

Зоология позв. филогения.ppt
- Количество слайдов: 137
 Происхождение хордовых • • • , Время – предположительно начало Кембрийского периода а может, и еще раньше, в поздневендское время. Гипотеза № 1. Разработана французским сравнительным анатомом Жоффруа Сент-Илером еще в первой половине XIX века. Он считал, что членистые животные (кольчатые черви и членистоногие) могут рассматриваться как перевернутые позвоночные. В начале XX века идею происхождения хордовых от перевернувшихся на спинную сторону членистоногих отстаивали Гаскелл, Паттен и другие биологи. Гипотеза утратила свою популярность прежде всего в связи с тем, что стали ясны фундаментальные эмбриологические отличия между кольчатыми червями и членистоногими, с одной стороны, и хордовыми - с другой. Гипотеза № 2. В начале нынешнего столетия были разработаны гипотезы происхождения хордовых от кишечнодышащих (тип Гемихордовые (Hemichordata), класс Кишечнодышашие (Enteropneusta)). У этих животных (как и у хордовых) имеются жаберные щели, а в передней части тела - опорный вырост кишечника (стомохорд), который сравнивали с хордой. Однако кишечнодышащие резко отличались от хордовых обратным направлением тока крови, расположением сердца, строением нервной системы и другими важными особенностями организации. Делались попытки выводить хордовых от иглокожих, немертин, моллюсков и других групп беспозвоночных, однако ни одна из них не имела успеха.
Происхождение хордовых • • • , Время – предположительно начало Кембрийского периода а может, и еще раньше, в поздневендское время. Гипотеза № 1. Разработана французским сравнительным анатомом Жоффруа Сент-Илером еще в первой половине XIX века. Он считал, что членистые животные (кольчатые черви и членистоногие) могут рассматриваться как перевернутые позвоночные. В начале XX века идею происхождения хордовых от перевернувшихся на спинную сторону членистоногих отстаивали Гаскелл, Паттен и другие биологи. Гипотеза утратила свою популярность прежде всего в связи с тем, что стали ясны фундаментальные эмбриологические отличия между кольчатыми червями и членистоногими, с одной стороны, и хордовыми - с другой. Гипотеза № 2. В начале нынешнего столетия были разработаны гипотезы происхождения хордовых от кишечнодышащих (тип Гемихордовые (Hemichordata), класс Кишечнодышашие (Enteropneusta)). У этих животных (как и у хордовых) имеются жаберные щели, а в передней части тела - опорный вырост кишечника (стомохорд), который сравнивали с хордой. Однако кишечнодышащие резко отличались от хордовых обратным направлением тока крови, расположением сердца, строением нервной системы и другими важными особенностями организации. Делались попытки выводить хордовых от иглокожих, немертин, моллюсков и других групп беспозвоночных, однако ни одна из них не имела успеха.
 • • Гипотеза № 3. Неотеническая гипотеза, выводящая хордовых от личинок асцидий. Личинки асцидий имеют и хорду, и нервную трубку, которые утрачиваются при метаморфозе. На основании этого многие биологи (Берилл, Уитер, Боун и др. ) предполагают, что хордовые произошли от ставших половозрелыми личинок асцидий. Правда, в этом случае остается непонятным, откуда взялись сами асцидии. Да и трудно представить, чтобы хорда и нервная трубка возникли у живущих 1 - 2 дня и не питающихся личинок. Более вероятно, что эти особенности - наследие свободноживущих предков асцидий, об организации которых неотенические гипотезы ничего не говорят. Ни одна из рассмотренных гипотез в настоящее время не может считаться истинной, но в каждой из них содержатся отдельные элементы, которые могут оказаться полезными при реконструкции путей происхождения хордовых.
• • Гипотеза № 3. Неотеническая гипотеза, выводящая хордовых от личинок асцидий. Личинки асцидий имеют и хорду, и нервную трубку, которые утрачиваются при метаморфозе. На основании этого многие биологи (Берилл, Уитер, Боун и др. ) предполагают, что хордовые произошли от ставших половозрелыми личинок асцидий. Правда, в этом случае остается непонятным, откуда взялись сами асцидии. Да и трудно представить, чтобы хорда и нервная трубка возникли у живущих 1 - 2 дня и не питающихся личинок. Более вероятно, что эти особенности - наследие свободноживущих предков асцидий, об организации которых неотенические гипотезы ничего не говорят. Ни одна из рассмотренных гипотез в настоящее время не может считаться истинной, но в каждой из них содержатся отдельные элементы, которые могут оказаться полезными при реконструкции путей происхождения хордовых.
. А - Saccoglossus kowalevskyi, видно разделение тела на хоботок,") Строение кишечнодышащих Enteropneusta (по Шпенгелю). А - Saccoglossus kowalevskyi, видно разделение тела на хоботок, воротничок и туловище; Б - продольный разрез переднего конца Ptychodera minuta: 1 - жаберные щели, 2 - сплетение кровеносных сосудов, 3 - перикардий, 4 - хоботная пора, 5 - спинной нервный тяж, 6 - спинной сосуд, 7 - жаберная кишка, 8 - брюшной сосуд, 9 - пищевод, 10 - глотка, 11 - сердечная лакуна, 12 - нотохорд, 13 - продольные мускулы хоботка
Строение кишечнодышащих Enteropneusta (по Шпенгелю). А - Saccoglossus kowalevskyi, видно разделение тела на хоботок, воротничок и туловище; Б - продольный разрез переднего конца Ptychodera minuta: 1 - жаберные щели, 2 - сплетение кровеносных сосудов, 3 - перикардий, 4 - хоботная пора, 5 - спинной нервный тяж, 6 - спинной сосуд, 7 - жаберная кишка, 8 - брюшной сосуд, 9 - пищевод, 10 - глотка, 11 - сердечная лакуна, 12 - нотохорд, 13 - продольные мускулы хоботка
 был обнаружен в Южном Китае") Палеонтологические находки низших хордовых • • • Юннанозоон (Yunnanozoon) был обнаружен в Южном Китае в 1995 году. Одни специалисты рассматривали его как древнейшего представителя бесчерепных хордовых (Acrania), другие относили эту форму к полухордовым (Hemichordata). Хайкоуэлла (Haikouella) была обнаружена в 1999 году в Хайкоу (Китай). Тело хайкоуэллы было ланцетовидным, длиной 25 -40 мм, отчетливо выделялись голова, туловище и хвост. Брюшная часть туловища утолщена, спинная сжата с боков. Имелись плавники: спинной и небольшие брюшной и хвостовой. По бокам головы расположена пара округлых структур, которые интерпретировали как глаза. На брюшной стороне головы находилось ротовое отверстие, окруженное небольшими щупальцами и ведущее в объемистую глотку, боковые стенки которой пронизывались жаберными щелями и укреплялись шестью парами жаберных дуг. В глотке имелись конические зубовидные выросты, а по ее дну тянулся желобок - эндостиль, - признак, характерный для всех хордовых. Глотка переходила в узкий пищевод, соединявшийся с извитой средней кишкой, за ней следовала прямая задняя, которая заканчивалась анальным отверстием на брюшной стороне тела, у основания хвоста. Над кишечником хайкоуэллы лежала хорда, в туловище утолщенная и смещенная в нижнюю часть тела. Над ней располагался продольный нервный тяж, который в голове образовывал мозг из трех отделов. Под глоткой и над ней также находились тяжи, но китайские палеонтологи интерпретировали их как главные кровеносные сосуды - брюшную и спинную аорты. Их связывали жаберные артерии, а сама брюшная аорта начиналась от похожего на пузырь вздутия, которое могло быть сердцем. У хайкоуэллы спинную часть туловища над хордой занимали метамерные мышцы, сегменты которых (миомеры) разделялись S-образными перегородками - септами. По бокам средней кишки находились половые железы: четыре пары у хайкоуэллы и 13 пар у юннанозоона.
Палеонтологические находки низших хордовых • • • Юннанозоон (Yunnanozoon) был обнаружен в Южном Китае в 1995 году. Одни специалисты рассматривали его как древнейшего представителя бесчерепных хордовых (Acrania), другие относили эту форму к полухордовым (Hemichordata). Хайкоуэлла (Haikouella) была обнаружена в 1999 году в Хайкоу (Китай). Тело хайкоуэллы было ланцетовидным, длиной 25 -40 мм, отчетливо выделялись голова, туловище и хвост. Брюшная часть туловища утолщена, спинная сжата с боков. Имелись плавники: спинной и небольшие брюшной и хвостовой. По бокам головы расположена пара округлых структур, которые интерпретировали как глаза. На брюшной стороне головы находилось ротовое отверстие, окруженное небольшими щупальцами и ведущее в объемистую глотку, боковые стенки которой пронизывались жаберными щелями и укреплялись шестью парами жаберных дуг. В глотке имелись конические зубовидные выросты, а по ее дну тянулся желобок - эндостиль, - признак, характерный для всех хордовых. Глотка переходила в узкий пищевод, соединявшийся с извитой средней кишкой, за ней следовала прямая задняя, которая заканчивалась анальным отверстием на брюшной стороне тела, у основания хвоста. Над кишечником хайкоуэллы лежала хорда, в туловище утолщенная и смещенная в нижнюю часть тела. Над ней располагался продольный нервный тяж, который в голове образовывал мозг из трех отделов. Под глоткой и над ней также находились тяжи, но китайские палеонтологи интерпретировали их как главные кровеносные сосуды - брюшную и спинную аорты. Их связывали жаберные артерии, а сама брюшная аорта начиналась от похожего на пузырь вздутия, которое могло быть сердцем. У хайкоуэллы спинную часть туловища над хордой занимали метамерные мышцы, сегменты которых (миомеры) разделялись S-образными перегородками - септами. По бокам средней кишки находились половые железы: четыре пары у хайкоуэллы и 13 пар у юннанозоона.
 и реконструкции хайкоуэллы (слева) и юннанозоона. Это самые древние хордовые") Фоссилизированные остатки (верхний ряд) и реконструкции хайкоуэллы (слева) и юннанозоона. Это самые древние хордовые из когда-либо и гделибо найденных. 1 - передний конец тела, 2 - глаз, 3 - ротовая полость, 4 передний жаберный сосуд, 5 - мозг, 6 - жаберные щели, 7 - эндостиль, 8 зубовидные структуры, 9 - спинная аорта, 10 - глотка, 11 - брюшная аорта, 12 спинной плавник, 13 - сердце, 14 - пищевод, 15 - нервный тяж, 16 - половые железы, 17 - желудок, 18 - хорда, 19 - брюшной плавник, 20 - миосепты, 21 кишка, 22 - анальное отверстие, 23 - миомеры.
Фоссилизированные остатки (верхний ряд) и реконструкции хайкоуэллы (слева) и юннанозоона. Это самые древние хордовые из когда-либо и гделибо найденных. 1 - передний конец тела, 2 - глаз, 3 - ротовая полость, 4 передний жаберный сосуд, 5 - мозг, 6 - жаберные щели, 7 - эндостиль, 8 зубовидные структуры, 9 - спинная аорта, 10 - глотка, 11 - брюшная аорта, 12 спинной плавник, 13 - сердце, 14 - пищевод, 15 - нервный тяж, 16 - половые железы, 17 - желудок, 18 - хорда, 19 - брюшной плавник, 20 - миосепты, 21 кишка, 22 - анальное отверстие, 23 - миомеры.
 Современный ланцетник В среднем кембрии") Отпечаток ланцетникоподобного организма, Кембрий (около 550 млн. лет назад) Современный ланцетник В среднем кембрии (около 550 млн. лет назад) уже существовали бесчерепные, родственные современным ланцетникам. Их ископаемые остатки были обнаружены на территории США
Отпечаток ланцетникоподобного организма, Кембрий (около 550 млн. лет назад) Современный ланцетник В среднем кембрии (около 550 млн. лет назад) уже существовали бесчерепные, родственные современным ланцетникам. Их ископаемые остатки были обнаружены на территории США
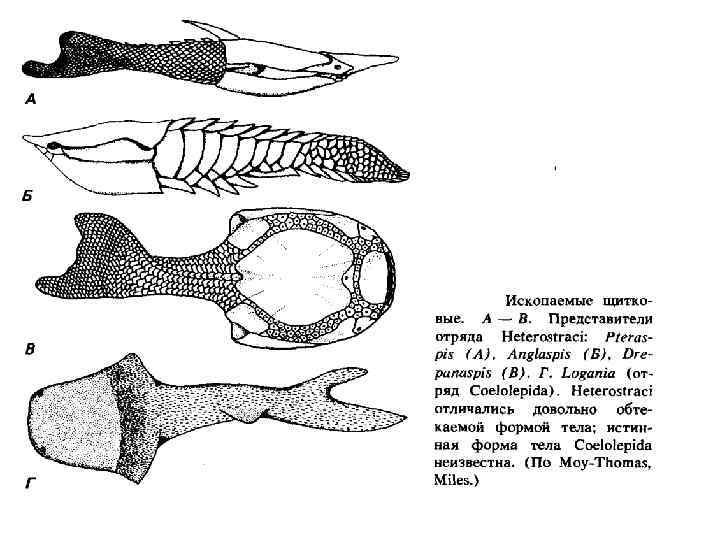
 Подтип Conodontochordata — Конодонтохордаты В морских отложениях с кембрия по триас часто находят мелкие зубцы — конодонты, построенные из костной ткани, характерной для представителей типа Хордовые. Эти зубцы очень разнообразны по форме. Поскольку конодонты быстро менялись со временем, их стали использовать в качестве руководящих ископаемых при исследовании палеозойских отложений.
Подтип Conodontochordata — Конодонтохордаты В морских отложениях с кембрия по триас часто находят мелкие зубцы — конодонты, построенные из костной ткани, характерной для представителей типа Хордовые. Эти зубцы очень разнообразны по форме. Поскольку конодонты быстро менялись со временем, их стали использовать в качестве руководящих ископаемых при исследовании палеозойских отложений.
 • Вначале палеонтологи думали, что конодонты находились на поверхности тела ранних хордовых, располагаясь примерно как чешуи. Потом стало ясно, что несколько конодонтов образовывали сложный конодонтовый аппарат из нескольких зубцов. И, наконец, были найдены останки конодонтоносителей, которые оказались угреобразными животными. • Вероятно, конодонтохордаты были фильтраторами и хищниками, населявшими толщу воды. Вероятно, в их крупной голове находились большие глаза, а в глотке размещался конодонтовый аппарат. Положение конодонтов в системе неясно. Некоторые авторы сближают конодонтохордат с миксинами.
