


ПЗ № 2. • Возбуждение в ЦНС. • Торможение и координационная деятельность ЦНС. • Рефлекс и функциональная система.

Формируемые компетенции: • Актуальность темы: данная тема вносит вклад в формирование следующих компетенций: ОК-1, ПК-5, ПК-9, ПК-16, ПК-27, ПК-31. • Знание изучаемой темы необходимо для понимания процессов регуляции функций организма человека.
. • Изучить")
Цели занятия: • Учебная: • Изучить свойства нервных центров (ПК-5, ПК 16). • Изучить закономерности возбуждения в ЦНС (ПК-5, ПК-16). • Изучить закономерности торможения в ЦНС (ПК-5, ПК-16). • Изучить принципы координационной деятельности ЦНС (ПК-5, ПК-16). • Научиться применять терминологию по изучаемой теме (ПК-5, ПК-16, ПК-27, ПК-31).

Цели занятия: • Развивающая: • Формирование системного подхода к пониманию регуляции функций организма человека (ОК-1, ПК-5, ПК-9, ПК-16, ПК-27). • Воспитательная: • Формирование способности и готовности реализовать этические и деонтологические аспекты врачебной деятельности в общении с коллегами (ПК-1).

Межпредметные связи Фармакология Неврология Патологическая физиология Нормальная физиология Биохимия Гистология, цитология, эмбриология Возбуждение в ЦНС. Торможение и координационная деятельность ЦНС. Рефлекс и функциональная система Клиническая патофизиология Анатомия Физика. Математика

Внутрипредметные связи ПЗ№ 1. Физиология нервов и синапсов ПЗ№ 8. Методы исследования функций ЦНС ПЗ№ 7. Нервная регуляция висцеральных функций. Физиология автономной (вегетативной) нервной системы ПЗ№ 3. Частная физиология ЦНС. Спинной мозг Возбуждение в ЦНС. Торможение и координационная деятельность ЦНС. Рефлекс и функциональная система ПЗ№ 6. Кора больших полушарий ПЗ№ 4. Частная физиология ЦНС. Продолговатый мозг, мост, мозжечок ПЗ№ 5. Частная физиология ЦНС. Ретикулярная формация. Промежуточный, передний мозг. Лимбическая система

1. Установите соответствие Принцип координационной деятельности ЦНС 1. А Облегчение Б Окклюзия 2. заключается В ослаблении эффекта одновременного действия двух сильных раздражителей по сравнению с суммой их раздельных эффектов. В превышении эффекта одновременного действия двух слабых раздражителей над суммой их раздельных эффектов.

2. Установите соответствие Принцип координационной заключается деятельности ЦНС 1. В участии в разных А Общий конечный путь рефлекторных реакциях одних и тех же эфферентных нейронов и эффекторов. 2. В наличии в мозге центра, Б Принцип доминанты обладающего повышенной возбудимостью, инертностью и способностью тормозить и суммировать возбуждения других центров.

3. Установите соответствие Свойство нервного проявляется центра А Посттетаническая 1. В способности изменять потенциация свою функцию, расширять функциональные возможности. 2. В усилении рефлекторной Б Низкая аккомодационная реакции после длительного способность ритмического раздражения нервного центра. 3. В способности реагировать на медленно нарастающие по силе раздражители.

4. Установите соответствие Вид торможения А Латеральное Б Возвратное В Реципрокное выполняет функцию 1. Подавляет возбуждение центра антагонистической функции. 2. Устраняет диффузную иррадиацию возбуждения. 3. Прекращает выход медиатора в синаптическую щель. 4. Ослабляет возбуждение мотонейронов их собственными импульсами через клетки Реншоу.

Нейронные цепи в ЦНС

Нервные сети • Локальные сети – содержат нейроны с короткими аксонами, осуществляющие взаимосвязи в пределах одного уровня. Обеспечивают задержку информации в пределах этого уровня. • Пример: кольцевые нейронные цепочки Лоренто де Но, возбуждение в которых циркулирует по замкнутому кругу. Возврат возбуждения к «первому» нейрону кольцевой цепи получил название реверберации возбуждения. • Локальные сети обеспечивают надежность нервной регуляции за счет дублирования элементов, так как многие нейроны локальных сетей имеют одинаковые синаптические связи и функционируют попеременно, то есть являются взаимозаменяемыми.

РЕВЕРБЕРАЦИЯ ВОЗБУЖДЕНИЯ В НЕРВНОЙ СЕТИ по Лоренто-де-Но

Нервные сети • Иерархические сети - обеспечивают постепенное включение нейронных структур более высокого уровня. • Если в результате афферентная информация поступает к возрастающему числу нейронов, этот принцип называется дивергенция. • Если от многих нейронов возбуждение сходится к меньшему числу нейронов, этот принцип называется конвергенция.

Нервные сети • Дивергентные сети с одним входом – представляют собой нейронные ансамбли, в которых один нейрон образует выходные связи с большим количеством других клеток разных иерархических уровней и разных нервных центров. • Эти сети не являются специфическими для реализации определенных рефлексов, а обеспечивают интеграцию разных рефлекторных актов и общее состояние активности нейронов разных отделов мозга.

