репликация регуляция(Пестряков).pptx
- Количество слайдов: 49



Разин Сергей Владимирович - Кафедра молекулярной биологии Биофак МГУ

Bidrectional DNA synthesis Replication forks will merge
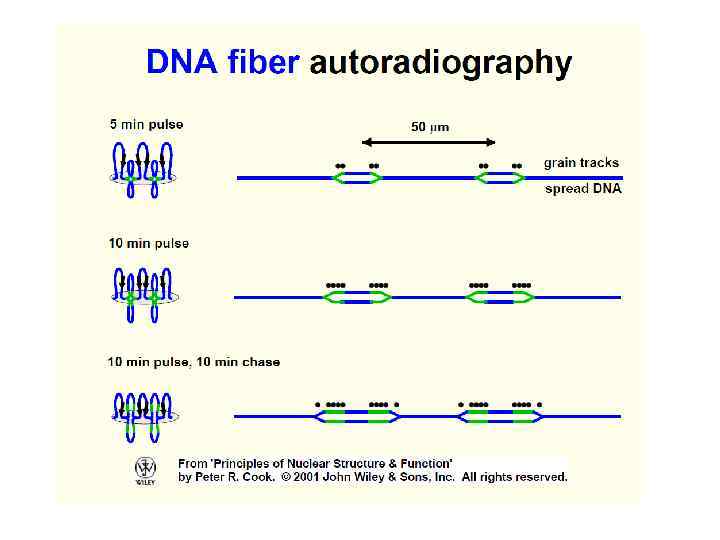

Репликоны впервые были обнаружены при изучении реплицирующейся ДНК высших эукариот с использованием метода “fiber autoradiography”

20 мин пульс 40 т. п. н. 1, 4 мкм =

ARS элементы Saccharomyces cerevisiae Cloning site Selective marker cлучайные фрагменты геномной ДНК стабильный рост если плазмида реплицируется Selective marker трансформация дрожжевых клеток и высев на селективную среду нестабильный рост если плазмида не реплицируется

Функциональные домены дрожжевых ARS элементов ORC : A six subunit protein complex which has been implicated as being the eukaryotic DNA replication initiator protein. Subunits are named according to their size, with ORC 1 being the largest and ORC 6 being the smallest subunit. Yeast ORC specifically binds to replication origins in an ATP dependent manner and has been shown to possess ATPase activity.

ARS консенсус ORC, Origin Recognition Complex состоит из 6 белков с молекулярными весами от 50 до 120 КД ORC белки чрезвычайно консервативны Они присутствуют в клетках как низших, так и высших эукариот

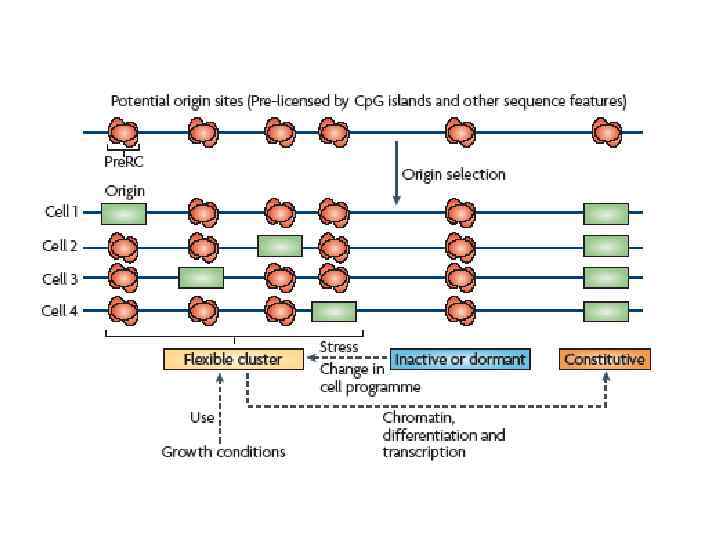
Все реально работающие участки начала репликации в хромосомах Saccharomyces cerevisiae колокализуются c ARS элементами Существует много ARS элементов, которые не работают в качестве участков начала репликации в дрожжевых хромосомах