• Вначале палеонтологи думали, что конодонты находились на поверхности тела ранних хордовых, располагаясь примерно как чешуи. Потом стало ясно, что несколько конодонтов образовывали сложный конодонтовый аппарат из нескольких зубцов. И, наконец, были найдены останки конодонтоносителей, которые оказались угреобразными животными. • Вероятно, конодонтохордаты были фильтраторами и хищниками, населявшими толщу воды. Вероятно, в их крупной голове находились большие глаза, а в глотке размещался конодонтовый аппарат. Положение конодонтов в системе неясно. Некоторые авторы сближают конодонтохордат с миксинами.
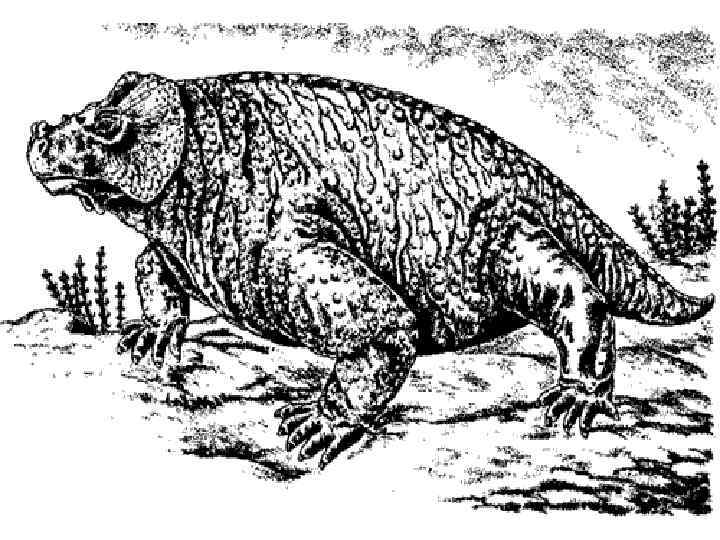
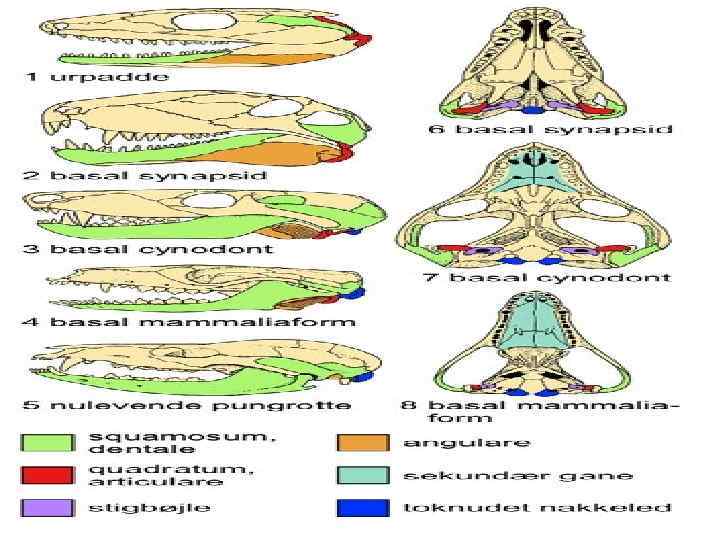
 Происхождение позвоночных • • Время появления – конец Ордовика – начало Силурийского периода. Предполагаемые предки – бесчерепные. Первая появившаяся группа позвоночных – представители раздела Бесчелюстные (Agnatha). Наиболее ранние известные представители позвоночных относятся к надклассу Щитковые (Ostracodermes). Надкласс включал в себя 2 класса: Птераспидоморфы (Pteraspidomorphi) и Цефаласпидоморфы (Cephalaspidomorphi) Класс Птераспидоморфы (Pteraspidomorphi) Имеется наружный скелет из пластин и крупных щитков, наиболее крупные пластины расположены на передней части тела. У некоторых представителей наружный скелет представлен разбросанными в коже шипами и мелкими пластинками. Глаза по бокам головы, обонятельные капсулы парные. Имелись зачаточные грудные плавники. Размеры от 3 см. до 1, 5 м. Известны из Силура – Девона Вероятные предки челюстноротых
Происхождение позвоночных • • Время появления – конец Ордовика – начало Силурийского периода. Предполагаемые предки – бесчерепные. Первая появившаяся группа позвоночных – представители раздела Бесчелюстные (Agnatha). Наиболее ранние известные представители позвоночных относятся к надклассу Щитковые (Ostracodermes). Надкласс включал в себя 2 класса: Птераспидоморфы (Pteraspidomorphi) и Цефаласпидоморфы (Cephalaspidomorphi) Класс Птераспидоморфы (Pteraspidomorphi) Имеется наружный скелет из пластин и крупных щитков, наиболее крупные пластины расположены на передней части тела. У некоторых представителей наружный скелет представлен разбросанными в коже шипами и мелкими пластинками. Глаза по бокам головы, обонятельные капсулы парные. Имелись зачаточные грудные плавники. Размеры от 3 см. до 1, 5 м. Известны из Силура – Девона Вероятные предки челюстноротых
 • • Наружный скелет сильно развит, голова покрыта сверху почти сплошным") Класс Цефаласпидоморфы (Cephalaspidomorphi) • • Наружный скелет сильно развит, голова покрыта сверху почти сплошным костным щитком, или голова и тело покрыты сверху несколькими рядами мелких пластинок, имеющих сверху слой дентина. Форма тела веретенообразная или сплющенная в дорзовентральном направлении. У некоторых вероятно имелись подвижные грудные плавники. Обонятельная капсула непарная. Жаберные мешки открывались наружу самостоятельными отверстиями. Имеются два подкласса: Костнощитковые (Osteostraci), и Бесщитковые (Anaspida). Бесщитковые включали мелких представителей с телом веретенообразной формы, грудными плавниками, обратно-гетероцеркальным хвостом. Тело было покрыто крупной чешуей, голова – мелкими костными пластинками. Возможно, являются предками миног и миксин (класс Круглоротые)
Класс Цефаласпидоморфы (Cephalaspidomorphi) • • Наружный скелет сильно развит, голова покрыта сверху почти сплошным костным щитком, или голова и тело покрыты сверху несколькими рядами мелких пластинок, имеющих сверху слой дентина. Форма тела веретенообразная или сплющенная в дорзовентральном направлении. У некоторых вероятно имелись подвижные грудные плавники. Обонятельная капсула непарная. Жаберные мешки открывались наружу самостоятельными отверстиями. Имеются два подкласса: Костнощитковые (Osteostraci), и Бесщитковые (Anaspida). Бесщитковые включали мелких представителей с телом веретенообразной формы, грудными плавниками, обратно-гетероцеркальным хвостом. Тело было покрыто крупной чешуей, голова – мелкими костными пластинками. Возможно, являются предками миног и миксин (класс Круглоротые)
 силурского периода") Щитковое (Anaspida, Pterolepis hitida) силурского периода
Щитковое (Anaspida, Pterolepis hitida) силурского периода
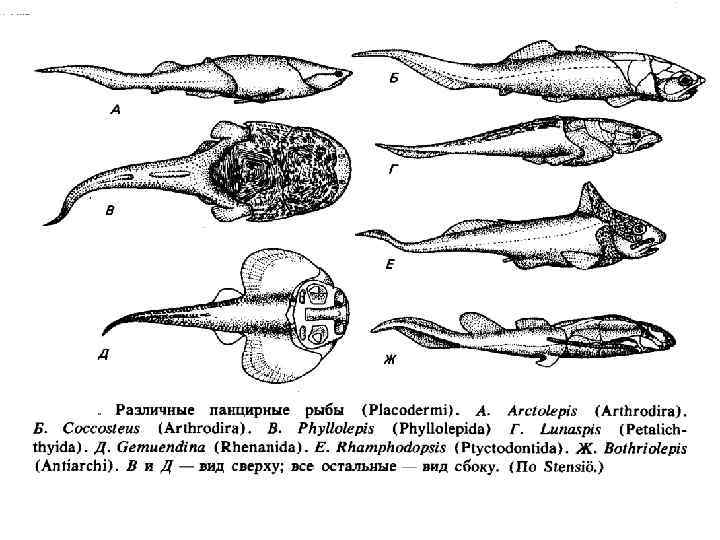
 Происхождение и эволюция рыб • Время появления – силур. В начале Девона уже имеется несколько различных групп рыб. • Предполагаемые предки – жившие в пресных или солоноватых водоемах птераспидоморфы. • Первым возникшими классами рыб видимо были Панцирные рыбы (Placodermi) и Челюстножаберные (Acanthodii) Панцирные рыбы (Placodermi) • Известны с раннего силура, расцвет пришелся на девон. В конце девонского периода вымерли. Ранние панцирные рыбы жили исключительно в пресной воде, но позже они переселились и в моря. • Характерным признаком панцирных рыб была защищённость головы и туловища костяным панцирем. Задняя часть туловища была покрыта чешуей или оставалась голой. Грудные плавники обычного типа или в виде костных шипов. У некоторых видов имелись брюшные плавники. Челюсти массивные, нередко окостеневали. Зубов на челюстях не было, но края челюстей имели острые грани. У некоторых на челюстях имелись жевательные пластины. Образовывались дуги позвонков. • Выделяется 2 подкласса: Артродиры (Arthrodira) и Антиархи (Antiarchi) • Размеры от 5 см. до 10 метров в длину. • Иногда плакодерм объединяют вместе с хрящевыми рыбами в один класс Elasmobranchiomorpha на правах подкласса.
Происхождение и эволюция рыб • Время появления – силур. В начале Девона уже имеется несколько различных групп рыб. • Предполагаемые предки – жившие в пресных или солоноватых водоемах птераспидоморфы. • Первым возникшими классами рыб видимо были Панцирные рыбы (Placodermi) и Челюстножаберные (Acanthodii) Панцирные рыбы (Placodermi) • Известны с раннего силура, расцвет пришелся на девон. В конце девонского периода вымерли. Ранние панцирные рыбы жили исключительно в пресной воде, но позже они переселились и в моря. • Характерным признаком панцирных рыб была защищённость головы и туловища костяным панцирем. Задняя часть туловища была покрыта чешуей или оставалась голой. Грудные плавники обычного типа или в виде костных шипов. У некоторых видов имелись брюшные плавники. Челюсти массивные, нередко окостеневали. Зубов на челюстях не было, но края челюстей имели острые грани. У некоторых на челюстях имелись жевательные пластины. Образовывались дуги позвонков. • Выделяется 2 подкласса: Артродиры (Arthrodira) и Антиархи (Antiarchi) • Размеры от 5 см. до 10 метров в длину. • Иногда плакодерм объединяют вместе с хрящевыми рыбами в один класс Elasmobranchiomorpha на правах подкласса.
 Окаменелость Bothriolepis panderi или астеролеписа
Окаменелость Bothriolepis panderi или астеролеписа
") Дунклеостей (Dunkleosteus)
Дунклеостей (Dunkleosteus)
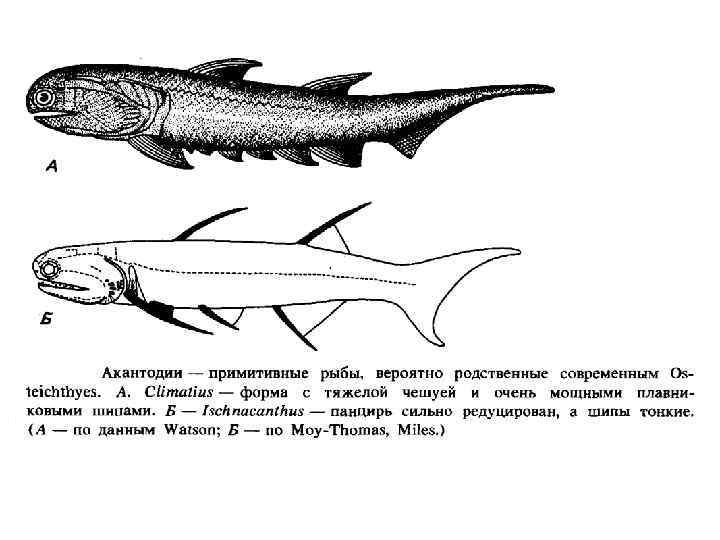
 • • • Возникли в Силуре, существовали до середины Пермского периода.") Класс Челюстножаберные (Acanthodii) • • • Возникли в Силуре, существовали до середины Пермского периода. Изначально населяли пресные подоемы, позднее проникли в моря. Размеры некрупные, тело веретенообразное. Голова и тело были покрыты костными пластинками и чешуями. Череп хрящевой, частично окостеневал. Челюстная дуга напоминала лежащие за ней жаберные дуги. Жаберные щели открывались самостоятельными отверстиями, у части видов имелись жаберные крышки. Имелась жаберная щель между челюстной и подъязычной дугой. Плавники поддерживались костными шипами. Парных плавников чаще всего более 2 -х пар. Иногда включаются в класс костные рыбы на правах подкласса.
Класс Челюстножаберные (Acanthodii) • • • Возникли в Силуре, существовали до середины Пермского периода. Изначально населяли пресные подоемы, позднее проникли в моря. Размеры некрупные, тело веретенообразное. Голова и тело были покрыты костными пластинками и чешуями. Череп хрящевой, частично окостеневал. Челюстная дуга напоминала лежащие за ней жаберные дуги. Жаберные щели открывались самостоятельными отверстиями, у части видов имелись жаберные крышки. Имелась жаберная щель между челюстной и подъязычной дугой. Плавники поддерживались костными шипами. Парных плавников чаще всего более 2 -х пар. Иногда включаются в класс костные рыбы на правах подкласса.