Свойства нервных центров • Нервный центр: • совокупность функционально связанных нейронов, расположенных в одной или нескольких структурах ЦНС и обеспечивающих осуществление регуляции определенных функций организма; • совокупность нейронов, обеспечивающих реализацию определенного рефлекса • Свойства нервных центров определяются: • свойствами образующих их нейронов; • особенностями структурно-функциональных связей нейронов; • свойствами центральных синапсов.

Свойства нервных центров • Пространственная и временная суммация – основана на свойстве каждого нейрона в центре к суммации как возбуждения, так и торможения. • Временная суммация – при увеличении частоты слабых афферентных сигналов амплитуда ВПСП нарастает до критического уровня из-за повышения эффективности синаптического проведения, что вызывает возбуждение нейронов и рефлекторный ответ.

Свойства нервных центров • Пространственная суммация – • слабые раздражения нескольких участков рецептивного поля, • в отдельности не способные реализовать рефлекс, • вызывают в нейронах центра несколько ВПСП, • которые суммируются, приводя к формированию на мембране нейрона ПД, • распространяющихся по эфферентным волокнам, вызывая рефлекторную реакцию.

Суммация в нервных центрах

Временная суммация

Пространственная суммация

Постсинаптические потенциалы ТПСП Мембранный потенциал Время ВПСП

Возбуждающий постсинаптический потенциал • Волна деполяризации, которая приближает МП к КУД, облегчая возможность генерации ПД

Временная суммация? Стимулы и ВПСП слишком далеко друг от друга по времени. Суммации нет Время

Временная суммация! Стимулы и ВПСП близко по времени, следующий ВПСП на одном синапсе суммируется с предыдущим, деполяризация нарастает и достигает КУД: генерируется ПД КУД Время

Пространственная суммация Близкие во времени ВПСП от двух синапсов суммируются, деполяризация нарастает и генерируется ПД THRESHOLD TIME

Электрофизиологическая картина суммации • Каждый последующий ВПСП начинается раньше, чем завершится предыдущий, в итоге ВПСП суммируются

Пространственная суммация

Свойства нервных центров • Трансформация ритма возбуждения – увеличение или уменьшение частоты нервных импульсов в эфферентных проводниках (на выходе) по сравнению с частотой афферентной импульсации (на входе центра), что связано с механизмом синаптической передачи (трансформация ритма как свойство синапса) и интегративной деятельностью нейронов. • В синапсе происходит трансформация ритма возбуждения – частота импульсов в нерве меньше или больше частоты возбуждения постсинаптической мембраны нейрона. • Лабильность синапса существенно меньше, чем нерва.

Трансформация ритма 50 А А ? 50 В Фазовые соотношения входящих импульсов 50 В А В 10 0 1 (следующие попадают в рефрактерность предыдущего

Трансформация ритма Триггерные свойства аксонного холмика Порог 30 мв Порог 10 мв Тело нейрона Ек Ео Аксонный холмик Ек Ео «На ружейный выстрел нейрон отвечает пулеметной очередью»
")
Свойства нервных центров • Посттетаническая потенциация – увеличение амплитуды ВПСП после серии частых (тетанизирующих) ритмических возбуждений, что связано с временной суммацией частых ВПСП и активацией синаптического проведения из-за увеличения числа квантов медиатора. • Длительность состояния потенциации синапсов может достигать нескольких часов, что играет роль в процессах обучения и памяти.

Свойства нервных центров • Тонус нервного центра – состояние некоторого уровня активности нейронов, обеспечивающей их готовность к рефлекторной деятельности и проявляющейся постоянной эфферентной импульсации низкой частоты к органам-эффекторам. • Обусловлен небольшим уровнем афферентных сигналов от различных рецептивных полей, действием на нейроны метаболитов и других гуморальных раздражителей из клеточной микросреды. • Проявления: спонтанная электрическая активность нейронов, некоторая активность эффекторов (тонус скелетной мускулатуры, гладких мышц сосудов и т. п. )
 электрическая активность – периодическое генерирование импульсов (потенциалов действия)")
Свойства нервных центров • Спонтанная (фоновая) электрическая активность – периодическое генерирование импульсов (потенциалов действия) нервными клетками центра в состоянии покоя, т. е. без специфического раздражения рецептивного поля рефлекса. • Обусловлена тем, что организму не свойственно абсолютное отсутствие раздражителей или информационный покой, при этом за счет дивергенции и конвергенции возбуждений в нервных сетях нейроны центра всегда получают возбуждающие импульсы от клеток других нервных центров. • В происхождении спонтанной активности нейронов играют роль метаболические сдвиги внутриклеточной среды и микроокружения клеток, суммация миниатюрных потенциалов постсинаптической мембраны, формирующиеся из-за утечки единичных квантов медиатора.

