

08_Гликолиз_окисление жирных кислот.pptx
- Количество слайдов: 28
 Пути получения энергии: гликолиз и окисление жирных кислот
Пути получения энергии: гликолиз и окисление жирных кислот
 Три стадии катаболических превращений основных питательн веществ в клетке
Три стадии катаболических превращений основных питательн веществ в клетке
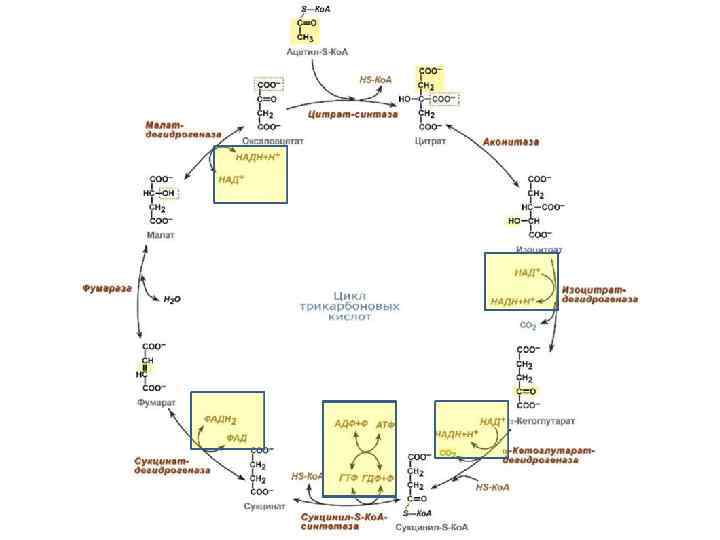
 Итого ц. Кребса: 2 NADH·H 2 FADH 2 2 ГТФ 2 NADH·H Итого АТФ: 2*3+2*2+2+2*3 = 24 38 ? ?
Итого ц. Кребса: 2 NADH·H 2 FADH 2 2 ГТФ 2 NADH·H Итого АТФ: 2*3+2*2+2+2*3 = 24 38 ? ?
; происходит в цитозоле. 1) Глюкоза") Гликолиз – бескислородное окисление глюкозы (получение энергии без кислорода); происходит в цитозоле. 1) Глюкоза - 2 молекулы молочной кислоты; 2) Глюкоза – пировиноградная кислота (ПВК) – спирт; 3) Глюкоза – ПВК + Co. A – ацетил-Co. A – ц. Кребса
Гликолиз – бескислородное окисление глюкозы (получение энергии без кислорода); происходит в цитозоле. 1) Глюкоза - 2 молекулы молочной кислоты; 2) Глюкоза – пировиноградная кислота (ПВК) – спирт; 3) Глюкоза – ПВК + Co. A – ацетил-Co. A – ц. Кребса
 С гликолизом сопряжен синтез АТР В ходе гликолиза значительная часть свободной энергии, содержащейся в молекуле глюкозы, запасается в форме АТР. Уравнение химического баланса для анаэробного гликолиза, протекающего в напряженно работающей скелетной мышце: На каждую расщепленную молекулу глюкозы из ADP и Р образуются две молекулы АТР.
С гликолизом сопряжен синтез АТР В ходе гликолиза значительная часть свободной энергии, содержащейся в молекуле глюкозы, запасается в форме АТР. Уравнение химического баланса для анаэробного гликолиза, протекающего в напряженно работающей скелетной мышце: На каждую расщепленную молекулу глюкозы из ADP и Р образуются две молекулы АТР.
 превращение глюкозы в лактат с высвобождением свободной") В анаэробном гликолизе выделяют два процесса 1) превращение глюкозы в лактат с высвобождением свободной энергии: 2) Образование АТР из ADP и фосфата, которое требует затраты энергии: Эти два процесса не могут идти независимо друг от друга; они обязательно должны быть сопряжены.
В анаэробном гликолизе выделяют два процесса 1) превращение глюкозы в лактат с высвобождением свободной энергии: 2) Образование АТР из ADP и фосфата, которое требует затраты энергии: Эти два процесса не могут идти независимо друг от друга; они обязательно должны быть сопряжены.