 и симфизная спираль Gomphonchus (справа)") Реконструкция головы Ischnacanthus (слева) и симфизная спираль Gomphonchus (справа)
Реконструкция головы Ischnacanthus (слева) и симфизная спираль Gomphonchus (справа)

 • Время возникновения – начало девона. Предполагаемые предки") Происхождение и эволюция хрящевых рыб (Chondrichthyes) • Время возникновения – начало девона. Предполагаемые предки – панцирные рыбы, или с ними имели общих предков. • С середины Девона известны морские акулоподобные рыбы кладоселахии. Размеры средние, тело веретенообразное, хвост гетероцеркальный. Имелась плакоидная чешуя. Скелет хрящевый, тела позвонков отсутствовали. Жаберных дуг более пяти. Рот находился на конце морды. Челюсти поддерживались подъязычной дугой. • В середине Девона от пресноводных кладоселяхий отделяется группа ксенокантид (Xenacanthida). Ксенакантиды были преимущественно пресноводными, населяя медленно текущие водотоки и мелководные озера. Скелет ксенаканта полностью хрящевой. Перед спинным плавником — длинный прямой шип. Спинной плавник один. Хвост дифицеркального типа. Анальных плавников два, парные плавники были с мясистыми основаниями. Жаберных дуг пять - семь. Челюсти амфистилические. Зубы ксенакантов имели две крупные вершины, направленные под углом друг к другу, и маленький зубчик, соединявшийся с зубом следующей генерации. Копулятивные органы есть. Вымерла эта группа в конце Перми – начале Триаса. • В середине Девона от кладоселяхий обособляются пластинчатожаберные. Уже в Юрском периоде пластинчатожаберные разделяются на акул и скатов. • С конца Девона известны первые цельноголовые. Палеозойских цельноголовых объединяют в группу брадиодонтов ( «древних скатов» ), известных с позднего девона до триаса.
Происхождение и эволюция хрящевых рыб (Chondrichthyes) • Время возникновения – начало девона. Предполагаемые предки – панцирные рыбы, или с ними имели общих предков. • С середины Девона известны морские акулоподобные рыбы кладоселахии. Размеры средние, тело веретенообразное, хвост гетероцеркальный. Имелась плакоидная чешуя. Скелет хрящевый, тела позвонков отсутствовали. Жаберных дуг более пяти. Рот находился на конце морды. Челюсти поддерживались подъязычной дугой. • В середине Девона от пресноводных кладоселяхий отделяется группа ксенокантид (Xenacanthida). Ксенакантиды были преимущественно пресноводными, населяя медленно текущие водотоки и мелководные озера. Скелет ксенаканта полностью хрящевой. Перед спинным плавником — длинный прямой шип. Спинной плавник один. Хвост дифицеркального типа. Анальных плавников два, парные плавники были с мясистыми основаниями. Жаберных дуг пять - семь. Челюсти амфистилические. Зубы ксенакантов имели две крупные вершины, направленные под углом друг к другу, и маленький зубчик, соединявшийся с зубом следующей генерации. Копулятивные органы есть. Вымерла эта группа в конце Перми – начале Триаса. • В середине Девона от кладоселяхий обособляются пластинчатожаберные. Уже в Юрском периоде пластинчатожаберные разделяются на акул и скатов. • С конца Девона известны первые цельноголовые. Палеозойских цельноголовых объединяют в группу брадиодонтов ( «древних скатов» ), известных с позднего девона до триаса.

") Ортакантус до 3, 5 м. Карбон-ранняя Пермь. Ксенакант (Xenacanthus)
Ортакантус до 3, 5 м. Карбон-ранняя Пермь. Ксенакант (Xenacanthus)
 Стефакантус-2 м длина, 350 млн лет.
Стефакантус-2 м длина, 350 млн лет.
 Отряд Евгенеодонты Helicoprion
Отряд Евгенеодонты Helicoprion
 Libanopristis Спатобатис, юрский период
Libanopristis Спатобатис, юрский период
 • Время возникновения – конец силура. Вероятные предки") Происхождение и эволюция костных рыб (OSTEICHTHYES) • Время возникновения – конец силура. Вероятные предки – челюстножаберные рыбы. • Ранние формы неизвестны. Уже в начале девона встречаются представители лопастеперых и лучеперых рыб. • Лучеперые рыбы возникли вероятнее всего в реках с быстрым течением. Наиболее ранние представители лучеперых относятся к надотряду Палеониски (Palaeonisci). Известны из среднего девона — раннего мела Европы, Сев. и Юж. Америки, Азии, Австралии. Расцвет в карбоне и перми. Дл. 30— 40 см. Тело разнообразной формы, покрыто ганоидной чешуёй. Верхнечелюстная кость неподвижно соединена с предкрышечной. Глаза и рот большие. Эндокраний окостеневший. Внутренний скелет частично окостеневший, тела позвонков не окостеневают. Хвостовой плавник гетероцеркальный, верх, лопасть его покрыта чешуёй. Передние края всех плавников с фулькрами (треугольными вильчатыми чешуями), образующими водорез, защищающий плавник. • Костистые рыбы возникли и эволюционировали вероятнее всего в морях. Костистые рыбы произошли скорее всего от костных ганоидов; ископаемые формы известны с триаса. Исходной группой костистых рыб скорее всего являлись рыбы надотряда Клюпеоидные (Clupeomorpha).
Происхождение и эволюция костных рыб (OSTEICHTHYES) • Время возникновения – конец силура. Вероятные предки – челюстножаберные рыбы. • Ранние формы неизвестны. Уже в начале девона встречаются представители лопастеперых и лучеперых рыб. • Лучеперые рыбы возникли вероятнее всего в реках с быстрым течением. Наиболее ранние представители лучеперых относятся к надотряду Палеониски (Palaeonisci). Известны из среднего девона — раннего мела Европы, Сев. и Юж. Америки, Азии, Австралии. Расцвет в карбоне и перми. Дл. 30— 40 см. Тело разнообразной формы, покрыто ганоидной чешуёй. Верхнечелюстная кость неподвижно соединена с предкрышечной. Глаза и рот большие. Эндокраний окостеневший. Внутренний скелет частично окостеневший, тела позвонков не окостеневают. Хвостовой плавник гетероцеркальный, верх, лопасть его покрыта чешуёй. Передние края всех плавников с фулькрами (треугольными вильчатыми чешуями), образующими водорез, защищающий плавник. • Костистые рыбы возникли и эволюционировали вероятнее всего в морях. Костистые рыбы произошли скорее всего от костных ганоидов; ископаемые формы известны с триаса. Исходной группой костистых рыб скорее всего являлись рыбы надотряда Клюпеоидные (Clupeomorpha).

") • Из лопастеперых рыб сильное распространение в девоне имеет надотряд Кистеперые рыбы (Crossopterygimorpha) представленный 2 отрядами – Рипидистиеобразные Osteolepidiformes (Rhipidistia) и Целакантообразные (Coelacanthifarmes). Целакантообразные обитали в морях, начиная с триаса постепенно начали вымирать. Рипидистиеобразные обитали в мелководных пресных водоемах. Форма тела их была торпедообразная, плавники короткие и сильные, имелись хоаны и легкие. От рипидистиеобразных произошли примитивные тетраподы. Вымерли рипидистиеобразные в начале перми. • Еще один отряд лопастеперых – Двоякодышащие (Dipneustomorpha) известен со среднего девона. Видимо двоякодышащие – потомки примитивных кистеперых, специализированные на питании моллюсками и ракообразными. Схема специализации челюстной мускулатуры у некоторых ископаемых саркоптеригий на примере Ectosteorhachis
• Из лопастеперых рыб сильное распространение в девоне имеет надотряд Кистеперые рыбы (Crossopterygimorpha) представленный 2 отрядами – Рипидистиеобразные Osteolepidiformes (Rhipidistia) и Целакантообразные (Coelacanthifarmes). Целакантообразные обитали в морях, начиная с триаса постепенно начали вымирать. Рипидистиеобразные обитали в мелководных пресных водоемах. Форма тела их была торпедообразная, плавники короткие и сильные, имелись хоаны и легкие. От рипидистиеобразных произошли примитивные тетраподы. Вымерли рипидистиеобразные в начале перми. • Еще один отряд лопастеперых – Двоякодышащие (Dipneustomorpha) известен со среднего девона. Видимо двоякодышащие – потомки примитивных кистеперых, специализированные на питании моллюсками и ракообразными. Схема специализации челюстной мускулатуры у некоторых ископаемых саркоптеригий на примере Ectosteorhachis
 Onychodus Platycephalichthys
Onychodus Platycephalichthys

 • Переход позвоночных от водного к наземному образу жизни") Происхождение и эволюция амфибий (Amphibia) • Переход позвоночных от водного к наземному образу жизни и появление земноводных сопровождалось появлением двух решающих приспособлений: дыхания кислородом атмосферы и передвижения по твёрдому субстрату, жаберное дыхание должно было замениться лёгочным, а конечности типа плавника — пятипалыми конечностями, представляющими собой многочленный рычаг, служащий для опоры тела о твёрдый субстрат. Приспособления к наземности независимо и параллельно развивались в нескольких линиях эволюции кистепёрых рыб отряда Рипидистиеобразные (Osteolepidiformes). Одной из наиболее «продвинутых» кистепёрых рыб являлся тиктаалик (Tiktaalik), имевший ряд переходных признаков, сближающих его с земноводными. К таким признакам относятся укороченный череп, отделенная от пояса передних конечностей и относительно подвижная голова, наличие локтевого и плечевого суставов. Плавник тиктаалика мог занимать несколько фиксированных положений, одно из которых предназначалось для того, чтобы животное могло находиться в приподнятом положении над грунтом (вероятно, чтобы «ходить» на мелководье). Тиктаалик дышал через ноздри, расположенные на конце плоской «крокодильей» морды. Воду, а возможно и атмосферный воздух в лёгкие нагнетали уже не жаберные крышки, а щёчные помпы. Некоторые из указанных приспособлений характерны также для кистепёрой рыбы пандерихтис (Panderichthys). Часто кистеперых рыб с явно наметившимися признаками «тетраподизации» выделяют в инфракласс Tetrapodomorpha.
Происхождение и эволюция амфибий (Amphibia) • Переход позвоночных от водного к наземному образу жизни и появление земноводных сопровождалось появлением двух решающих приспособлений: дыхания кислородом атмосферы и передвижения по твёрдому субстрату, жаберное дыхание должно было замениться лёгочным, а конечности типа плавника — пятипалыми конечностями, представляющими собой многочленный рычаг, служащий для опоры тела о твёрдый субстрат. Приспособления к наземности независимо и параллельно развивались в нескольких линиях эволюции кистепёрых рыб отряда Рипидистиеобразные (Osteolepidiformes). Одной из наиболее «продвинутых» кистепёрых рыб являлся тиктаалик (Tiktaalik), имевший ряд переходных признаков, сближающих его с земноводными. К таким признакам относятся укороченный череп, отделенная от пояса передних конечностей и относительно подвижная голова, наличие локтевого и плечевого суставов. Плавник тиктаалика мог занимать несколько фиксированных положений, одно из которых предназначалось для того, чтобы животное могло находиться в приподнятом положении над грунтом (вероятно, чтобы «ходить» на мелководье). Тиктаалик дышал через ноздри, расположенные на конце плоской «крокодильей» морды. Воду, а возможно и атмосферный воздух в лёгкие нагнетали уже не жаберные крышки, а щёчные помпы. Некоторые из указанных приспособлений характерны также для кистепёрой рыбы пандерихтис (Panderichthys). Часто кистеперых рыб с явно наметившимися признаками «тетраподизации» выделяют в инфракласс Tetrapodomorpha.
 380 миллионов лет назад, рыба Tiktaalik roseae
380 миллионов лет назад, рыба Tiktaalik roseae
.") • На территории Латвии в 1960 -м г. были обнаружены окаменелости вентастеги (Ventastega). Относятся они к верхнему девону. Реконструкция облика вентастеги примерно такая. На голове с лопатообразным тупым рылом сидели сдвинутые назад небольшие глаза. За глазами расположен теменной глаз, а вперед от глаз и до конца морды расширяется треугольная депрессия. В средней части этой депрессии имелся неокостеневший участок. Широкая челюсть была усажена мелкими острыми зубами, среди которых выделялись увеличенные «клыки» . На длинном теле — около метра-полутора в длину — имелись две пары коротеньких конечностей с пальцами (сколько пальцев — неизвестно) и сзади хвост с плавником, который поддерживали плавниковые лучи около 7 см в длину. По сравнению с другими девонскими четвероногими у него было увеличено дыхательное отверстие. • Одним из наиболее примитивных истинных тетрапод была позднедевонская акантостега (Acanthostega) из отложений восточной Гренландии. Акантостега достигала длины около 60 см. Конечности не имели запястий, что говорит о крайне низкой приспособленности к передвижению по суше, и на каждой из них насчитывалось 8 пальцев. Строение скелета указывает на наличие внутренних жабр. Слабые конечности, которые не смогли бы выдержать вес животного, и короткие рёбра, говорят о его преимущественно водном образе жизни.
• На территории Латвии в 1960 -м г. были обнаружены окаменелости вентастеги (Ventastega). Относятся они к верхнему девону. Реконструкция облика вентастеги примерно такая. На голове с лопатообразным тупым рылом сидели сдвинутые назад небольшие глаза. За глазами расположен теменной глаз, а вперед от глаз и до конца морды расширяется треугольная депрессия. В средней части этой депрессии имелся неокостеневший участок. Широкая челюсть была усажена мелкими острыми зубами, среди которых выделялись увеличенные «клыки» . На длинном теле — около метра-полутора в длину — имелись две пары коротеньких конечностей с пальцами (сколько пальцев — неизвестно) и сзади хвост с плавником, который поддерживали плавниковые лучи около 7 см в длину. По сравнению с другими девонскими четвероногими у него было увеличено дыхательное отверстие. • Одним из наиболее примитивных истинных тетрапод была позднедевонская акантостега (Acanthostega) из отложений восточной Гренландии. Акантостега достигала длины около 60 см. Конечности не имели запястий, что говорит о крайне низкой приспособленности к передвижению по суше, и на каждой из них насчитывалось 8 пальцев. Строение скелета указывает на наличие внутренних жабр. Слабые конечности, которые не смогли бы выдержать вес животного, и короткие рёбра, говорят о его преимущественно водном образе жизни.