Экранный принцип функционирования корковых полей • заключается в том, что • рецептор проецирует свой сигнал не на один нейрон коры, а на их поле, которое образуется коллатералями и связями нейронов. • В результате сигнал фокусируется не точка в точку, а на множестве нейронов, что обеспечивает его полный анализ и возможность передачи в другие заинтересованные в процессе структуры. • Экранный принцип реализуется благодаря особой организации взаимодействия входных и выходных элементов коры.
 импульсы поступают в кору снизу, поднимаются")
Экранный принцип функционирования корковых полей • Входные (афферентные) импульсы поступают в кору снизу, поднимаются к звездчатым и пирамидным клеткам 3 -4 -5 слоев коры. • От звездчатых клеток 4 -го слоя сигнал идет к пирамидным нейронам 3 -го слоя, а отсюда - по ассоциативным волокнам - к другим полям, областям коры мозга. • Звездчатые клетки 3 поля переключают сигналы, идущие в кору, на пирамидные нейроны 5 слоя, • отсюда обработанный сигнал уходит из коры к другим структурам мозга.

Экранный принцип функционирования корковых полей • В коре входные и выходные элементы вместе со звездчатыми клетками образуют так называемые «колонки» - функциональные единицы коры, организованные в вертикальном направлении. • Доказательством этому является то, что если микроэлектрод погружается перпендикулярно в кору, то на своем пути он встречает нейроны, реагирующие на один вид раздражения, если же микроэлектрод идет горизонтально по коре, то он встречает нейроны, реагирующие на разные виды стимулов.

Экранный принцип функционирования корковых полей • Колонка имеет диаметр около 500 мкм и определяется зоной распределения коллатералей восходящего афферентного таламокортикального волокна. • Соседние колонки имеют взаимосвязи, объединяющие множество колонок в организации той или иной реакции. • Возбуждение одной из колонок приводит к торможению соседних. • Каждая колонка может иметь ряд ансамблей, реализующих какую-либо функцию по вероятностатистическому принципу. • Этот принцип заключается в том, что в реакции при повторном раздражении участвует не вся группа нейронов, а только ее часть, причем, в каждом случае эта часть участвующих нейронов может быть разная.

Свойства нервных центров • Последействие и пролонгирование возбуждения – связаны с длительными следовыми потенциалами в нейронах, улучшением синаптического проведения, наличием кольцевых нейронных цепей и реверберацией возбуждения. • Эти процессы играют роль в процессах обучения и памяти.

Дивергенция нервных импульсов

Принцип конвергенции

←Одновременная индукция Индукция - возникновение противоположного по знаку нервного процесса вслед за существующим процессом (последовательная И. ) или за его территориальными пределами (одновременная И. ). Описана английским физиологом Ч. Шеррингтоном (1906). И. называется положительной, если первичный процесс - торможение, вслед за которым, по законам И. , возникает возбуждение, и отрицательной, если соотношение обратное.

Индукция нервных процессов • возникновение или усиление противоположного нервного процесса вокруг очага возбуждения или торможения - пространственная или одновременная И • возникновение противоположного нервного процесса в том же самом пункте коры головного мозга последовательная И • возникновение процесса возбуждения, вызванное торможением, называется положительной И • возникновение торможения вслед возбуждению – отрицательной И. • В коре ГМ после возбуждения всегда возникает торможение активности, и наоборот, после торможения - возбуждение (последовательная индукция).
 индукции в клетках, смежных с теми, где только что вызывалось")
Индукция • При (+) индукции в клетках, смежных с теми, где только что вызывалось торможение, после прекращения действия тормозного сигнала возникает состояние повышенной возбудимости. • Вследствие этого импульсы, поступающие к нейронам при действии положительного раздражителя, вызывают повышенный эффект. • При (–) индукции в клетках коры, окружающих возбужденные нейроны, возникает процесс торможения.
 индукция ограничивает иррадиацию процесса возбуждения в коре мозга. • (–) индукцией")
Индукция • (–) индукция ограничивает иррадиацию процесса возбуждения в коре мозга. • (–) индукцией можно объяснить торможение условных рефлексов более сильными посторонними раздражениями (внешнее безусловное торможение). • Такое сильное раздражение вызывает в коре мозга интенсивное возбуждение нейронов, вокруг которых появляется широкая зона торможения нейронов, захватывающая клетки, возбужденные условным раздражителем. • Явления (–) и (+) индукции в коре ГМ подвижны, постоянно сменяют друга. В разных пунктах коры мозга одновременно могут возникать очаги возбуждения и торможения, (+) и (–) индукции.