 !!!! Превращение 1 моль глюкозы в лактат в стандартных условиях приводит к высвобождению гораздо большего количества свободной энергии (47, 0 ккал), чем необходимо для образования 2 моль АТР из ADP и фосфата (2 • 7, 3 = + 14, 6 ккал). В живой клетке при нормальных внутриклеточных концентрациях АТР, ADP и Р, а также глюкозы и лактата эффективность запасания высвобождающейся при гликолизе энергии в форме АТР превышает 60%. Суммарное ΔG = -47, 0 + 14, 6 = = — 32, 4 ккал/моль, т. е. Суммарная сопряженная реакция гликолиза сопровождается очень большим снижением свободной энергии, поэтому в живых клетках гликолиз представляет собой необратимый процесс, идущий практически до конца.
!!!! Превращение 1 моль глюкозы в лактат в стандартных условиях приводит к высвобождению гораздо большего количества свободной энергии (47, 0 ккал), чем необходимо для образования 2 моль АТР из ADP и фосфата (2 • 7, 3 = + 14, 6 ккал). В живой клетке при нормальных внутриклеточных концентрациях АТР, ADP и Р, а также глюкозы и лактата эффективность запасания высвобождающейся при гликолизе энергии в форме АТР превышает 60%. Суммарное ΔG = -47, 0 + 14, 6 = = — 32, 4 ккал/моль, т. е. Суммарная сопряженная реакция гликолиза сопровождается очень большим снижением свободной энергии, поэтому в живых клетках гликолиз представляет собой необратимый процесс, идущий практически до конца.
 В продуктах гликолиза сохраняется еще много свободной энергии При гликолизе высвобождается только небольшая часть всей энергии, заключенной в молекуле глюкозы. Общее ΔG при полном окислении глюкозы до СО 2 и Н 2 О — 686 ккал/моль. При гликолитическом расщеплении глюкозы на две молекулы лактата ΔG = — 47, 0 ккал/моль, т. е. (47/686)· 100 = 6, 9% того количества энергии, которое может высвободиться при полном окислении глюкозы. Большая часть биологически доступной энергии, заключенной в молекуле глюкозы, сохраняется в продуктах гликолиза-двух молекулах лактата. Она может высвободиться только в том случае, если продукты гликолиза подвергнутся полному до СО 2 и Н 2 О молекулярным кислородом.
В продуктах гликолиза сохраняется еще много свободной энергии При гликолизе высвобождается только небольшая часть всей энергии, заключенной в молекуле глюкозы. Общее ΔG при полном окислении глюкозы до СО 2 и Н 2 О — 686 ккал/моль. При гликолитическом расщеплении глюкозы на две молекулы лактата ΔG = — 47, 0 ккал/моль, т. е. (47/686)· 100 = 6, 9% того количества энергии, которое может высвободиться при полном окислении глюкозы. Большая часть биологически доступной энергии, заключенной в молекуле глюкозы, сохраняется в продуктах гликолиза-двух молекулах лактата. Она может высвободиться только в том случае, если продукты гликолиза подвергнутся полному до СО 2 и Н 2 О молекулярным кислородом.
 Анаэробный гликолиз до стадии лактата нельзя считать малоэффективным процессом, в котором энергия расходуется неэкономно. Напротив, гликолиз - удивительный по своему совершенству процесс, поскольку он обеспечивает получение энергии из глюкозы без кислорода. В организме животных лактат, образующийся в работающих мышцах и диффундирующий в кровь, может возвращаться в цикл; он поступает в печень и здесь в период восстановления после напряженной мышечной работы вновь превращается в глюкозу.
Анаэробный гликолиз до стадии лактата нельзя считать малоэффективным процессом, в котором энергия расходуется неэкономно. Напротив, гликолиз - удивительный по своему совершенству процесс, поскольку он обеспечивает получение энергии из глюкозы без кислорода. В организме животных лактат, образующийся в работающих мышцах и диффундирующий в кровь, может возвращаться в цикл; он поступает в печень и здесь в период восстановления после напряженной мышечной работы вновь превращается в глюкозу.