 вентастега
вентастега
") реконструкция Acanthostega gunnari Ихтиостега (реконструкция)
реконструкция Acanthostega gunnari Ихтиостега (реконструкция)
 Ихтиостега
Ихтиостега
 так же была обнаружена в верхнедевонских отложениях восточной Гренландии.") • Ихтиостега (Ichthyostega ) так же была обнаружена в верхнедевонских отложениях восточной Гренландии. Ихтиостеги были около 1, 5 метров длиной и имели по семь пальцев на задних ногах. Точное количество пальцев на передних лапах пока не установлено, но вероятно, что их было тоже семь. На хвосте у них был плавник «рыбьего» типа поддерживаемый невральными и гемальными дугами. Ноздри располагались у нижнего края челюстей. Слезная кость примыкает к ноздре, но не к глазнице. Межвисочная кость отсутствует. Заднетеменная кость непарная. Челюстная кость соприкосается с квадратноскуловой. Сохраняются подкрышечные и предкрышечные кости. Носовые кости широкие. Глазницы овальные и располагаются в центральной части черепа. Кожа была покрыта мелкой рыбьей чешуей, сохранялся клейтрум. Вероятнее всего имелись внутренние жабры. Скорее всего была полностью водным животным. • Перечисленные тетраподы похожи друг на друга и относятся не только к одному отряду Ichthyostegalia, но и к одному семейству Ichthyostegidae. Череп ихтиостегалий достаточно специализован: в нем потеряны некоторые кости, которые сохраняются у значительно более поздних амфибий (утрачена межвисочная кость и заднетеменные кости слились в одну непарную кость).
• Ихтиостега (Ichthyostega ) так же была обнаружена в верхнедевонских отложениях восточной Гренландии. Ихтиостеги были около 1, 5 метров длиной и имели по семь пальцев на задних ногах. Точное количество пальцев на передних лапах пока не установлено, но вероятно, что их было тоже семь. На хвосте у них был плавник «рыбьего» типа поддерживаемый невральными и гемальными дугами. Ноздри располагались у нижнего края челюстей. Слезная кость примыкает к ноздре, но не к глазнице. Межвисочная кость отсутствует. Заднетеменная кость непарная. Челюстная кость соприкосается с квадратноскуловой. Сохраняются подкрышечные и предкрышечные кости. Носовые кости широкие. Глазницы овальные и располагаются в центральной части черепа. Кожа была покрыта мелкой рыбьей чешуей, сохранялся клейтрум. Вероятнее всего имелись внутренние жабры. Скорее всего была полностью водным животным. • Перечисленные тетраподы похожи друг на друга и относятся не только к одному отряду Ichthyostegalia, но и к одному семейству Ichthyostegidae. Череп ихтиостегалий достаточно специализован: в нем потеряны некоторые кости, которые сохраняются у значительно более поздних амфибий (утрачена межвисочная кость и заднетеменные кости слились в одну непарную кость).
 Тулерпетон во многих отношениях не похож на этих животных. Это животное около 60 сантиметров длиной, найденное в морских верхнедевонских отложениях в Тульской области. На передней конечности 6 пальцев, на задней, вероятно, 7 (или 6). Тулерпетон, видимо, сохранял связь плечевого пояса с черепом, характерную для рыб, в то время как, например, ихтиостега, эту связь утратила.
Тулерпетон во многих отношениях не похож на этих животных. Это животное около 60 сантиметров длиной, найденное в морских верхнедевонских отложениях в Тульской области. На передней конечности 6 пальцев, на задней, вероятно, 7 (или 6). Тулерпетон, видимо, сохранял связь плечевого пояса с черепом, характерную для рыб, в то время как, например, ихтиостега, эту связь утратила.
 Когда четвероногие действительно переходят к наземному перемещению, с ними происходят разительные изменения. Связь плечевого пояса с черепом разрывается. Голова оказывается балансиром, позволяющим поднять туловище в районе плечевого пояса над опорой.
Когда четвероногие действительно переходят к наземному перемещению, с ними происходят разительные изменения. Связь плечевого пояса с черепом разрывается. Голова оказывается балансиром, позволяющим поднять туловище в районе плечевого пояса над опорой.
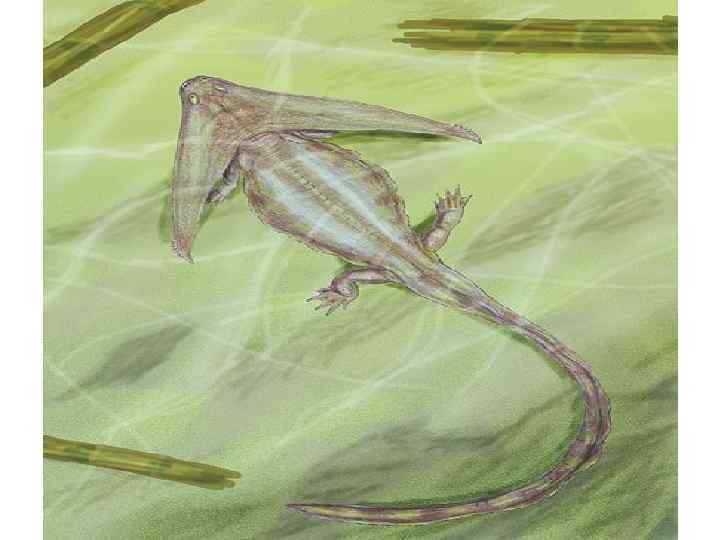

 • В дальнейшем, в каменноугольном периоде, возник ряд ветвей, которым придают таксономическое значение надотрядов или подклассов. Надотряд лабиринтодонтов (Labyrinthodontia) был очень разнообразен. Ранние формы имели сравнительно небольшие размеры и рыбообразное тело. Более поздние достигали весьма крупных размеров (1 м и более) в длину, тело их было уплощено и заканчивалось коротким толстым хвостом. Чешуя имелась на брюхе и боках тела. Лабиринтодонты существовали до конца триаса и занимали околоводные и водные местообитания. Питались преимущественно рыбой. К некоторым лабиринтодонтам относительно близки предки бесхвостых - отряд Proanura, известный с конца карбона и из пермских отложений.
• В дальнейшем, в каменноугольном периоде, возник ряд ветвей, которым придают таксономическое значение надотрядов или подклассов. Надотряд лабиринтодонтов (Labyrinthodontia) был очень разнообразен. Ранние формы имели сравнительно небольшие размеры и рыбообразное тело. Более поздние достигали весьма крупных размеров (1 м и более) в длину, тело их было уплощено и заканчивалось коротким толстым хвостом. Чешуя имелась на брюхе и боках тела. Лабиринтодонты существовали до конца триаса и занимали околоводные и водные местообитания. Питались преимущественно рыбой. К некоторым лабиринтодонтам относительно близки предки бесхвостых - отряд Proanura, известный с конца карбона и из пермских отложений.
. Они") • В карбоне возникла и вторая ветвь первичных амфибий — лепоспондилы (Lepospondyli). Они имели мелкие размеры и были хорошо приспособлены к жизни в воде. Некоторые из них вторично утратили конечности. Они просуществовали до середины пермского периода. В целом все палеозойские амфибии вымерли в течение триаса. Отдельные представители дожили до начала мезозоя.
• В карбоне возникла и вторая ветвь первичных амфибий — лепоспондилы (Lepospondyli). Они имели мелкие размеры и были хорошо приспособлены к жизни в воде. Некоторые из них вторично утратили конечности. Они просуществовали до середины пермского периода. В целом все палеозойские амфибии вымерли в течение триаса. Отдельные представители дожили до начала мезозоя.
 • Самые древние ископаемые остатки бесхвостых амфибий известны из нижнего триаса северной части Мадагаскара. Здесь найден единственный представитель особого отряда Proanura — Protobatrachus massinoti. У этого первичнобесхвостого, длина тела которого около 10 см, было 16 позвонков и все туловищные, за исключением первого, несли ребра. Между короткими подвздошными костями лежали 3 хвостовых позвонка. Локтевая и лучевая, а также большая и малая берцовые кости еще не были слиты. Начиная с верхней юры и нижнего мела появляются уже типичные бесхвостые амфибии различных подотрядов.
• Самые древние ископаемые остатки бесхвостых амфибий известны из нижнего триаса северной части Мадагаскара. Здесь найден единственный представитель особого отряда Proanura — Protobatrachus massinoti. У этого первичнобесхвостого, длина тела которого около 10 см, было 16 позвонков и все туловищные, за исключением первого, несли ребра. Между короткими подвздошными костями лежали 3 хвостовых позвонка. Локтевая и лучевая, а также большая и малая берцовые кости еще не были слиты. Начиная с верхней юры и нижнего мела появляются уже типичные бесхвостые амфибии различных подотрядов.

 Лепоспондилы Диплокаулус
Лепоспондилы Диплокаулус
 Pantilus Adelospondylus
Pantilus Adelospondylus
 Зуб лабиринтодонта, поперечный срез
Зуб лабиринтодонта, поперечный срез
 Лабиринтодонты Эриопс
Лабиринтодонты Эриопс
") Камакопс (Kamacops acervalis)
Камакопс (Kamacops acervalis)


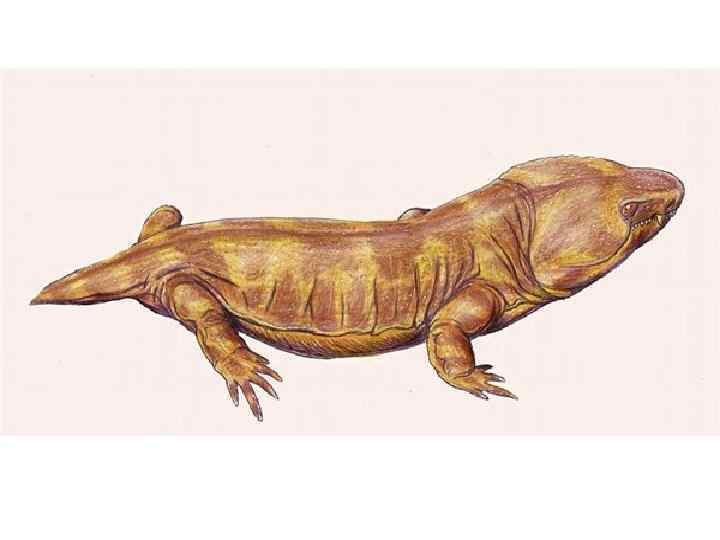
 дала начало рептилиям. Вероятно по") • Третья ветвь древних амфибий – Антракозавры (Anthracosauria) дала начало рептилиям. Вероятно по происхождению близки к лабиринтодонтам. Череп относительно высокий, иногда сжатый с боков. Сохраняется кинетизм черепа — как у кистепёрых рыб — подвижность между «щеками» и крышей черепа. Барабанной перепонки, видимо, нет, хотя существует вырезка по заднему краю щеки. Зубы обычно мощные, часто есть «клыки» на небе. Имелся один затылочный мыщелок. Тело удлиненное, хвост мощный, сжатый с боков. У некоторых описан «хвостовой плавник» из удлиненных супраневральных радиалий, как у рыб. Конечности у наземных форм короткие, но мощные, у водных форм — слабые. Обычно группу ограничивают 4— 5 семействами, существовавшими с самого начала карбона до середины перми. Иногда «антракозаврами» называют всех рептилиоморф. В середине пермского периода от антракозавров отделился отряд Seymouriamorpha, имеющий переходные признаки между амфибиями и рептилиями. Часто сеймуриоморф относят к таксону Рептилиоморф (Reptiliomorpha) или Батрахозавров (Batrachosauria).
• Третья ветвь древних амфибий – Антракозавры (Anthracosauria) дала начало рептилиям. Вероятно по происхождению близки к лабиринтодонтам. Череп относительно высокий, иногда сжатый с боков. Сохраняется кинетизм черепа — как у кистепёрых рыб — подвижность между «щеками» и крышей черепа. Барабанной перепонки, видимо, нет, хотя существует вырезка по заднему краю щеки. Зубы обычно мощные, часто есть «клыки» на небе. Имелся один затылочный мыщелок. Тело удлиненное, хвост мощный, сжатый с боков. У некоторых описан «хвостовой плавник» из удлиненных супраневральных радиалий, как у рыб. Конечности у наземных форм короткие, но мощные, у водных форм — слабые. Обычно группу ограничивают 4— 5 семействами, существовавшими с самого начала карбона до середины перми. Иногда «антракозаврами» называют всех рептилиоморф. В середине пермского периода от антракозавров отделился отряд Seymouriamorpha, имеющий переходные признаки между амфибиями и рептилиями. Часто сеймуриоморф относят к таксону Рептилиоморф (Reptiliomorpha) или Батрахозавров (Batrachosauria).
 Голова антракозавра Anthracosaurus russelli Личинка сеймуриоморфной амфибии Ariekanerpeton sigalovi
Голова антракозавра Anthracosaurus russelli Личинка сеймуриоморфной амфибии Ariekanerpeton sigalovi
 • Время появления – середина – конец каменноугольного периода.") Происхождение и эволюция рептилий (Reptilia) • Время появления – середина – конец каменноугольного периода. Вероятные предки – амфибии из группы Антракозавров (Anthracosauria). • В среднем карбоне от антракозавров обособилась новая ветвь – Рептилиоморфы (Reptiliomorpha) или Батрахозавры (Batrachosauria). Они обладали переходными признаками между амфибиями и рептилиями. Начиналось преобразование первых 2 -х позвонков в атлант и эпистрофей, имелся 1 затылочный мыщелок. Вероятно уже имелась грудная клетка и реберное дыхание. Размножались с типичной для амфибий стадией водной личинки. Когда предки рептилий приобрели способность размножаться на суше неизвестно, вероятно это произошло в конце карбона. • С конца каменноугольного периода появляется группа Парарептилий (Parareptilia). У большинства парарептилий наблюдаются признаки, характерные для земноводных. Наиболее существенные: череп без височных впадин с хорошо развитым теменным отверстием; нёбные зубы; двояковогнутые позвонки; очень короткий шейный отдел; короткие и массивные кости конечностей и их поясов. Кожные покровы парарептилий сохраняли много характерных для амфибий кожных желез. Группа достигла рассвета в Перми и вымерла в середине триаса. Имелись и некрупные ящерицеподобные формы (проколофоны) и крупные растительноядные (парейазавры). Вероятнее всего парарептилии уже имели амниотическое яйцо и были способны к размножению на суше.