Иррадиация • Когда торможение не в состоянии сдерживать возбудительный процесс в определенной зоне, возникает иррадиация возбуждения в коре. • Иррадиация может происходить по коре от нейрона к нейрону, по системам ассоциативных волокон 1 -го слоя, тогда она имеет очень малую скорость - 0, 5 -2, 0 м в секунду. • Иррадиация возбуждения возможна и за счет аксонных связей пирамидных клеток 3 -го слоя коры между соседними структурами, в том числе, между разными анализаторами. • Иррадиация возбуждения обеспечивает взаимоотношение состояний областей коры при организации условнорефлекторного и других форм поведения.
 - распространение возбуждения (торможения) по")
Принципы распространения процессов возбуждения в ЦНС Иррадиация возбуждения (торможения) - распространение возбуждения (торможения) по нервной системе. Условия, влияющие на иррадиацию возбуждения. 1. Сила раздражителя. 2. Возбудимость ЦНС. 3. Функции тормозных нейронов.

Иррадиация процессов возбуждения и торможения Схема иррадиации возбуждения Схема иррадиации торможения
. • При достаточной силе нервных")
• Иррадиация проявляется при слабой интенсивности возбуждения (торможения). • При достаточной силе нервных процессов они концентрируются. • При чрезмерной силе нервных процессов они вновь начинают иррадиировать. • Дальность распространения нервных процессов от исходного пункта зависит от их силы: • чем интенсивнее нервный процесс в исходном пункте, тем дальше он распространяется и тем сильнее его воздействие на соседние участки мозга.

Закономерности функционирования химических синапсов • Односторонний характер проведения возбуждения. • Наличие химических передатчиков – медиаторов. • Свойства синапсов определяются природой медиатора и постсинаптических рецепторов. • Наличие хемочувствительных рецепторуправляемых каналов в постсинаптической мембране. • Квантовый характер высвобождения медиатора.

Закономерности функционирования синапсов • Количество квантов медиатора пропорционально частоте приходящих к синапсу нервных импульсов. • Эффективность синаптической передачи возрастает при частом использовании синапсов (эффект тренировки из-за ускорения синтеза медиатора, увеличения концентрации ионизированного кальция в пресинаптической терминали) и падает при бездействии (т. н. функциональная пластичность синапсов.

Закономерности функционирования синапсов • Длительная чрезмерная частота импульсов ослабляет или прекращает синаптическую передачу (эффект истощения). • Синаптическая передача не подчиняется закону «все или ничего» . • Скорость проведения информации в синапсе значительно меньше, чем скорость распространения возбуждения в нервном волокне (синаптическая задержка).

Закономерности функционирования синапсов • В синапсе происходит трансформация ритма возбуждения – частота импульсов в нерве меньше или больше частоты возбуждения постсинаптической мембраны нейрона. • Лабильность синапса существенно меньше, чем нерва. • Из всех звеньев рефлекторной дуги синапсы наиболее утомляемы и чувствительны к ядам и недостатку кислорода.

Свойства нервных центров • Центральная задержка рефлекса – характеризуется временем распространения информации в структурах нервного центра, г. о. в синапсах, где скорость проведения сигнала существенно меньше, чем в нервных волокнах. • Центральная задержка зависит от количества синапсов между нейронами центра и представляет собой сумму синаптических задержек.

Схема явления центрального облегчения
 При раздражении А возбуждаются")
Центральное облегчение При раздражении В возбуждаются 2 нейрона (5, 6) При раздражении А возбуждаются 2 нейрона (1, 2) А В 1 2 3 4 5 6 Клетки периферической каймы При раздражении А + В возбуждаются 6 нейронов (1, 2, 3, 4, 5, 6) Клетки центральной части нейронного пула

Схема явления окклюзии
 При раздражении")
Центральная окклюзия При раздражении А возбуждаются 4 нейрона (1, 2, 3, 4) При раздражении В возбуждаются 4 нейрона (3, 4, 5, 6) А В 1 2 3 4 5 6 Клетки центральной части нейронного пула При раздражении А + В возбуждаются 6 нейронов (1, 2, 3, 4, 5, 6)

Свойства нервных центров • Пластичность нервных центров – способность перестраивать функциональные свойства для более эффективной регуляции функций, осуществления новых, ранее несвойственных этому центру рефлексов или восстановления функций после повреждения части нейронов центра. • Пластичность обеспечивает изменение эффективности и направленности связей между нервными клетками, является рабочим механизмом обучения. • В основе пластичности лежат функциональные особенности синапсов и мембран нейронов ( «тренировка» синапсов, посттетаническая потенциация, периодичность функционирования синапсов и нейронов, пространственная и временная суммация постсинаптических потенциалов), наличие дублирующих систем нейронов и нервных волокон.
 • Характер и длительность восстановительных процессов могут изменяться в")
Принцип переключения (И. М. Сеченов) • Характер и длительность восстановительных процессов могут изменяться в зависимости от режима деятельности спортсменов в послерабочий, восстановительный, период. • В опытах И. М. Сеченова было показано, что в определенных условиях более быстрое и более значительное восстановление работоспособности обеспечивается не пассивным отдыхом, а переключением на другой вид деятельности, т. е. активным отдыхом. • В частности, он обнаружил, что работоспособность руки, утомленной работой на ручном эргографе, восстанавливалась быстрее и полнее, когда период отдыха ее был заполнен работой другой руки.
 • Анализируя этот феномен, И. М. Сеченов предположил, что")
Принцип переключения (И. М. Сеченов) • Анализируя этот феномен, И. М. Сеченов предположил, что афферентные импульсы, поступающие во время отдыха от других работающих мышц, способствуют лучшему восстановлению работоспособности нервных центров, как бы заряжая их энергией. • Кроме того, работа одной рукой вызывает увеличение кровотока в сосудах другой руки, что также может способствовать более быстрому восстановлению работоспособности утомленных мышц.