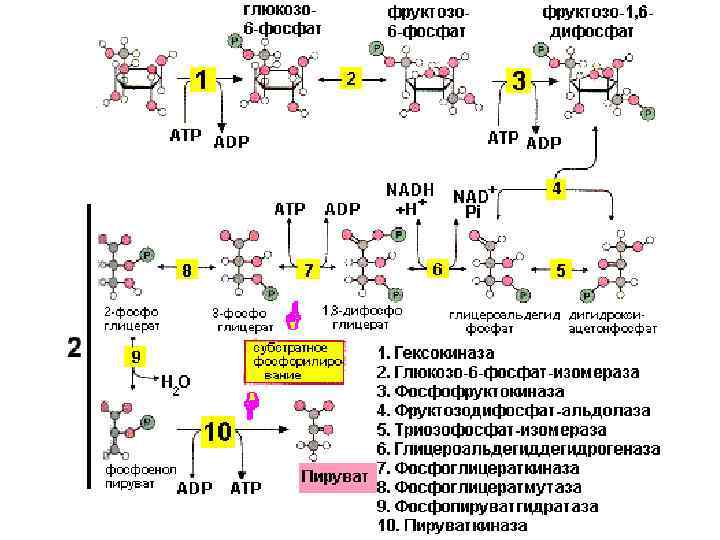
 Сопряженное фосфорилирование: Р. 7 Р. 10
Сопряженное фосфорилирование: Р. 7 Р. 10
 Восстановление пирувата до лактата В анаэробных условиях образовавшийся при гликолизе NADH реокисляется не за счет кислорода (который отсутствует), а за счет пирувата. восстанавливающегося при этом в лактат (лактатдегидрогеназа). Образуются 2 молекулы NAD, которые снова участвуют в гликолизе.
Восстановление пирувата до лактата В анаэробных условиях образовавшийся при гликолизе NADH реокисляется не за счет кислорода (который отсутствует), а за счет пирувата. восстанавливающегося при этом в лактат (лактатдегидрогеназа). Образуются 2 молекулы NAD, которые снова участвуют в гликолизе.
 Спиртовое брожение
Спиртовое брожение
 Энергетический баланс гликолиза: -2 АТФ +2 NADH*H +2 АТФ Итого: -2+2*3+2+2 = 8 АТФ (в аэробных условиях) Примечание: При аэробных условиях в животных тканях продуктом гликолиза является пируват, a NADH, образовавшийся в ходе окисления глицеральдегид-3 фосфата, реокисляется (т. е. снова превращается в NAD +) за счет молекулярного кислорода Итого: -2+2+2 = 2 АТФ (в анаэробных условиях) Примечание: При анаэробных условиях NADH расходуется до восстановления пирувата до лактата (или в реакциях спиртового брожения при образовании спирта)
Энергетический баланс гликолиза: -2 АТФ +2 NADH*H +2 АТФ Итого: -2+2*3+2+2 = 8 АТФ (в аэробных условиях) Примечание: При аэробных условиях в животных тканях продуктом гликолиза является пируват, a NADH, образовавшийся в ходе окисления глицеральдегид-3 фосфата, реокисляется (т. е. снова превращается в NAD +) за счет молекулярного кислорода Итого: -2+2+2 = 2 АТФ (в анаэробных условиях) Примечание: При анаэробных условиях NADH расходуется до восстановления пирувата до лактата (или в реакциях спиртового брожения при образовании спирта)
 Пируватдегидрогеназный комплекс Окислительное декарбоксилирование пирувата Итого: +2 NADH*H 6 АТФ
Пируватдегидрогеназный комплекс Окислительное декарбоксилирование пирувата Итого: +2 NADH*H 6 АТФ
 Окисление жирных кислот Триацилглицеролы Около 95% всей биологически доступной энергии в молекуле триацилглицеролов заключают в себе остатки трех жирных кислот с длинной цепью и только 5% приходится на долю остатка глицерола.
Окисление жирных кислот Триацилглицеролы Около 95% всей биологически доступной энергии в молекуле триацилглицеролов заключают в себе остатки трех жирных кислот с длинной цепью и только 5% приходится на долю остатка глицерола.