Происхождение и эволюция рептилий (Reptilia) • Время появления – середина – конец каменноугольного периода. Вероятные предки – амфибии из группы Антракозавров (Anthracosauria). • В среднем карбоне от антракозавров обособилась новая ветвь – Рептилиоморфы (Reptiliomorpha) или Батрахозавры (Batrachosauria). Они обладали переходными признаками между амфибиями и рептилиями. Начиналось преобразование первых 2 -х позвонков в атлант и эпистрофей, имелся 1 затылочный мыщелок. Вероятно уже имелась грудная клетка и реберное дыхание. Размножались с типичной для амфибий стадией водной личинки. Когда предки рептилий приобрели способность размножаться на суше неизвестно, вероятно это произошло в конце карбона. • С конца каменноугольного периода появляется группа Парарептилий (Parareptilia). У большинства парарептилий наблюдаются признаки, характерные для земноводных. Наиболее существенные: череп без височных впадин с хорошо развитым теменным отверстием; нёбные зубы; двояковогнутые позвонки; очень короткий шейный отдел; короткие и массивные кости конечностей и их поясов. Кожные покровы парарептилий сохраняли много характерных для амфибий кожных желез. Группа достигла рассвета в Перми и вымерла в середине триаса. Имелись и некрупные ящерицеподобные формы (проколофоны) и крупные растительноядные (парейазавры). Вероятнее всего парарептилии уже имели амниотическое яйцо и были способны к размножению на суше.
 проколофон Bradysaurus Скутозавр
проколофон Bradysaurus Скутозавр
 • Черепахи возникли как околоводные или полуводные животные, лишь позднее появились наземные формы. Древние формы мало отличались от современных. Предки точно не известны, есть гипотезы о происхождении черепах от парейазавров, капториноморфов, плакодонтов и др. , но данные ДНК-анализа показывают близость черепах к диапсидным рептилиям. Первые известные науке черепахи Proganochelys quenstedi и Odontochelys появились в Триасовый период. Бокошейные черепахи обособились в конце триаса, скрытошейные – в начале юры. Известно множество ископаемых черепах, среди которых крупнейшей наземной черепахой была меиолания, с длиной панциря около 2, 5 м. Самой большой из известных науке черепах является жившая в меловом периоде и обитавшая в море черепаха Archelon ischyros. Размер одного из обнаруженных скелетов этого вида достигает более 4 метров
• Черепахи возникли как околоводные или полуводные животные, лишь позднее появились наземные формы. Древние формы мало отличались от современных. Предки точно не известны, есть гипотезы о происхождении черепах от парейазавров, капториноморфов, плакодонтов и др. , но данные ДНК-анализа показывают близость черепах к диапсидным рептилиям. Первые известные науке черепахи Proganochelys quenstedi и Odontochelys появились в Триасовый период. Бокошейные черепахи обособились в конце триаса, скрытошейные – в начале юры. Известно множество ископаемых черепах, среди которых крупнейшей наземной черепахой была меиолания, с длиной панциря около 2, 5 м. Самой большой из известных науке черепах является жившая в меловом периоде и обитавшая в море черепаха Archelon ischyros. Размер одного из обнаруженных скелетов этого вида достигает более 4 метров
 Odontochelys
Odontochelys
 Proganochelys quenstedi
Proganochelys quenstedi
 Archelon ischyros
Archelon ischyros
 • В середине - конце карбона появляются примитивные рептилии, близкие к стволу диапсидных рептилий (капториноморфные котилозавры). Это Капториниды, Болорозавры и Миллерозавры. Череп чаще анапсидный, без окон, но у некоторых (например, болозавров) появляется нижнее височное окно. Ушная вырезка отсутствует, но у миллерозавров имеется ушная вырезка, образованная, как и у диапсид квадратноскуловой и чешуйчатой костями. Череп часто крупный, особенно большеголовостью отличаются собственно капторины. У примитивных форм зубы конические, острые, однорядные, увеличены зубы в передней и средней части челюсти - животные были скорее всего насекомоядными или всеядными. У более поздних Captorhinidae щечные зубы становятся многорядными (одновременно функционировало до 12 рядов зубов), часто уплощенными - для таких форм предполагается моллюскоядность и растительноядность. У специализированных растительноядных - болозавров - зубы имеют булавовидные, долотовидные или расширенные и уплощенные коронки. • Представители отряда мезозавры появляются в конце карбона. Некрупные крокодилоподобные рептилии, длина до метра. Зубы длинные и тонкие, конечности превращены в ласты. В конце карбона – начале перми населяли водоемы, питались, видимо, рыбой.
• В середине - конце карбона появляются примитивные рептилии, близкие к стволу диапсидных рептилий (капториноморфные котилозавры). Это Капториниды, Болорозавры и Миллерозавры. Череп чаще анапсидный, без окон, но у некоторых (например, болозавров) появляется нижнее височное окно. Ушная вырезка отсутствует, но у миллерозавров имеется ушная вырезка, образованная, как и у диапсид квадратноскуловой и чешуйчатой костями. Череп часто крупный, особенно большеголовостью отличаются собственно капторины. У примитивных форм зубы конические, острые, однорядные, увеличены зубы в передней и средней части челюсти - животные были скорее всего насекомоядными или всеядными. У более поздних Captorhinidae щечные зубы становятся многорядными (одновременно функционировало до 12 рядов зубов), часто уплощенными - для таких форм предполагается моллюскоядность и растительноядность. У специализированных растительноядных - болозавров - зубы имеют булавовидные, долотовидные или расширенные и уплощенные коронки. • Представители отряда мезозавры появляются в конце карбона. Некрупные крокодилоподобные рептилии, длина до метра. Зубы длинные и тонкие, конечности превращены в ласты. В конце карбона – начале перми населяли водоемы, питались, видимо, рыбой.
 Labidosaurikos Belebey vegrandis Мезозавр
Labidosaurikos Belebey vegrandis Мезозавр
 – отряд ихтиозавры. Появились в триасе, существовали до") • Представители подкласса Ихтиоптеригии (Ichthyopterygia) – отряд ихтиозавры. Появились в триасе, существовали до верхнего мела. Форма тела рыбообразная, на конце хвостового стебля имелся вертикальный хвостовой плавник, чаще всего гипоцеркальный. Череп эвриапсидный. Парные конечности превращены в ласты. Ноздри и хоаны сдвинуты назад. Глаза крупные, имеется склеральное кольцо. На берег не выходили, рождали в воде живых детенышей. Питались рыбой, аммонитами, мелкими морскими рептилиями. Экскалибозавр
• Представители подкласса Ихтиоптеригии (Ichthyopterygia) – отряд ихтиозавры. Появились в триасе, существовали до верхнего мела. Форма тела рыбообразная, на конце хвостового стебля имелся вертикальный хвостовой плавник, чаще всего гипоцеркальный. Череп эвриапсидный. Парные конечности превращены в ласты. Ноздри и хоаны сдвинуты назад. Глаза крупные, имеется склеральное кольцо. На берег не выходили, рождали в воде живых детенышей. Питались рыбой, аммонитами, мелкими морскими рептилиями. Экскалибозавр
 Шонизавр Платиптеригий
Шонизавр Платиптеригий
. Существовал подкласс до") • Так же в конце карбона появляется подкласс Завроптеригии (Sauropterygia). Существовал подкласс до мелового периода. Наиболее значимыми представителями подкласса являлись плезиозавры, нотозавры и плакодонты. Череп эвриапсидный. Водные рептилии, конечности превращены в ласты, у многих видов плезиозавров имелась непропорционально длинная шея. По суше большинство видов плезиозавров передвигаться не могло. Нотозавры вероятнее всего являлись предками плезиозавров и вели прибрежно-водный образ жизни. Плезиозавры и нотозавры являлись хищниками. Плакодонты имели панцирь и были растительноядны. Muraenosaurus
• Так же в конце карбона появляется подкласс Завроптеригии (Sauropterygia). Существовал подкласс до мелового периода. Наиболее значимыми представителями подкласса являлись плезиозавры, нотозавры и плакодонты. Череп эвриапсидный. Водные рептилии, конечности превращены в ласты, у многих видов плезиозавров имелась непропорционально длинная шея. По суше большинство видов плезиозавров передвигаться не могло. Нотозавры вероятнее всего являлись предками плезиозавров и вели прибрежно-водный образ жизни. Плезиозавры и нотозавры являлись хищниками. Плакодонты имели панцирь и были растительноядны. Muraenosaurus
 Нотозавр. Henodus chelyops Пистозавр Лиоплеврадон Плезиозавр
Нотозавр. Henodus chelyops Пистозавр Лиоплеврадон Плезиозавр
 • В конце карбона - начале перми появилась крупная ветвь диапсидных пресмыкающихся. Наиболее примитивные диапсидные – эозухии. Размеры мелкие (до 0, 5 м. ), внешне напоминали ящериц. Позвонки амфицельные. Мелкие зубы имелись на челюстях и небных костях. Вымерли в начале триаса. • В перми от эозухий отделился отряд Клювоголовые. Размеры до 50 см. Имелись насекомоядные, растительноядные и водные формы. Большая часть клювоголовых вымерли в начале юры. • В конце перми от примитивных диапсид обособился отряд Чешуйчатые. Уже в меловой период ящерицы были многочисленны и разнообразны. Сильное распространение получило семейство мозозавры, близкое к варанам. Населяли моря, охотились на рыб, аммонитов и морских рептилий. Были яйцеживородящи. Змеи появились в конце мелового периода. Одной из самых примитивных форм был пахирахис. Существует две гипотезы происхождения змей – водная и наземная. По водной гипотезе змеи произошли от ящериц, близких к мозозаврам. По наземной гипотезе предками змей были какие-то роющие ящерицы.
• В конце карбона - начале перми появилась крупная ветвь диапсидных пресмыкающихся. Наиболее примитивные диапсидные – эозухии. Размеры мелкие (до 0, 5 м. ), внешне напоминали ящериц. Позвонки амфицельные. Мелкие зубы имелись на челюстях и небных костях. Вымерли в начале триаса. • В перми от эозухий отделился отряд Клювоголовые. Размеры до 50 см. Имелись насекомоядные, растительноядные и водные формы. Большая часть клювоголовых вымерли в начале юры. • В конце перми от примитивных диапсид обособился отряд Чешуйчатые. Уже в меловой период ящерицы были многочисленны и разнообразны. Сильное распространение получило семейство мозозавры, близкое к варанам. Населяли моря, охотились на рыб, аммонитов и морских рептилий. Были яйцеживородящи. Змеи появились в конце мелового периода. Одной из самых примитивных форм был пахирахис. Существует две гипотезы происхождения змей – водная и наземная. По водной гипотезе змеи произошли от ящериц, близких к мозозаврам. По наземной гипотезе предками змей были какие-то роющие ящерицы.
 плеврозавр пахирахис мозозавр Najash rionegrina
плеврозавр пахирахис мозозавр Najash rionegrina
. Череп архозавров") • Наиболее разнообразным и распространенным в мезозое был подкласс Архозавры (Archosauria). Череп архозавров диапсидного типа. Зубы сидят в альвеолах. Исходная группа архозавров – текодонты. Они возникли в конце перми и получили наибольшее развитие в триасе. Возможно являлись полифилетической группой. К текодонтам относились протерозухии, паразухии, псевдозухии Размеры их были от 15 см. до 3 – 5 м. Вели наземный и древесный образ жизни. Передвигались чаще на 4 конечностях, но для некоторых (орнитозухий) была характерна бипедальная локомоция. Между глазницей и ноздрей в крыше черепа имеется еще одно, так называемое суборбитальное или предглазничное, отверстие.
• Наиболее разнообразным и распространенным в мезозое был подкласс Архозавры (Archosauria). Череп архозавров диапсидного типа. Зубы сидят в альвеолах. Исходная группа архозавров – текодонты. Они возникли в конце перми и получили наибольшее развитие в триасе. Возможно являлись полифилетической группой. К текодонтам относились протерозухии, паразухии, псевдозухии Размеры их были от 15 см. до 3 – 5 м. Вели наземный и древесный образ жизни. Передвигались чаще на 4 конечностях, но для некоторых (орнитозухий) была характерна бипедальная локомоция. Между глазницей и ноздрей в крыше черепа имеется еще одно, так называемое суборбитальное или предглазничное, отверстие.
") Орнитозух Скелет Рутиодона Архозавр (Archosaurus rossicus)
Орнитозух Скелет Рутиодона Архозавр (Archosaurus rossicus)
 • В конце триаса от группы текодонтов отделилась группа крокодиломорф. Наиболее ранние представители крокодиломорф - протозухии. В юре крокодилы образовали большое количество форм в различных водоемах, в том числе в морях. Морские крокодилы имели ластоподобные конечности и хвостовой плавник. Имелись сухопутные крокодилы. Протозух Дакозавр
• В конце триаса от группы текодонтов отделилась группа крокодиломорф. Наиболее ранние представители крокодиломорф - протозухии. В юре крокодилы образовали большое количество форм в различных водоемах, в том числе в морях. Морские крокодилы имели ластоподобные конечности и хвостовой плавник. Имелись сухопутные крокодилы. Протозух Дакозавр
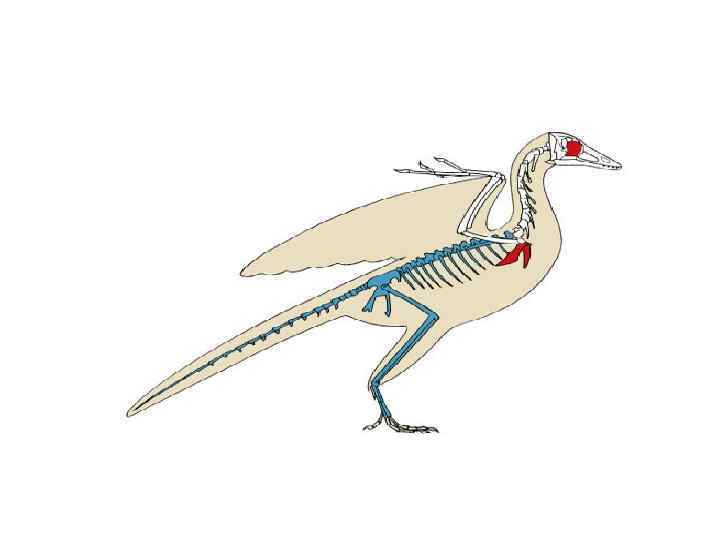
. Вероятные предки – какая-то из групп текодонтов.") • Всередине триаса появились Птерозавры (Pterosauria). Вероятные предки – какая-то из групп текодонтов. Несущий элемент крыла – сильно удлиненный четвертый палец. Летательная перепонка образовывалась за счет складок кожи на боках тела. В скелете имелась грудина с килем, пневматизм костей, срастание костей черепа, крупные глаза. У примитивных форм на челюстях имелись зубы, у эволюционно продвинутых форм зубы редуцированы и челюсти покрывал роговой чехол. Вероятно были теплокровны, тело части видов было покрыто волосоподобными образованиями. Существовали птерозавры до конца мелового периода. рамфоринх
• Всередине триаса появились Птерозавры (Pterosauria). Вероятные предки – какая-то из групп текодонтов. Несущий элемент крыла – сильно удлиненный четвертый палец. Летательная перепонка образовывалась за счет складок кожи на боках тела. В скелете имелась грудина с килем, пневматизм костей, срастание костей черепа, крупные глаза. У примитивных форм на челюстях имелись зубы, у эволюционно продвинутых форм зубы редуцированы и челюсти покрывал роговой чехол. Вероятно были теплокровны, тело части видов было покрыто волосоподобными образованиями. Существовали птерозавры до конца мелового периода. рамфоринх
 Галлодактилус. Фобетор.