Принципы организации ЦНС Принцип общего конечного пути «Воронка» Ч. Шеррингтон Соотношение чувствительных и двигательных нейронов: 10: 1

ПРИНЦИП ОБЩЕГО КОНЕЧНОГО ПУТИ КОРА ПОДКОРКА СТВОЛ СПИННОЙ МОЗГ

Принцип цефализации и кортиколизации функций Чем выше уровень организации ЦНС, тем больше рефлекторных центров подчиняется коре больших полушарий. При этом спинальные двигательные программы не могут функционировать самостоятельно, но с помощью коры БП вовлекаются в разнообразные формы двигательной активности.

Свойства нервных центров • Утомление нервных центров – снижение эффективности их деятельности в виде повышения порогов возбуждения, связанное с утомлением синапсов и метаболическими сдвигами типа энергетического истощения в нервных клетках. • Утомление формируется при чрезмерной продолжительности действия раздражителей или их интенсивности, напряженном умственном труде или физической работе.
 Спинальный шок: Обратимое выключение спинальных рефлексов вследствие прекращения супраспинальных влияний. После")
Принцип иерархии (соподчинения) Спинальный шок: Обратимое выключение спинальных рефлексов вследствие прекращения супраспинальных влияний. После прекращения спинального шока спинальные рефлексы восстанавливаются, проводниковая функция спинного мозга – нет.
 • Доминанта - временно господствующий рефлекс")
ОПРЕДЕЛЕНИЕ ДОМИНАНТЫ ( по А. А. Ухтомскому, 1931) • Доминанта - временно господствующий рефлекс или поведенческий акт, которым трансформируется и направляется для данного времени прочих равных условиях работа прочих рефлекторных дуг, рефлекторного аппарата и поведения в целом.
 Свойства доминантного центра • Повышенная возбудимость. • Повышенная способность")
Принцип доминанты (А. А. Ухтомский) Свойства доминантного центра • Повышенная возбудимость. • Повышенная способность к суммации возбуждений. 3. Устойчивое длительное возбуждение. 4. Торможение других центров. Функция доминантного центра - осуществление данной поведенческой реакции для удовлетворения соответствующей потребности. Суммация возбуждений: 1. От рефлексогенных зон. 2. От экстерорецепторов (обстановочная импульсация). 3. Пусковая импульсация.

Свойства доминантного очага • Обладает повышенной возбудимостью. • Обладает стойкостью, инертностью возбуждения. • Обладает сопряженно тормозящим действием на другие центры. • Может иметь повышенную возбудимость под действием предварительных гуморальных влияний или слабых адекватных для него стимулов. • Становится способным к суммации возбуждений независимо от их модальности, т. е. при развитии доминанты посторонние для доминантного центра импульсы идут на подкрепление текущей установки.

ПРИНЦИП ДОМИНАНТЫ Раздражители Нервные центры Рефлексы

• Торможение – активный физиологический процесс, возникающий в ЦНС на основе возбуждения и приводящий либо к его снижению, либо к его полному прекращению. • Торможение подавляет возникновение и распространение возбуждения в элементах нервной системы и тем самым не позволяет реализоваться рефлекторным актам.

Торможение • И. М. Сеченов открыл эффект центрального торможения. • Он обнаружил, что химическое раздражение зрительных бугров тормозит простые спинномозговые безусловные рефлексы. • Экспериментально наличие специфических тормозных нейронов подтвердили Экклс, Реншоу: • обнаружили существование специфических вставочных нейронов, при активации которых происходило торможение двигательных нейронов.
 И. М. Сеченов исследовал сгибательный рефлекс лягушки (по методике Тюрка), который возникал в")
1) И. М. Сеченов исследовал сгибательный рефлекс лягушки (по методике Тюрка), который возникал в ответ на погружение лапы животного в раствор кислоты. • 2) Раздражение зрительного бугра (таламуса) лягушки кристаллами Na. Cl тормозило рефлекс – удлиняло времени сгибательного рефлекса. Это получило название сеченовского, или центрального, торможения.

Первичное торможение • Первичное торможение возникает в специальных тормозных клетках, примыкающих к тормозному нейрону. При этом тормозные нейроны выделяют соответствующие нейромедиаторы. • ПОСТСИНАПТИЧЕСКОЕ - основной вид первичного торможения, развивающийся в постсинаптической мембране аксо-соматических и аксо-дендритических синапсов под влиянием активации тормозных нейронов (клеток Реншоу) и вставочных нейронов. • При этом типе торможения происходит гиперполяризация постсинаптической мембраны, что и обуславливает торможение.