У высших эукариот поиски автономно реплицирующихся последовательностей привели к очень невыразительным и противоречивым результатам С весьма низкой эффективностью в эукариотических клетках реплицируется любая плазмидная ДНК. Включение в эту ДНК эукариотических инсерций повышает эффективнось репликации вне зависимости от последовательности ДНК. В то же время существует позитивная корреляция между длиной инсерции и эффективностью автономной репликации
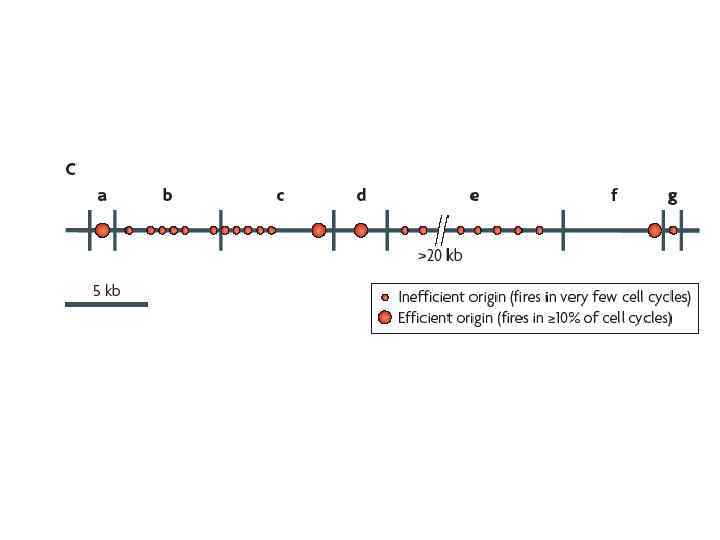

Кластерный анализ позволил идентифицировать 41 зону инициации репликации, между которыми распределялись 307 индивидуальных участков начала репликации в пределах изучаемой области протяженностью ~ 1, 5 MB 100 KB зоны инициации репликации индивидуальные инициации репликации (белые вертикальные линии)

Репликационные вилки, двигающиеся от активного ori супрессируют другие ori, находящиеся в пределах зоны 55+/-38 KB в обе стороны от активного ori. Внутри области инициации наблюдается полная суппрессия всех потенциальных ori На картинке суппрессированные области инициации показаны серыми горизонтальными линиями

При снижении скорости репликации происходит активация «спящих» ori

В клетках высших эукариот инициация репликации в принципе может происходить где угодно. В эмбриональных клетках (до стадии средней бластулы) и в экстрактах из клеток ксенопуса инициация происходит через короткие интервалы (на молекуле ДНК) и без всякой специфичности в отношении последовательностей В неэмбриональных клетках сушествуют предпочтительные места инициации которые могут быть достаточно короткими (репликатор домена бета-глобиновых генов) и достаточно протяженными (зона инициации репликации локуса DHFR) Короткие репликаторы характеризуются наличием ряда необходимых элементов (АТ-богатые последовательности, полупуриновые/полипиримидиновые блоки, изогнутые последовательности ДНК). Набор и сочетание этих элементов могут быть разными в разных репликаторах. Вопрос о том, нужна ли транскрипция близлежащих генов для активации репликатора остается пока неясным