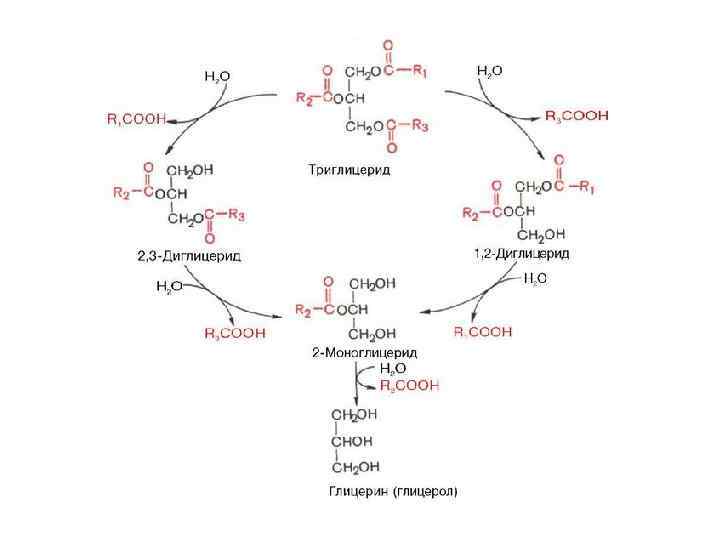
 Значительная часть образовавшегося при гидролизе") Окисление глицерина Обмен глицерина может осуществляться несколькими путями. 1) Значительная часть образовавшегося при гидролизе липидов глицерина используется для ресинтеза триглицеридов. 2) Второй путь обмена глицерина - включение продукта его окисления в гликолиз или в глюконеогенез.
Окисление глицерина Обмен глицерина может осуществляться несколькими путями. 1) Значительная часть образовавшегося при гидролизе липидов глицерина используется для ресинтеза триглицеридов. 2) Второй путь обмена глицерина - включение продукта его окисления в гликолиз или в глюконеогенез.
 Независимо от пути обмена начальным этапом является процесс фосфорилирования глицерина, донором фосфатной группы является молекула АТФ: Большая часть α-глицерофосфата используется для синтеза триглицеридов
Независимо от пути обмена начальным этапом является процесс фосфорилирования глицерина, донором фосфатной группы является молекула АТФ: Большая часть α-глицерофосфата используется для синтеза триглицеридов
 Р. 6 гликолиза
Р. 6 гликолиза
 Окисление жирных кислот В 1904 г. Ф. Кнооп показал, что в митохондрии в ходе окисления происходит постепенное уменьшение на два углеродных атома с карбоксильного конца высшей жирной кислоты. Ф. Кнооп назвал механизм окисления ВЖК β-окислением Первым этапом распада жирных кислот является их активирование; этот процесс катализируется ацил-Ко. Асинтетазой, которая локализована в мембранах эндоплазматической сети и в наружной мембране митохондрий:
Окисление жирных кислот В 1904 г. Ф. Кнооп показал, что в митохондрии в ходе окисления происходит постепенное уменьшение на два углеродных атома с карбоксильного конца высшей жирной кислоты. Ф. Кнооп назвал механизм окисления ВЖК β-окислением Первым этапом распада жирных кислот является их активирование; этот процесс катализируется ацил-Ко. Асинтетазой, которая локализована в мембранах эндоплазматической сети и в наружной мембране митохондрий:
 Поскольку процесс активирования ВЖК идет вне митохондрий, то далее необходим транспорт ацила через мембрану внутрь митохондрий. Транспорт происходит с участием находящегося на внешней стороне мембраны карнитина, на который передается ацил с ацил-Ко. А из цитоплазмы клетки. Затем ацилкарнитин диффундирует через мембрану митохондрии и передает свой ацил коэнзиму А, находящемуся в матриксе митохондрии. Перенос ацила между Ко. А и карнитином является ферментативным процессом, катализируемым ацил-Ко. А-карнитин-трансферазой
Поскольку процесс активирования ВЖК идет вне митохондрий, то далее необходим транспорт ацила через мембрану внутрь митохондрий. Транспорт происходит с участием находящегося на внешней стороне мембраны карнитина, на который передается ацил с ацил-Ко. А из цитоплазмы клетки. Затем ацилкарнитин диффундирует через мембрану митохондрии и передает свой ацил коэнзиму А, находящемуся в матриксе митохондрии. Перенос ацила между Ко. А и карнитином является ферментативным процессом, катализируемым ацил-Ко. А-карнитин-трансферазой
 В матриксе митохондрии происходит процесс β-окисления.