Первичное торможение • ВОЗВРАТНОЕ - нейрон воздействует на клетку Реншоу, которая в ответ тормозит этот же нейрон. • РЕЦИПРОКНОЕ - явление торможения нейронов мышцсгибателей при возбуждении эффекторных нейронов мышц-разгибателей через вставочный тормозной нейрон. • ЛАТЕРАЛЬНОЕ – тормозная клетка тормозит расположенные рядом нейроны. Подобные явления развиваются между биполярными и ганглиозными клетками сетчатки, что создает условия для более четкого видения предмета. • ВОЗВРАТНОЕ ОБЛЕГЧЕНИЕ – нейтрализация торможения нейрона при торможении тормозных клеток другими тормозными клетками. • ПРЕСИНАПТИЧЕСКОЕ - возникает в обычных нейронах, связано с процессом возбуждения.

Разновидности постсинаптического торможения

Постсинаптическое торможение • Приводит к урежению или полному прекращению генерации распространяющихся ПД. • В тормозных синапсах выделяются тормозные медиаторы (глицин, гамма-аминомасляная кислота). • Тормозной синапс может иметь тот же медиатор, что и возбуждающий, но иную природу рецепторов постсинаптической мембраны. • Тормозной эффект может быть достигнут при активации • калиевых каналов (К+ выходит наружу) и • хлорных каналов (Сl- входит внутрь клетки). • Происходит гиперполяризация постсинаптической мембраны, возникает ТПСП – тормозной постсинаптический потенциал.
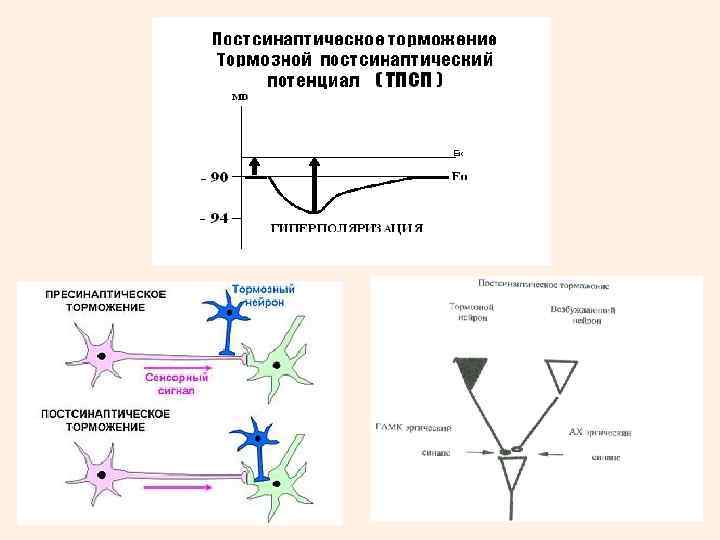

Фильтрация сенсорной информации посредством пресинаптического и постсинаптического торможения

• Пресинаптическое торможение • открыто в ЦНС сравнительно недавно, поэтому изучено меньше • заключается в торможении пресинаптической мембраны возбуждающих синапсов по аксональным синапсам. • На пресинаптических терминалях располагаются окончания аксонов других нервных клеток, образующих здесь аксо-аксональные синапсы. • Медиаторы их деполяризуют мембрану терминалей и приводят в состояние, подобное катодической депрессии Вериго. • Блокирует распространение возбуждения по аксону. • Свойственно для структур спинного мозга, ствола мозга. • Пресинаптическое торможение обычно длительное.
 высвобождается ГАМК, что приводит к активации")
Механизм пресинаптического торможения • В аксо-аксональном синапсе (А-1) высвобождается ГАМК, что приводит к активации хлорных каналов. • Из-за высокого потенциала покоя мембраны аксона ионы Сl+ выходят через мембрану наружу, что приводит к небольшой деполяризации постсинаптической мембраны, которая является одновременно участком мембраны другого аксона (А-2). • Деполяризация мембраны аксона А-2 ведет к инактивации натриевых каналов. • При прохождении нервного импульса по аксону А-2 этот участок мембраны не деполяризуется до КУД. Возникает блок проведения нервного импульса, он не поступает к синаптической области А-2, медиатор не выделяется.
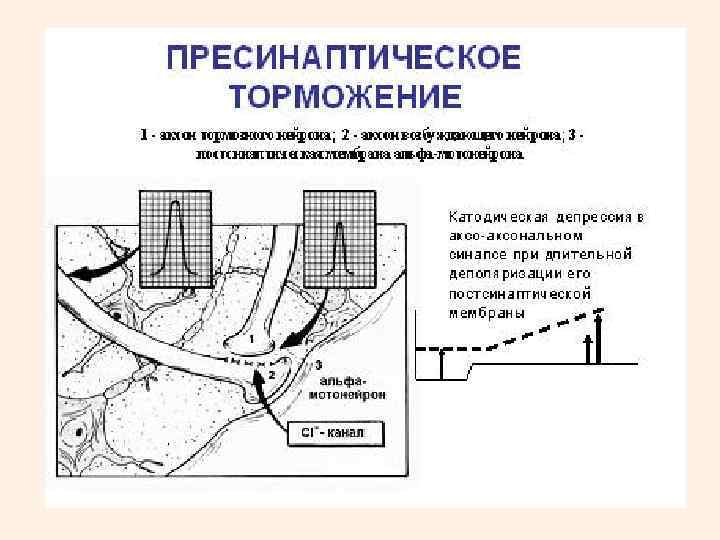