Cell Cycle Checkpoints… Gap 2 Gap 1
 – синтез")
G 1 b фаза - транскрипция, трансляция необходимых макромолекул S фаза (синтез) – синтез ДНК, удвоение хромосом G 2 фаза – конденсация хроматина, подготовка к митозу M фаза (митоз) – разделение хромосом, цитокинез, деление клетки (практически полное отсутствие матричного биосинтеза)
")
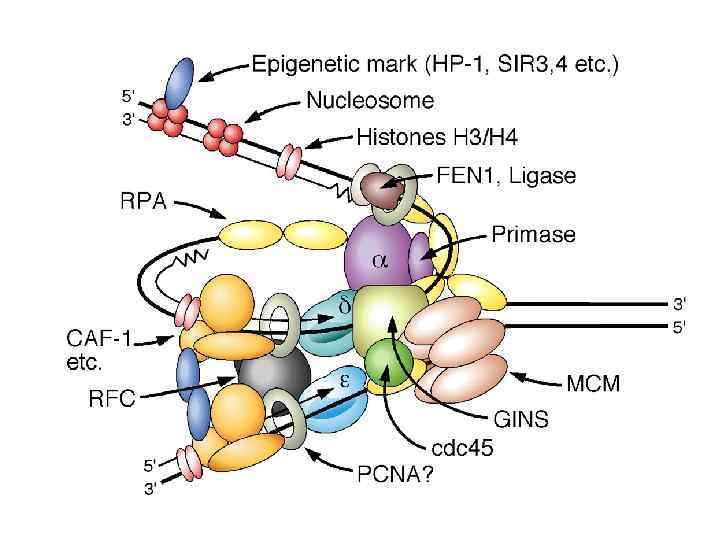
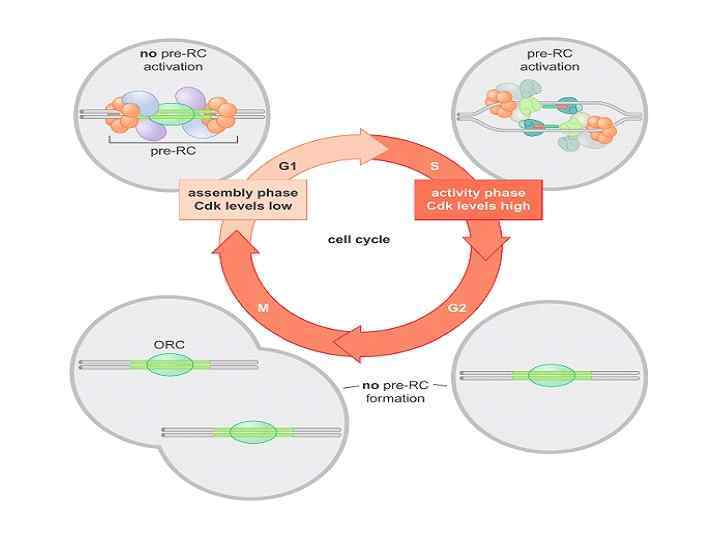
В поздней М-ранней G 1 фазах Cdc 6 отвечает за загрузку неактивной геликазы MCM(27) в пререпликативные комплексы. При этом загрузка АТФ-зависима и возможна только в присутствии Ctd 1, формирующего промежуточные комплексы с MCM(2 -7) Лицензированный ориджин – участок ДНК на котором собран комплекс ORC-cdc 6 -cdt 1 -MCM(2 -7)

Процессивные хеликазы, участвующие в репликации ДНК SV-40 T антиген (3’-5’ прцессивная хеликаза, денатурирующая ДНК со скоростью 75 – 100 п. н. /мин. Использует энергию АТФ) Низшие и высшие эукариоты – MCM 2 -7 комплекс (3’-5’ процессивная хеликаза. Активируется фосфорилированием. Испоьзует энергию АТФ Каталитический центр Участок связывания АТФ

Активация pre. RC комплекса происходит путем фосфорилирования компонентов циклин-зависимыми киназами в G 1/S контрольной точке (checkpoint)
 вызывает их активацию и начальное расплетание ДНК Фосфорилирование pre. RC")
Фосфор-е геликаз MCM(2 -7) вызывает их активацию и начальное расплетание ДНК Фосфорилирование pre. RC инициирует последовательную загрузку факторов Cdc 45 и GINS (факторы отвечают за загрузку пол-альфа-праймазного комплекса и связь геликазы и реплисомы, соответственно) Сформированные одноцепочечные участки ДНК поддерживаются в таком состоянии фактором RPA

После загрузки пол-альфа-праймазный комплекс формирует РНК-ДНК-праймеры затравки репликации

После синтеза 30 -мерных РНК-ДНК-праймеров с праймированной структурой связывается фактор RFC, который в свою очередь в АТФ-зависимой манере загружает на праймер-матричный дуплекс “скользящий зажим” (фактор процессивности репликативных ДНК полимераз) PCNA. Следующим этапом происходит ассоциация репликативных полимераз дельта и эпсилон и формирование репликативных вилок, формирование реплисомы закончено.

