помощью рестриктаз подготовить молекулы для скрещивания, то есть разрезать разные ДНК с образованием одинаковых липких концов. 2. Можно управлять процессом рекомбинации, так как он происходит в пробирке и не защищен запрещающими механизмами организма. 3. Можно предсказать результат, т. к. отбирается потомство одной молекулы ДНК (молекулярное клонирование).
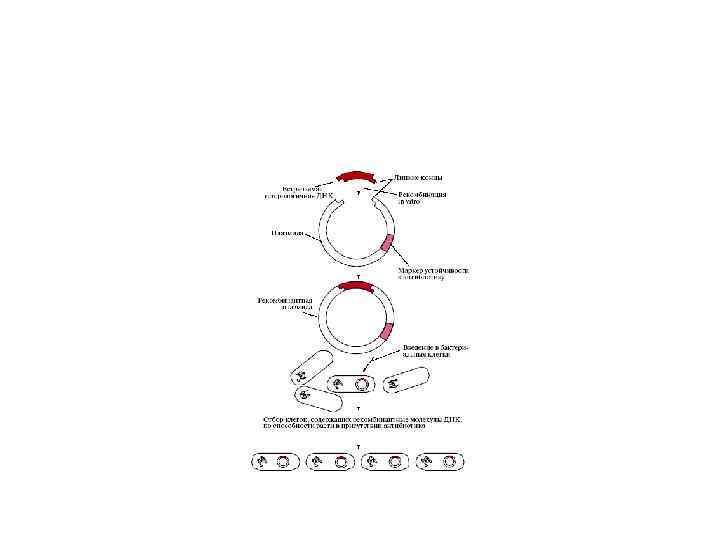
Получение чужеродной ДНК; 2) Рекомбинация in vitro ДНК-вектора и") Клонирование фрагмента ДНК в плазмиде 1)Получение чужеродной ДНК; 2) Рекомбинация in vitro ДНК-вектора и ДНКгена; 3) Введение рекомбинантной плазмиды в клетку; 4) Молекулярное клонирование.
Клонирование фрагмента ДНК в плазмиде 1)Получение чужеродной ДНК; 2) Рекомбинация in vitro ДНК-вектора и ДНКгена; 3) Введение рекомбинантной плазмиды в клетку; 4) Молекулярное клонирование.
 Получение к. ДНК для клонирования
Получение к. ДНК для клонирования