Аксо-аксональный синапс • Структура для пресинаптического торможения

Пресинаптическое торможение в ЦНС

Латеральное торможение • Латеральное торможение обеспечивает контрастность восприятия, для идентификации воспринимаемого объекта. • Проявляется в формировании вокруг зоны возбуждения зоны заторможенных нейронов по механизму одновременной индукции. Зона торможения превышает зону возбуждения в два раза. • Торможение возникает также в результате последовательной индукции – в нейронах коры после возбуждения всегда возникает торможение.

Схема латерального торможения в сенсорных системах. Тормозные интернейроны выделены черным цветом

• Латеральное торможение происходит вследствие активации тормозных интернейронов переключательного ядра коллатералями возбуждающих нейронов. • Чем сильнее возбужден релейный сенсорный нейрон, тем больше он активирует тормозные нейроны, которые подавляют активность соседних релейных нейронов. • Релейные нейроны, передающие сигналы от центра рецептивного поля, сильнее других повышают активность тормозных нейронов, а их влияние на соседние релейные нейроны делает еще большим контраст между возбужденными и тормозимыми нейронами, выделяя линию передачи избранных сигналов.

Возвратное торможение • осуществляет мотонейрон через клетку Реншоу, обеспечивая готовность воспринять новую команду от пирамидной клетки. • Проявляется в угнетении (подавлении) активности нейрона, вызываемом возвратной коллатералью аксона нервной клетки. • Так, мотонейрон переднего рога серого вещества спинного мозга перед выходом из спинного мозга дает боковую (возвратную) ветвь. Она возвращается назад и заканчивается на тормозных нейронах – клетках Реншоу. • Аксоны клеток Реншоу заканчиваются на мотонейронах, оказывая на них тормозное действие. • Это пример торможения по принципу обратной связи. • В такой цепи усиление возбуждения, поступающего к мышце, усиливает торможение мотонейрона под действием клетки Реншоу.

Возвратное торможение

• Реципрокное торможение развивается в центрах противоположной деятельности например, в центрах сгибателей при возбуждении центра разгибателей.
 иннервации")
Принцип реципрокной (взаимосочетанной) иннервации
, пресинаптическое (Б) и возвратное (В) торможение в локальных нейронных цепях спинного мозга")
Реципрокное (А), пресинаптическое (Б) и возвратное (В) торможение в локальных нейронных цепях спинного мозга 1 - мотонейрон, 2 - тормозный интернейрон, 3 афферентные терминали.
")
Принцип реципрокности (сопряженного торможения)

Вторичное торможение • Не требует специальных тормозных структур, возникает в результате изменения функциональной активности обычных возбудимых структур, всегда связано с процессом возбуждения. • Виды вторичного торможения: • а) запредельное, возникающее при большом потоке информации, поступающей в клетку. Поток информации лежит за пределами работоспособности нейрона; • б) пессимальное, возникающее при высокой частоте раздражения;
 парабиотическое, возникающее при сильно и длительно действующем раздражении; • г)")
Вторичное торможение • в) парабиотическое, возникающее при сильно и длительно действующем раздражении; • г) торможение вслед за возбуждением, возникающее вследствие снижения функционального состояния нейронов после возбуждения; • д) торможение по принципу отрицательной индукции; • е) торможение условных рефлексов.

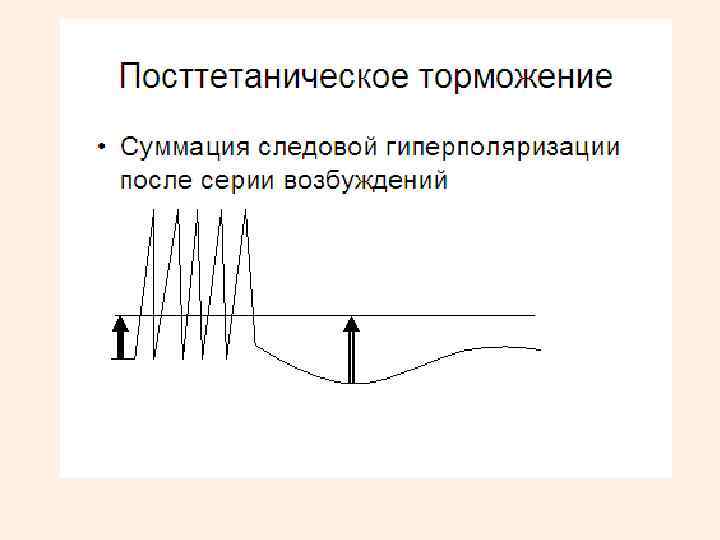
• Возбуждение вслед за торможением: Посттетаническое торможение • Особый вид торможения, которое возникает в случае, если после окончания возбуждения в клетке возникает сильная гиперполяризация мембраны. • Возбуждающий постсинаптический потенциал (ВПСП) в этих условиях оказывается недостаточным для критической деполяризации мембраны и генерации распространяющегося возбуждения. • Причина такого торможения в том, что следовые потенциалы способны к суммации, и после серии частых импульсов возникает суммация положительного следового потенциала.