Репликация ДНК у животных После того как синтез ДНК закончен места инициации вновь заполняются комплексом ORC (связывание АТФ-зависимое) на протяжении частично G 2 и М-G 1 фаз, т. о. обеспечивается выбор ориджинов для будущего раунда репликации. Повторная сборка pre-RC комплексов в этих местах в рамках текущего цикла невозможна. Почему? На этих стадиях клеточного цикла не хватает факторов cdc 6, ctd 1 (фосфорилирование приводит к активации деградации этих белков, кроме того cdt 1 выводится из оборота через связывание с белком Геминином). На ориджин невозможна загрузка геликазы MCM 2 -7, ориджин “не лицензирован”. Второй путь запрета повторной сборки репликативных комплексов – убиквитинилирование отдельной субъединицы Orc 1, при которой невозможно формирование продуктивного комплекса ORC в ориджине, а также ее полиубиквитинилирование, ведущее к протеолитической деградации, на указанных стадиях клеточного цикла.

ORC цикл, исключающий возможность повторной инициации до прохождения митоза

По ходу осуществления репликации сестринские хроматиды скрепляются когезиновыми сшивками
. Секурин")
В анафазе митоза клейзин расщепляется сепаразой, которая становится активной после удаления ингибитора (секурина). Секурин убиквитинилируется при посредстве APC (Anaphase Promoting Complex)

Индивидуальные гены реплицируются в разное время по ходу S фазы. Предложено несколько методических подходов, позволяющих определить предпочтительное время репликации гена (replication timing) Определение относительной представленности пробы в препаратах реплицирующейся ДНК, изолированных на различных стадиях S фазы Подсчет относительного количества клеток (в асинхронной культуре), содержащих две копии представляющего интерес фрагмента ДНК
 молчащие гены обычно являются")
активные гены обычно являются раннереплицирующимися (реплицируются в начале S фазы) молчащие гены обычно являются позднереплицирующимися (реплицируются в конце S фазы)

В ядре эукариотической клетки репликоны организованы в кластеры После включения в новосинтезированную ДНК бромодезоксиуридина эти кластеры можно увидеть с использованием иммунофлуоресцентного окрашивания. Распределение активных центров репликации изменяется по ходу S-фазы

Разные типы фокусов репликации можно увидеть и анализируя распределение PCNA Мышиние клетки 3 Т 6 в геном которых интегрирован ген, кодирующий GFP-PCNA Early S Mid S Late S

В начале S-фазы в клетках присутствует несколько сотен фокусов репликации

Что движется и что остается неподвижным ? DNA pol

cdc-гены, сеll division control • Реакции обратимого фосфорования • Киназы, активируемые циклинами • Циклины • Фосфатазы • протеолиз • Убиквитинилирование (poly. Ub и mono. Ub) – протеолиз и изменение белок-белковых изаимодействий (соответственно)

Cell Cycle Checkpoints… Gap 2 Gap 1
 – решается вопрос о дальнейшем продвижении по")
Начало клеточного цикла – restriction point (R) – решается вопрос о дальнейшем продвижении по G 1 фазе (достаточно питания, есть внешний сигнал - факторы роста) G 1 b фаза - транскрипция, трансляция необходимых макромолекул - окончательное лицензирование ориджинов репликации G 1/S checkpoint – решается вопрос о готовности к синтезу ДНК - индукция инициации синтеза ДНК S фаза – синтез ДНК, удвоение хромосом DNA damage/replication stall checkpoint G 2 фаза – конденсация хроматина, подготовка к митозу G 2/M checkpoint – решается вопрос о готовности к делению (полностью реплицированная ДНК, отсутствие повреждений ДНК) M фаза (митоз) – разделение хромосом, цитокинез, деление клетки (практически полное отсутствие матричного биосинтеза) M checkpoint – проверяется крепление хромосом на веретене деления

Ключевые ферменты – циклин-зависимые киназы: 2 субъединицы – CDK – каталитическая субъединица циклин – активирующая с-а Концентрация циклинов по стадиям клеточного цикла