Галлодактилус. Фобетор.

 • В конце триаса от орнитозухий, передвигавшихся преимущественно на 2 ногах обособились еще 2 группы – ящеротазовые динозавры (Saurischia) и птицетазовые динозавры (Ornithischia). Обе группы существовали до позднего мела. • Ящеротазовые динозавры имели закрытый таз (у большинства терапод таз становится открытым), четырехкамерное сердце, диафрагму. Передвижение изначально было двуногим, но позднее многие представители вторично перешли к четвероногому передвижению. Имелись как хищные, так и растительноядные формы. Отряд подразделяется на Терапод (Theropoda) и Зауропод (Sauropoda). К группе терапод относятся хищные двуногие динозавры. Многие представители имели признаки, характерные для птиц – перья, строение пояса передних конечностей, открытый таз, преобразование передней конечности в подобие крыла. Большинство вероятнее всего были теплокровными. К зауроподам относятся крупные растительноядные динозавры с длинной шеей, хвостом и большим бочкообразным телом, передвигавшиеся на 4 -х ногах.
• В конце триаса от орнитозухий, передвигавшихся преимущественно на 2 ногах обособились еще 2 группы – ящеротазовые динозавры (Saurischia) и птицетазовые динозавры (Ornithischia). Обе группы существовали до позднего мела. • Ящеротазовые динозавры имели закрытый таз (у большинства терапод таз становится открытым), четырехкамерное сердце, диафрагму. Передвижение изначально было двуногим, но позднее многие представители вторично перешли к четвероногому передвижению. Имелись как хищные, так и растительноядные формы. Отряд подразделяется на Терапод (Theropoda) и Зауропод (Sauropoda). К группе терапод относятся хищные двуногие динозавры. Многие представители имели признаки, характерные для птиц – перья, строение пояса передних конечностей, открытый таз, преобразование передней конечности в подобие крыла. Большинство вероятнее всего были теплокровными. К зауроподам относятся крупные растительноядные динозавры с длинной шеей, хвостом и большим бочкообразным телом, передвигавшиеся на 4 -х ногах.
 Аллозавр Авимим Брахиозавр Микрораптор
Аллозавр Авимим Брахиозавр Микрораптор
 Тероподный динозавр Caudipterix
Тероподный динозавр Caudipterix
 Пигостиль у тероподных динозавров Овираптор Nomyngia. Из Barsbold et al. , 2000 Теризинозавр Beipiaosaurus. Из Xu et al. , 2003.
Пигостиль у тероподных динозавров Овираптор Nomyngia. Из Barsbold et al. , 2000 Теризинозавр Beipiaosaurus. Из Xu et al. , 2003.
 • Птицетазовые динозавры имели открытый таз, напоминающий таз птиц. Ранние представители были двуногими, позже птицетазовые повторно освоили хождение на четырёх конечностях. В ходе эволюции у поздних представителей отряда концевые фаланги пальцев стали более плоскими и приобрели вид копыт. У многих развились приспособления для защиты от хищников: броня, рога, шипы. Были травоядными (за исключением ранних всеядных видов). Зубы имели плоские жевательные поверхности.
• Птицетазовые динозавры имели открытый таз, напоминающий таз птиц. Ранние представители были двуногими, позже птицетазовые повторно освоили хождение на четырёх конечностях. В ходе эволюции у поздних представителей отряда концевые фаланги пальцев стали более плоскими и приобрели вид копыт. У многих развились приспособления для защиты от хищников: броня, рога, шипы. Были травоядными (за исключением ранних всеядных видов). Зубы имели плоские жевательные поверхности.
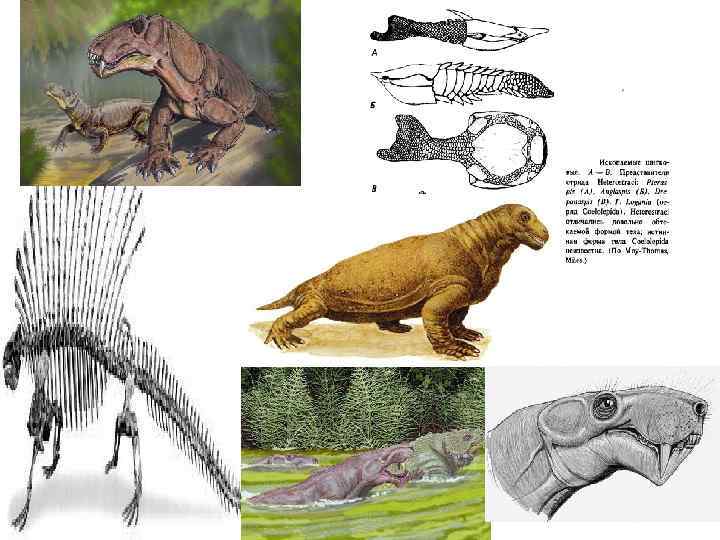
 • основные гипотезы: • 1. Птицы произошли от тероподных") Происхождение и эволюция птиц (Aves) • основные гипотезы: • 1. Птицы произошли от тероподных динозавров группы манираптор или же от целурозавров в юре • 2. Птицы произошли от мелких текодонтов псевдозухий в триасе
Происхождение и эволюция птиц (Aves) • основные гипотезы: • 1. Птицы произошли от тероподных динозавров группы манираптор или же от целурозавров в юре • 2. Птицы произошли от мелких текодонтов псевдозухий в триасе
 и ноги (внизу) в раннем эмбриогенезе у веерохвостых птиц. В") Развитие пальцев крыла (вверху) и ноги (внизу) в раннем эмбриогенезе у веерохвостых птиц. В крыле у птиц остаются II, III и IV пальцы III IV II III II I Из A. Feduccia et al. , 2005. IV
Развитие пальцев крыла (вверху) и ноги (внизу) в раннем эмбриогенезе у веерохвостых птиц. В крыле у птиц остаются II, III и IV пальцы III IV II III II I Из A. Feduccia et al. , 2005. IV
 Наиболее значимые ископаемые формы: Археоптерикс. 8 скелетов данного животного было обнаружено в позднеюрских отложениях Баварии. С самого начала появления археоптерикса на научной арене отмечалось, что строение его скелета является по существу рептильным. В основном наличие оперения заставляло относить его к птицам. С этим, в частности, связано длительное хранение в коллекциях в качестве рептилий некоторых его экземпляров, при которых не сохранилось отпечатков перьев или они были плохо видны. Энанциорнисы. 20 лет назад была открыта совершенно новая группа птиц, названная энанциорнисами. Они найдены в меловых отложениях на всех континентах, кроме Антарктиды. Строение многих костей энанциорнисов может быть выведено из строения костей археоптерикса. Существует некоторый ряд продвинутых или прогрессивных признаков, указывающих на прямые родственные связи энанциорнисов и археоптерикса. Но ящерохвостые птицы почти не имеют общих продвинутых признаков с большинством остальных вымерших и современных птиц и отличаются от них множеством тонких деталей строения. Поэтому большинство остальных птиц относится к другой эволюционной линии.
Наиболее значимые ископаемые формы: Археоптерикс. 8 скелетов данного животного было обнаружено в позднеюрских отложениях Баварии. С самого начала появления археоптерикса на научной арене отмечалось, что строение его скелета является по существу рептильным. В основном наличие оперения заставляло относить его к птицам. С этим, в частности, связано длительное хранение в коллекциях в качестве рептилий некоторых его экземпляров, при которых не сохранилось отпечатков перьев или они были плохо видны. Энанциорнисы. 20 лет назад была открыта совершенно новая группа птиц, названная энанциорнисами. Они найдены в меловых отложениях на всех континентах, кроме Антарктиды. Строение многих костей энанциорнисов может быть выведено из строения костей археоптерикса. Существует некоторый ряд продвинутых или прогрессивных признаков, указывающих на прямые родственные связи энанциорнисов и археоптерикса. Но ящерохвостые птицы почти не имеют общих продвинутых признаков с большинством остальных вымерших и современных птиц и отличаются от них множеством тонких деталей строения. Поэтому большинство остальных птиц относится к другой эволюционной линии.
 • КОНФУЦИУСОРНИС. Обитал на территории Китая в течение раннего мела, примерно 120 млн. лет до нашей эры. Как у современных птиц, клюв конфуциусорниса не имел зубов. Пальцы крыльев имели когти. Обитал у водоемов, скорее всего был способен к полету. Грудина не имела киля. Имелись брюшные ребра. Обитал у водоемов, скорее всего был способен к планирующему полету. • ПРОТОАВИС. Был описан около 10 лет назад из верхнего триаса штата Техас, в США. Строение скелета протоависа имеет удивительные прогрессивные особенности. В первую очередь это – обширная и тонкостенная мозговая коробка, гетероцельное строение шейных позвонков, удлиненные коракоид и лопатка, сочленяющиеся через ямку на коракоиде и выступ на лопатке, слияние концов 3 -й и 4 -й метакарпалий в кисти, глубокие ямки для почек с внутренней стороны подвздошных костей, мыщелок под малую берцовую кость на дистальном эпифизе бедра и другие. Эти прогрессивные признаки характеризуют состояние близкое к таковому у веерохвостых птиц - настоящих птиц. В целом протоавис имел строение скелета несравненно более продвинутое по направлению к веерохвостым птицам, по сравнению с археоптериксом. Скорее всего, по большому набору продвинутых признаков, протоавис должен быть помещен в основание ствола веерохвостых птиц.
• КОНФУЦИУСОРНИС. Обитал на территории Китая в течение раннего мела, примерно 120 млн. лет до нашей эры. Как у современных птиц, клюв конфуциусорниса не имел зубов. Пальцы крыльев имели когти. Обитал у водоемов, скорее всего был способен к полету. Грудина не имела киля. Имелись брюшные ребра. Обитал у водоемов, скорее всего был способен к планирующему полету. • ПРОТОАВИС. Был описан около 10 лет назад из верхнего триаса штата Техас, в США. Строение скелета протоависа имеет удивительные прогрессивные особенности. В первую очередь это – обширная и тонкостенная мозговая коробка, гетероцельное строение шейных позвонков, удлиненные коракоид и лопатка, сочленяющиеся через ямку на коракоиде и выступ на лопатке, слияние концов 3 -й и 4 -й метакарпалий в кисти, глубокие ямки для почек с внутренней стороны подвздошных костей, мыщелок под малую берцовую кость на дистальном эпифизе бедра и другие. Эти прогрессивные признаки характеризуют состояние близкое к таковому у веерохвостых птиц - настоящих птиц. В целом протоавис имел строение скелета несравненно более продвинутое по направлению к веерохвостым птицам, по сравнению с археоптериксом. Скорее всего, по большому набору продвинутых признаков, протоавис должен быть помещен в основание ствола веерохвостых птиц.
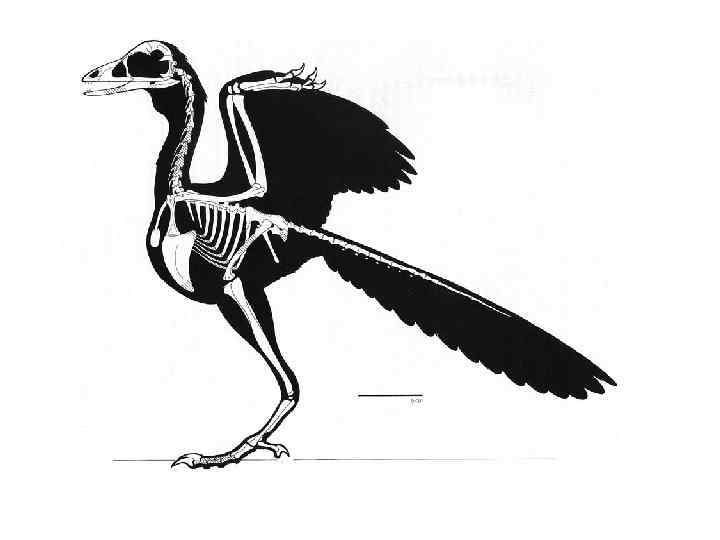
 Археоптерикс
Археоптерикс
 Скелет археоптерикса и современной птицы
Скелет археоптерикса и современной птицы
 Энанциорнисы Nanantius Sinornis Iberomesornis
Энанциорнисы Nanantius Sinornis Iberomesornis
 мозг энанциорниса
мозг энанциорниса
 КОНФУЦИУСОРНИС
КОНФУЦИУСОРНИС
 Протоавис
Протоавис
 Mononykus – птица или тераподный динозавр?
Mononykus – птица или тераподный динозавр?
 Гипотезы появления у птиц полета. • Гипотеза № 1. Сверху – вниз. Птицы произошли от древесных лазающих рептилий, полет появился как планирующий, затем постепенно стал активным. Минусы гипотезы – для лазания и полета используются совершенно разные группы мышц. • Гипотеза № 2. Снизу – вверх. Предками птиц были бегающие животные, которые использовали крылоподобные передние конечности для коротких перелетов во время быстрого бега. Плюсы гипотезы – наличие большого числа оперенных терапод, которые были бегающими животными и имели оперение передних конечностей, напоминающее маховые перья.