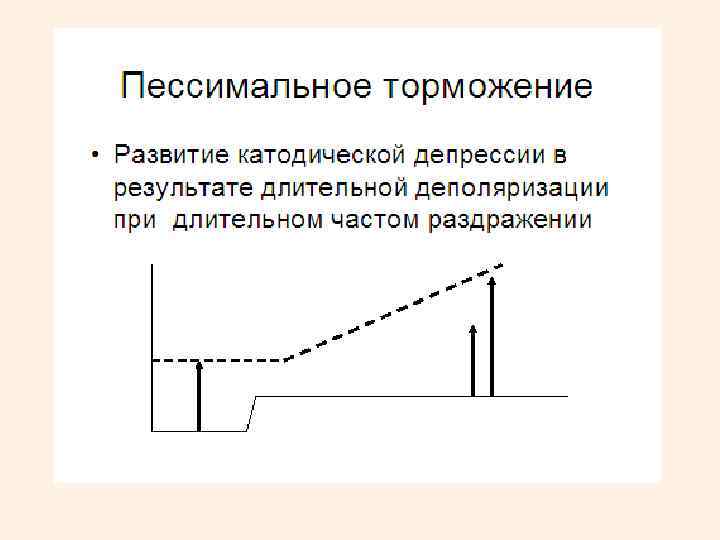
• Пессимальное торможение • возникает в возбуждающих синапсах в результате сильной деполяризации постсинаптической мембраны под влиянием слишком частых импульсов (как пессимум в нервно-мышечном препарате). • Первоначально ответное возбуждение имеет высокую частоту. Через некоторое время стимулируемый нейрон переходит в состояние торможения. • При избыточном поступлении нервных импульсов формируется стойкая деполяризация постсинаптической мембраны, приводящая к инактивации натриевых каналов. • К пессимальному торможению особо склонны промежуточные нейроны спинного мозга, нейроны РФ. • При стойкой деполяризации в них наступает состояние, подобное катодической депрессии Вериго.

Примеры нарушения торможения в ЦНС • НАРУШЕНИЕ ПОСТСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ: • СТРИХНИН БЛОКАДА РЕЦЕПТОРОВ ТОРМОЗНЫХ СИНАПСОВ • СТОЛБНЯЧНЫЙ ТОКСИН - НАРУШЕНИЕ ОСВОБОЖДЕНИЯ ТОРМОЗНОГО МЕДИАТОРА • НАРУШЕНИЕ ПРЕСИНАПТИЧЕСКОГО ТОРМОЖЕНИЯ: • ПИКРОТОКСИН БЛОКАДА ПРЕСИНАПТИЧЕСКИХ СИНАПСОВ

Интеграция и координация в деятельности ЦНС • Осуществляются на основе процессов конвергенции и дивергенции, обеспечивающих иррадиацию и концентрацию возбуждения. • В координационной деятельности нервных центров важную роль играет взаимодействие рефлексов, проявляющееся в различных эффектах.

Ответьте на вопросы задачи • Как известно, в деятельности ГМ имеет место процесс торможения. В процессе рассматривания сложного изображения или прослушивания музыкального фрагмента испытуемый выделяет их световые, цветовые и звуковые характеристики. • Вопросы. • 1. Дайте определение центрального торможения. • 2. Какие виды центрального торможения вам известны? • 3. Какой вид центрального торможения лежит в основе улучшения различения частоты звуков, выделения контуров изображения, дифференциации соседних точек прикосновения на коже? • Ответы. • 1. Физиологический процесс, возникающий в ЦНС на основе возбуждения и приводящий либо к уменьшению, либо к его полному прекращению. • 2. Возвратное, реципрокное, латеральное. • 3. Латеральное торможение.

Ответьте на вопросы задачи • Как известно, в деятельности головного мозга имеет место процесс торможения. В процессе рассматривания сложного изображения или прослушивания музыкального фрагмента испытуемый выделяет их световые, цветовые и звуковые характеристики. • Вопросы. • 1. Дайте определение центрального торможения. • 2. Какие виды центрального торможения вам известны? • 3. Какой вид центрального торможения лежит в основе улучшения различения частоты звуков, выделения контуров изображения, дифференциации соседних точек прикосновения на коже?

СПАСИБО ЗА ВНИМАНИЕ