Ключевые переключения под действием циклин-зависимых киназ: Фосфорилирование целевых белков “включает или выключает” процесс R (restriction point) и далее G 1 b – Cyclin D/CDK 4, Cyclin D/CDK 6 G 1/S checkpoint – Cyclin E/CDK 2 S фаза (включая внутренний checkpoint) – Cyclin A/CDK 2 G 2 и M фазы – Cyclin. B 3/CDK 1, Cyclin. B 3/CDK 2 G 2/M checkpoint – Cyclin A/CDK 1 М фаза – Cyclin B/CDK 1

контроль начала S фазы Cdk 2 - циклин E Сdk 4 Cdk 6 циклин D Cdk 4 активирует работу транскрипционного фактора E 2 F-1, включающего гены, работа которых необходима для репликации ДНК. Cdk 4 инактивирует белок Rb, который является ингибитором E 2 F-1 Cdk 2, 4. 6 являются сенсорами различных сигналов. Частью сенсорной системы являются ингибиторы Cdk (CKI).
 1 2 (G 1/S чекпоинт и далее до G 1")
(до G 1/S чекпоинта) 1 2 (G 1/S чекпоинт и далее до G 1 a) Фосфорилирование компонентов ORC 1. Препятствует формированию новых pre. RC 2. Активирует-запускает ориджин

контроль М фазы Перенос цитоплазмы из митотической клетки в интерфазную стимулирует вхождение интерфазной клетки в митоз MPF - Mitotic Promoting Factor MPF = Cdk 1 + Cyclin B Активная Cdk 1 фосфорилирует комплекс мишеней, участвующих в начале митоза (белки хромосом, ядерной оболочки, ядрышка, центросом и т. д. )

Cdk 1, как и другие киназы, регулирующие клеточный цикл, экспрессируется постоянно, тогда как уровень экспрессии циклинов изменяется в зависимости от фазы клеточного цикла

Активность самой Cdk 1 регулируется на двух уровнях: 1. Присоединение циклина B = стимуляция активности 2. Фосфорилирование треонина 14 и тирозина 15 киназой Wee 1 = инактивация (стерически препятствует связыванию АТФ с активным центром) 2 а. Дефосфорилирование - процесс обратный фосфорилированию, катализируется фосфатазами. Инактивация Cdk 1 после фосфорилирования Wee 1 обратима. Восстановление активности Cdk 1 (а следовательно и прохождение М-фазы) происходит под действием фосфатазы cdc 25 G 2/M дефосфорилирование фосфатазой Cdc 25 M/G 1 циклин В убиквитинилируется белками APC и подвергается протеасомной деградации

Координация репарации ДНК и прохождения клеточного цикла в G 2/M checkpoint-е

Checkpoint G 2/M - на исправление повреждений ДНК cdc 2 = CDK 1 – киназа, отвечающая за продвижение в М-фазе клеточного цикла (составная часть MPC) Rad 3 = ATM/ATR – киназы, активирующиеся в ответ на повреждения ДНК Wee 1 – киназа, инактивирующая CDK 1 Cdc 25 – фосфатаза, снимающая ингибирование CDK 1 1. 2. 3. 4. Повреждения ДНК активируют киназы rad 3 (ATM/ATR) Киназы rad 3 (ATR/ATM) фосфорилируют киназу chk 1, значительно стимулируя ее активность Киназа chk 1 фосфорилирует киназу wee 1, увеличивая ее стабильность, а также фосфорилирует фосфатазу cdc 25, ингибируя ее и препятствуя попаданию cdc 25 в ядро Происходит сильный сдвиг в сторону фосфорилирования CDK 1 (cdc 2) в отсутствии дефосфорилирования. Киназа CDK 1 и соответственно фактор MPF инактивированы – клетка тормозится в фазе G 2 до тех пор пока повреждения ДНК не будут исправлены.
 CKI – ингибиторы циклин-зависимых киназ транскрипционный фактор, активирующий работу генов,")
стадия R (restriction point) CKI – ингибиторы циклин-зависимых киназ транскрипционный фактор, активирующий работу генов, участвующих в репликации ДНК
репликация регуляция(Пестряков).pptx