Гипотезы появления у птиц полета. • Гипотеза № 1. Сверху – вниз. Птицы произошли от древесных лазающих рептилий, полет появился как планирующий, затем постепенно стал активным. Минусы гипотезы – для лазания и полета используются совершенно разные группы мышц. • Гипотеза № 2. Снизу – вверх. Предками птиц были бегающие животные, которые использовали крылоподобные передние конечности для коротких перелетов во время быстрого бега. Плюсы гипотезы – наличие большого числа оперенных терапод, которые были бегающими животными и имели оперение передних конечностей, напоминающее маховые перья.
 Наземная гипотеза происхождения полета птиц по Chiappe, 2006
Наземная гипотеза происхождения полета птиц по Chiappe, 2006
 Древесная гипотеза происхождения полета птиц по Chatterjee, 1997
Древесная гипотеза происхождения полета птиц по Chatterjee, 1997
") Другие гипотезы происхождения полета Dial, 2003 - гипотеза атакующего проависа (Garner, Taylor, Thomas, 1999) - околоводное (Лопарев, 1996) и водное происхождение полета (Савельев, 2005) Peters, 2002
Другие гипотезы происхождения полета Dial, 2003 - гипотеза атакующего проависа (Garner, Taylor, Thomas, 1999) - околоводное (Лопарев, 1996) и водное происхождение полета (Савельев, 2005) Peters, 2002
 КОМПРОМИССНАЯ ГИПОТЕЗА И ЭВОЛЮЦИОННОМОРФОЛОГИЧЕСКИЙ ПОДХОД Е. Н. Курочкиным разработана компромиссная гипотеза происхождения полета птиц. Она сочетает элементы наземной и древесной гипотез и включает новые подходы: - бипедализм – первичная ключевая адаптация; - анизодактильная лапа – универсальная структура, обеспечившая надежную опору на земле и на ветвях; - анизодактильная лапа также обеспечила возможность редукции длинного хвоста; - длинные крыловые пальцы с когтями у проависов предназначались не для цепляния на стволах, а для цепляния за ветви при поддержании равновесия.
КОМПРОМИССНАЯ ГИПОТЕЗА И ЭВОЛЮЦИОННОМОРФОЛОГИЧЕСКИЙ ПОДХОД Е. Н. Курочкиным разработана компромиссная гипотеза происхождения полета птиц. Она сочетает элементы наземной и древесной гипотез и включает новые подходы: - бипедализм – первичная ключевая адаптация; - анизодактильная лапа – универсальная структура, обеспечившая надежную опору на земле и на ветвях; - анизодактильная лапа также обеспечила возможность редукции длинного хвоста; - длинные крыловые пальцы с когтями у проависов предназначались не для цепляния на стволах, а для цепляния за ветви при поддержании равновесия.
 БИПЕДАЛИЗМ – движение только на задних конечностях Бипедализм освободил передние конечности от функции опоры и дал возможность развиться прыжковой локомоции на земле и на деревьях
БИПЕДАЛИЗМ – движение только на задних конечностях Бипедализм освободил передние конечности от функции опоры и дал возможность развиться прыжковой локомоции на земле и на деревьях
 АНИЗОДАКТИЛЬНАЯ СТОПА – три пальца вперед, один назад Анизодактильная лапа – исходный генерализованный тип стопы у птиц, появилась у предков веерохвостых птиц на самых ранних этапах их эволюции. Обеспечивает надежную опору на земле и на любых насестах, позволяя надежно их обхватывать и передвигаться прыжками по ветвям.
АНИЗОДАКТИЛЬНАЯ СТОПА – три пальца вперед, один назад Анизодактильная лапа – исходный генерализованный тип стопы у птиц, появилась у предков веерохвостых птиц на самых ранних этапах их эволюции. Обеспечивает надежную опору на земле и на любых насестах, позволяя надежно их обхватывать и передвигаться прыжками по ветвям.
 ИЗ ВЕРХНЕТРИАСОВЫХ ОТЛОЖЕНИЙ СЕВ. АМЕРИКИ Стопа протоависа Реконструкция профиля") ПРОТОАВИС (Protoavis texensis Chatterjee, 1991) ИЗ ВЕРХНЕТРИАСОВЫХ ОТЛОЖЕНИЙ СЕВ. АМЕРИКИ Стопа протоависа Реконструкция профиля протоависа (из Chatterjee, 1999)
ПРОТОАВИС (Protoavis texensis Chatterjee, 1991) ИЗ ВЕРХНЕТРИАСОВЫХ ОТЛОЖЕНИЙ СЕВ. АМЕРИКИ Стопа протоависа Реконструкция профиля протоависа (из Chatterjee, 1999)
 Анизодактильная лапа дала возможность предкам птиц крепко держаться на ветвях при вспрыгивании на деревья
Анизодактильная лапа дала возможность предкам птиц крепко держаться на ветвях при вспрыгивании на деревья
 При надежном удержании на ветвях передние конечности сохраняли свободу машущих движений для балансирования и для формирования машущего крыла при спуске путем парашютирования, а не планирования
При надежном удержании на ветвях передние конечности сохраняли свободу машущих движений для балансирования и для формирования машущего крыла при спуске путем парашютирования, а не планирования
 Надежное удерживание на ветвях и сдвиг центра тяжести при дальнейшей эволюции крыла привели к полной редукции длинного хвоста в ряду настоящих веерохвостых птиц
Надежное удерживание на ветвях и сдвиг центра тяжести при дальнейшей эволюции крыла привели к полной редукции длинного хвоста в ряду настоящих веерохвостых птиц
 • На основе новых палеонтологических находок и их сравнительного изучения возникла 3 -я гипотеза происхождения птиц, объединяющая 1 -ю и 2 -ю: • Эволюция птиц происходила 2 -мя независимыми линиями – ящерохвостые птицы произощли от оперенных терапод, к этой же линии относятся археоптериксы и их потомки – энанциорнисы. Появились ящерохвостые птицы в юре, а к концу мелового периода вымерли. Веерохвостые птицы ведут свою родословную от псевдозухий и появились намного раньше ящерохвостых, еще в позднем триасе. Предковой формой веерохвостых является протоавис. Экологическая радиация веерохвостых началась в начале мелового периода.
• На основе новых палеонтологических находок и их сравнительного изучения возникла 3 -я гипотеза происхождения птиц, объединяющая 1 -ю и 2 -ю: • Эволюция птиц происходила 2 -мя независимыми линиями – ящерохвостые птицы произощли от оперенных терапод, к этой же линии относятся археоптериксы и их потомки – энанциорнисы. Появились ящерохвостые птицы в юре, а к концу мелового периода вымерли. Веерохвостые птицы ведут свою родословную от псевдозухий и появились намного раньше ящерохвостых, еще в позднем триасе. Предковой формой веерохвостых является протоавис. Экологическая радиация веерохвостых началась в начале мелового периода.
 Ранние веерохвостые птицы • Гесперорнис. Известны из мела США, Чили и Великобритании. Череп палеогнатический, крылья сильно редуцированы, задние конечности хорошо развиты и приспособлены к плавательным движениям, грудина лишена киля. Некоторые гесперорнисообразные имели зубы в альвеолах. Длина до 2 м. Вели плавающий образ жизни; не летали; по суше, вероятно, передвигались с трудом. По-видимому, гесперорнисы вели такой же образ жизни, как и современные галапагосские бескрылые бакланы. • Ихтиорнис. Известны из верхнего мела Северной Америки. Размером примерно с голубя. Имели грудину с хорошо развитым килем. Челюсти имели зубы сидящие в альвеолах. Скелет крыла типичен для птиц. Питались видимо рыбой, по образу жизни скорее всего были сходны с современными чайками. • Gansus. Известен из мелового периода западного Китая. Был похож на современную гагару, на лапах имелись перепонки. Имелись зубы. Вел видимо водоплавающий образ жизни. • Вегавис. Обнаружен на острове Вега в Антарктике. Жил в начале мела. Возможно ранний представитель гусеобразных.
Ранние веерохвостые птицы • Гесперорнис. Известны из мела США, Чили и Великобритании. Череп палеогнатический, крылья сильно редуцированы, задние конечности хорошо развиты и приспособлены к плавательным движениям, грудина лишена киля. Некоторые гесперорнисообразные имели зубы в альвеолах. Длина до 2 м. Вели плавающий образ жизни; не летали; по суше, вероятно, передвигались с трудом. По-видимому, гесперорнисы вели такой же образ жизни, как и современные галапагосские бескрылые бакланы. • Ихтиорнис. Известны из верхнего мела Северной Америки. Размером примерно с голубя. Имели грудину с хорошо развитым килем. Челюсти имели зубы сидящие в альвеолах. Скелет крыла типичен для птиц. Питались видимо рыбой, по образу жизни скорее всего были сходны с современными чайками. • Gansus. Известен из мелового периода западного Китая. Был похож на современную гагару, на лапах имелись перепонки. Имелись зубы. Вел видимо водоплавающий образ жизни. • Вегавис. Обнаружен на острове Вега в Антарктике. Жил в начале мела. Возможно ранний представитель гусеобразных.
 Гесперорнис
Гесперорнис
 Ихтиорнис
Ихтиорнис
 Вегавис Gansus
Вегавис Gansus
 • После вымирания динозавров птицы заняли экологическую нишу крупных хищников. В третичном периоде известно много видов крупных нелетающих хищных птиц. Позднее эти формы были вытеснены млекопитающими. Phororhacos Titanis walleri
• После вымирания динозавров птицы заняли экологическую нишу крупных хищников. В третичном периоде известно много видов крупных нелетающих хищных птиц. Позднее эти формы были вытеснены млекопитающими. Phororhacos Titanis walleri
 отделилась от общего ствола очень") • Последняя ветвь пресмыкающихся – подкласс Зверообразные (Synapsida) отделилась от общего ствола очень рано (или возникла независимо? ), сохранив множество амфибийных черт. Известны синапсиды с позднего карбона. Для синапсид характерен череп синапсидного типа, кожа была богата железами. Наиболее примитиным среди синапсид является отряд пеликозавров. Сохраняли типично рептильный внешний облик (за исключением того, что не имели достаточно развитых чешуйных покровов). Позвонки амфицельные. Известны с конца карбона до середины перми. Потомками пеликозавров являлись Терапсиды или зверозубые (Therapsida). Для них характерны признаки, роднящие их с млекопитающими – гетеродонтные зубы, богатая железами кожа, наличие на коже волосоподобных образований, строение конечностей. Появились терапсиды в начале перми и большинство представителей вымерло в конце перми – начале триаса. Исключение составили цинодонты — от них произошли млекопитающие. Цинодонты дожили до раннего мелового периода.
• Последняя ветвь пресмыкающихся – подкласс Зверообразные (Synapsida) отделилась от общего ствола очень рано (или возникла независимо? ), сохранив множество амфибийных черт. Известны синапсиды с позднего карбона. Для синапсид характерен череп синапсидного типа, кожа была богата железами. Наиболее примитиным среди синапсид является отряд пеликозавров. Сохраняли типично рептильный внешний облик (за исключением того, что не имели достаточно развитых чешуйных покровов). Позвонки амфицельные. Известны с конца карбона до середины перми. Потомками пеликозавров являлись Терапсиды или зверозубые (Therapsida). Для них характерны признаки, роднящие их с млекопитающими – гетеродонтные зубы, богатая железами кожа, наличие на коже волосоподобных образований, строение конечностей. Появились терапсиды в начале перми и большинство представителей вымерло в конце перми – начале триаса. Исключение составили цинодонты — от них произошли млекопитающие. Цинодонты дожили до раннего мелового периода.
 Офиакодон Cotylorhynchus Эдафозавр диметродон
Офиакодон Cotylorhynchus Эдафозавр диметродон
: Дейноцефалы (Dinocephalia) Горгонопии (Gorgonopia) иностранцевия титанофонеус мосхопс") Диноморфы (Dinomorpha): Дейноцефалы (Dinocephalia) Горгонопии (Gorgonopia) иностранцевия титанофонеус мосхопс
Диноморфы (Dinomorpha): Дейноцефалы (Dinocephalia) Горгонопии (Gorgonopia) иностранцевия титанофонеус мосхопс
: Дицинодонты Lystrosaurus Тероцефалы (Therocephalia)") Аномодонты (Anomodontia): Дицинодонты Lystrosaurus Тероцефалы (Therocephalia)
Аномодонты (Anomodontia): Дицинодонты Lystrosaurus Тероцефалы (Therocephalia)
") Цинодонты (Cynodontia)
Цинодонты (Cynodontia)
 отряда Зверозубых. Так, цинодонты") • Наиболее вероятные предки млекопитающих - подотряд Цинодонты (Cynodontia) отряда Зверозубых. Так, цинодонты имели вторичное костное небо, которое несло заклыковые зубы (гомологичные коренным зубам млекопитающих), а также многочисленные щечные железы, гомологичные слюнным железам млекопитающих. В плечевом поясе наблюдалась редукция коракоида, а в тазовом - разрастание подвзодшных костей; наличие же поясничного отдела позвоночника, видимо, означало появление диафрагмы. На костях челюстей некоторых представителей обнаружены следы прикрепления губных мышц. Вероятно, тогда же появились и молочные железы - видоизмененные потовые; вероятно, первоначальной их функцией было не столько "выкармливание", сколько "выпаивание" детенышей и снабжение их солями. Тогда же, вероятно, сложилось разделение сердца на четыре камеры, ставшее предпосылкой теплокровности. Наиболее эволюционно продвинутых цинодонтов относят к группе маммалиаморф. • Первые млекопитающие появляются в отложениях верхнего триаса. Первой появившейся группой млекопитающих были Триконодонты (Triconodonta). Известны из верхнего триаса — верхнего мела Сев. Америки и Евразии. Размеры мелкие реже средние. В строении черепа сохранились черты, свойственные зверообразным пресмыкающимся цинодонтам. На коренных зубах три бугорка, расположенных вдоль оси зуба (отсюда название— трёхконусозубые). Зубная формула примитивная. Были хищниками, питались в зависимости от размера насекомыми или мелкими позвоночными. Вымерли к концу мелового периода не оставив потомков.