В матриксе митохондрии происходит процесс β-окисления.
 1 стадия: окисление ацил-Ко. А путем отщепления двух атомов водорода от α- и β-углеродных атомов ацила коферментом соответствующей дегидрогеназ. 2 стадия: присоединение молекулы воды таким образом, что ОНгруппа присоединяется к β-, а атом водорода - к αуглеродному атому:
1 стадия: окисление ацил-Ко. А путем отщепления двух атомов водорода от α- и β-углеродных атомов ацила коферментом соответствующей дегидрогеназ. 2 стадия: присоединение молекулы воды таким образом, что ОНгруппа присоединяется к β-, а атом водорода - к αуглеродному атому:
 3 стадия: окисление β-оксиацил-Ко. А, катализируемое дегидрогеназой, в результате действия которой образуется β-кетоацил-Ко. А: 4 стадия: негидролитический распад β-кетоацил-Ко. А и перенос ацила, укороченного на два углеродных атома по сравнению с первоначальным, на молекулу Ко. А:
3 стадия: окисление β-оксиацил-Ко. А, катализируемое дегидрогеназой, в результате действия которой образуется β-кетоацил-Ко. А: 4 стадия: негидролитический распад β-кетоацил-Ко. А и перенос ацила, укороченного на два углеродных атома по сравнению с первоначальным, на молекулу Ко. А:
 Образовавшийся ацетил-Ко. А подвергается окислению в цикле трикарбоновых кислот, а ацил-Ко. А, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил. Ко. А (4 -углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-Ко. А. Например, при окислении пальмитиновой кислоты (С 16) повторяется 7 циклов β-окисления. При окислении жирной кислоты, содержащей n углеродных атомов, происходит n/2– 1 цикл β-окисления (т. е. на один цикл меньше, чем n/2, так как при окислении бутирил-Ко. А сразу происходит образование 2 молекул ацетил-Ко. А).
Образовавшийся ацетил-Ко. А подвергается окислению в цикле трикарбоновых кислот, а ацил-Ко. А, укоротившийся на два углеродных атома, снова многократно проходит весь путь β-окисления вплоть до образования бутирил. Ко. А (4 -углеродное соединение), который в свою очередь окисляется до 2 молекул ацетил-Ко. А. Например, при окислении пальмитиновой кислоты (С 16) повторяется 7 циклов β-окисления. При окислении жирной кислоты, содержащей n углеродных атомов, происходит n/2– 1 цикл β-окисления (т. е. на один цикл меньше, чем n/2, так как при окислении бутирил-Ко. А сразу происходит образование 2 молекул ацетил-Ко. А).
 Энергетический баланс окисления жирных кислот (на примере пальмитиновой кислоты - C 16 H 32 O 2): + FADH 2 + NADH*H т. е. 2+3 = 5 АТФ (при каждом цикле β-окисления) C 16 H 32 O 2 - число циклов = 16/2 -1 = 7 т. е. 7*5 = 35 АТФ. 1 молекула ацетил-Ко. А в ц. Кребса = 12 АТФ В процессе β-окисления пальмитиновой кислоты образуется 8 молекул ацетил-Ко. А=8*12 =96 АТФ - АТФ (на образование активной формы пальмитиновой кислоты (пальмитоил-Ко. А) Итого= 35+96 -1 = 130 АТФ
Энергетический баланс окисления жирных кислот (на примере пальмитиновой кислоты - C 16 H 32 O 2): + FADH 2 + NADH*H т. е. 2+3 = 5 АТФ (при каждом цикле β-окисления) C 16 H 32 O 2 - число циклов = 16/2 -1 = 7 т. е. 7*5 = 35 АТФ. 1 молекула ацетил-Ко. А в ц. Кребса = 12 АТФ В процессе β-окисления пальмитиновой кислоты образуется 8 молекул ацетил-Ко. А=8*12 =96 АТФ - АТФ (на образование активной формы пальмитиновой кислоты (пальмитоил-Ко. А) Итого= 35+96 -1 = 130 АТФ