• Наиболее вероятные предки млекопитающих - подотряд Цинодонты (Cynodontia) отряда Зверозубых. Так, цинодонты имели вторичное костное небо, которое несло заклыковые зубы (гомологичные коренным зубам млекопитающих), а также многочисленные щечные железы, гомологичные слюнным железам млекопитающих. В плечевом поясе наблюдалась редукция коракоида, а в тазовом - разрастание подвзодшных костей; наличие же поясничного отдела позвоночника, видимо, означало появление диафрагмы. На костях челюстей некоторых представителей обнаружены следы прикрепления губных мышц. Вероятно, тогда же появились и молочные железы - видоизмененные потовые; вероятно, первоначальной их функцией было не столько "выкармливание", сколько "выпаивание" детенышей и снабжение их солями. Тогда же, вероятно, сложилось разделение сердца на четыре камеры, ставшее предпосылкой теплокровности. Наиболее эволюционно продвинутых цинодонтов относят к группе маммалиаморф. • Первые млекопитающие появляются в отложениях верхнего триаса. Первой появившейся группой млекопитающих были Триконодонты (Triconodonta). Известны из верхнего триаса — верхнего мела Сев. Америки и Евразии. Размеры мелкие реже средние. В строении черепа сохранились черты, свойственные зверообразным пресмыкающимся цинодонтам. На коренных зубах три бугорка, расположенных вдоль оси зуба (отсюда название— трёхконусозубые). Зубная формула примитивная. Были хищниками, питались в зависимости от размера насекомыми или мелкими позвоночными. Вымерли к концу мелового периода не оставив потомков.
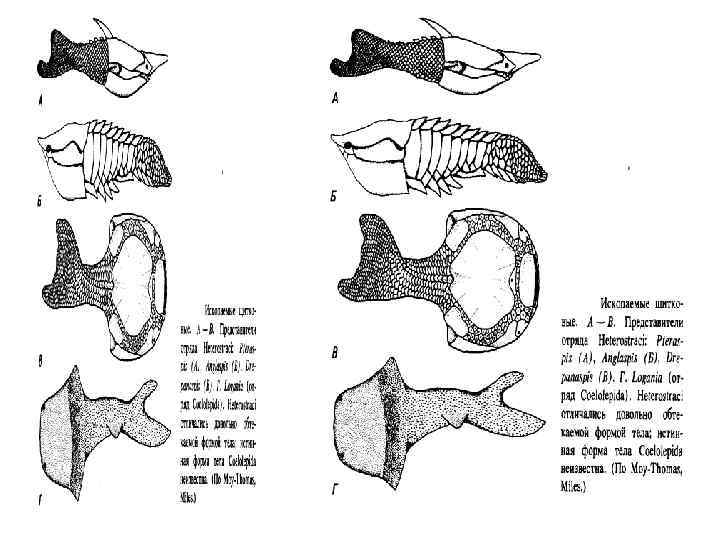
. Систематическое") Castorocauda lutrasimilis. Найден в отложениях юрского периода (около 164 млн лет назад ). Систематическое положение не ясно, скорее всего был маммалиаморфом.
Castorocauda lutrasimilis. Найден в отложениях юрского периода (около 164 млн лет назад ). Систематическое положение не ясно, скорее всего был маммалиаморфом.
 Gobiconodon Репеномамус
Gobiconodon Репеномамус
 , получившие своё") • В начале юры появляются многобугорчатые (инфракласс Аллотерии, отряд Multituberculata) , получившие своё название в связи с наличием на коренных зубах многочисленных бугорков. Это была специализированная группа животных с очень сильно развитыми резцами и без клыков. Размеры имели мелкие. Судя по строению таза, рождали мелких детенышей, наподобие сумчатых. У некоторых обнаружено присутствие сумчатых костей. Многобугорчатые представляли специфических растительноядных зверей, и их нельзя считать предками последующих групп млекопитающих. Вымерли многобугорчатые в начале палеогена. • В юрском периоде появились пантотерии (инфракласс Трёхбугорчатые (Trituberculata). Известны с нижней юры до верхнего мела Евразии, Сев. Америки и Африки. Размером с мышь или крысу. Найдены лишь остатки челюстей и зубов. Судя по строению зубов — насекомоядные; возможно, питались также птицами, яйцами, плодами. В мелу от Пантотериев произошли сумчатые и плацентарные.
• В начале юры появляются многобугорчатые (инфракласс Аллотерии, отряд Multituberculata) , получившие своё название в связи с наличием на коренных зубах многочисленных бугорков. Это была специализированная группа животных с очень сильно развитыми резцами и без клыков. Размеры имели мелкие. Судя по строению таза, рождали мелких детенышей, наподобие сумчатых. У некоторых обнаружено присутствие сумчатых костей. Многобугорчатые представляли специфических растительноядных зверей, и их нельзя считать предками последующих групп млекопитающих. Вымерли многобугорчатые в начале палеогена. • В юрском периоде появились пантотерии (инфракласс Трёхбугорчатые (Trituberculata). Известны с нижней юры до верхнего мела Евразии, Сев. Америки и Африки. Размером с мышь или крысу. Найдены лишь остатки челюстей и зубов. Судя по строению зубов — насекомоядные; возможно, питались также птицами, яйцами, плодами. В мелу от Пантотериев произошли сумчатые и плацентарные.
 Многобугорчатые, Ptilodus Meniscoessus
Многобугорчатые, Ptilodus Meniscoessus

 • Однопроходные в ископаемом состоянии известны с начала мелового периода. Ископаемые формы близки к утконосам, однако их челюсти несут хорошо развитые зубы. Ископаемые находки известны из Авсралии и Южной Америки. Размеры были довольно крупными – до метра в длину. Предполагается, что вели полуводный образ жизни, питаясь водными беспозвоночными. Происхождение однопроходных не ясно, существуют 2 гипотезы, по одной из них однопроходные произошли независимо от остальных млекопитающих от каких-то маммалиаморф, по другой – являются потомками пантотериев. Steropodon
• Однопроходные в ископаемом состоянии известны с начала мелового периода. Ископаемые формы близки к утконосам, однако их челюсти несут хорошо развитые зубы. Ископаемые находки известны из Авсралии и Южной Америки. Размеры были довольно крупными – до метра в длину. Предполагается, что вели полуводный образ жизни, питаясь водными беспозвоночными. Происхождение однопроходных не ясно, существуют 2 гипотезы, по одной из них однопроходные произошли независимо от остальных млекопитающих от каких-то маммалиаморф, по другой – являются потомками пантотериев. Steropodon
 • Инфракласс сумчатых обособился в меловом периоде. Предполагаемое место возникновения – Северная Америка, по другим данным – Азия. Наиболее древние формы сумчатых были видимо близки к отряду Микробиотерииды (Microbiotheria). В Южной Америке во время отсутствия Панамского перешейка сумчатые имели очень высокое разнообразие и занимали различные экологические ниши. После соединения с Северной Америкой подавляющее большинство их вымерло. Из Южной Америки через Антарктиду сумчатые проникли в Австралию. Thylacosmilus atrox Diprotodon
• Инфракласс сумчатых обособился в меловом периоде. Предполагаемое место возникновения – Северная Америка, по другим данным – Азия. Наиболее древние формы сумчатых были видимо близки к отряду Микробиотерииды (Microbiotheria). В Южной Америке во время отсутствия Панамского перешейка сумчатые имели очень высокое разнообразие и занимали различные экологические ниши. После соединения с Северной Америкой подавляющее большинство их вымерло. Из Южной Америки через Антарктиду сумчатые проникли в Австралию. Thylacosmilus atrox Diprotodon
 • Уже в конце мелового периода сформировались отряды насекомоядных и тенрекообразных. Млекопитающие, близкие к предковым формам этих отрядов, дали начало отряды приматов и, возможно, отряду рукокрылых. Первые приматы известны из верхнего мела Северной Америки. • В конце мелового периода появляются первые кондиляртры – предки копытных. В Палеогене группа достигает значительного расцвета. Размеры различны — от лисицы до крупной лошади. Головной мозг очень маленький. Бугорчатые коренные зубы свидетельствуют о плохом приспособлении к перетиранию растительной пищи, на что указывают и сильно развитые клыки. Ноги короткие, пятипалые, оканчивающиеся копытами. Некоторые кондиляртры были травоядными, но имелись так же всеядные и хищные формы. • Хищная группа кондиляртров – арктоционы – дала начало группе мезонихий – среднего размера или крупных хищных млекопитающих, близких к предкам парнокопытных и китообразных. Мезонихии на пальцах имели не когти, а копыта.
• Уже в конце мелового периода сформировались отряды насекомоядных и тенрекообразных. Млекопитающие, близкие к предковым формам этих отрядов, дали начало отряды приматов и, возможно, отряду рукокрылых. Первые приматы известны из верхнего мела Северной Америки. • В конце мелового периода появляются первые кондиляртры – предки копытных. В Палеогене группа достигает значительного расцвета. Размеры различны — от лисицы до крупной лошади. Головной мозг очень маленький. Бугорчатые коренные зубы свидетельствуют о плохом приспособлении к перетиранию растительной пищи, на что указывают и сильно развитые клыки. Ноги короткие, пятипалые, оканчивающиеся копытами. Некоторые кондиляртры были травоядными, но имелись так же всеядные и хищные формы. • Хищная группа кондиляртров – арктоционы – дала начало группе мезонихий – среднего размера или крупных хищных млекопитающих, близких к предкам парнокопытных и китообразных. Мезонихии на пальцах имели не когти, а копыта.
 Эндрюсарх Арктоцион Фенакодус
Эндрюсарх Арктоцион Фенакодус
 Ископаемые непарнокопытные Кадуркодон Халикотерий и анцилотерий Индрикотерий Пропалеотерий
Ископаемые непарнокопытные Кадуркодон Халикотерий и анцилотерий Индрикотерий Пропалеотерий
 Ископаемые парнокопытные Энтелодон диакодексис Доркатерий Сиватерий
Ископаемые парнокопытные Энтелодон диакодексис Доркатерий Сиватерий
 • Китообразные произошли от мелких примитивных парнокопытных в первой половине палеогена. Одним из самых примитивных китообразных был пакицетис – околоводное млекопитающее размером с собаку, жившее 50 млн. лет назад на морских побережьях Индийского субматерика. Более специализированным к водному образу жизни был амбулоцетус, но и он способен был ходить по суше. Родоцетус был приспособлен в основном к жизни в море, но на берег выходить был способен. Полностью водными животными были базилозавр и дорудон, однако у низ имелись рудименты задних конечностей с 3 пальцами. Жили эти киты на мелководье и плавали хуже, чем современные виды. После перехода китообразных к жизни в открытом океане наметилось их разделение на усатых и зубатых китов. У ранних усатых китов еще сохраняются некрупные зубы и одновременно появляется китовый ус, возникающий, как разрастание роговых гребней неба.
• Китообразные произошли от мелких примитивных парнокопытных в первой половине палеогена. Одним из самых примитивных китообразных был пакицетис – околоводное млекопитающее размером с собаку, жившее 50 млн. лет назад на морских побережьях Индийского субматерика. Более специализированным к водному образу жизни был амбулоцетус, но и он способен был ходить по суше. Родоцетус был приспособлен в основном к жизни в море, но на берег выходить был способен. Полностью водными животными были базилозавр и дорудон, однако у низ имелись рудименты задних конечностей с 3 пальцами. Жили эти киты на мелководье и плавали хуже, чем современные виды. После перехода китообразных к жизни в открытом океане наметилось их разделение на усатых и зубатых китов. У ранних усатых китов еще сохраняются некрупные зубы и одновременно появляется китовый ус, возникающий, как разрастание роговых гребней неба.
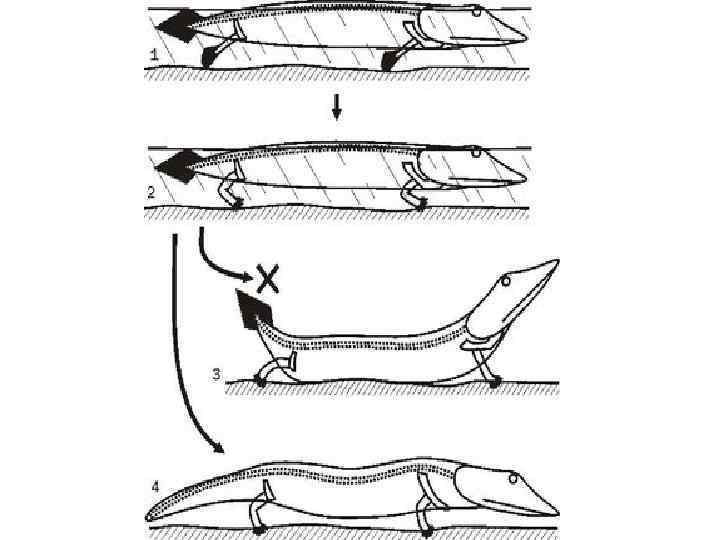
 Амбулоцетус Indohyus Aetiocetus Дорудон Пакицет Родоцетус Базилозавр
Амбулоцетус Indohyus Aetiocetus Дорудон Пакицет Родоцетус Базилозавр
 • В самом начале палеогена появляются хищные. Их предковые формы дают две ветви – собственно хищных и креодонтов. • Креодонты отличаются малым объемом мозга и строением челюстей – они менее подвижны. Однако в палеогене креодонты образуют большое число видов. В неогене креодонты полностью вымирают, не оставив потомков. • Хищные появляются практически одновременно с креодонтами. Исходная группа – миациды. Вскоре появляется предковая линия подотряда кошкоподобных – виверравиды. Миациды же дают начало подотряду псоподобных. • В конце палеогена некоторые хищные переходят к водному образу жизни и образуют семейства тюленей, ушастых тюленей и моржовых. Предковые формы тюленей близки к семейству куньих, а ушастых тюленей и моржовых – к медвежьим. Предковой формой настоящих тюленей считается Потамотерий – представитель семейства куньих.
• В самом начале палеогена появляются хищные. Их предковые формы дают две ветви – собственно хищных и креодонтов. • Креодонты отличаются малым объемом мозга и строением челюстей – они менее подвижны. Однако в палеогене креодонты образуют большое число видов. В неогене креодонты полностью вымирают, не оставив потомков. • Хищные появляются практически одновременно с креодонтами. Исходная группа – миациды. Вскоре появляется предковая линия подотряда кошкоподобных – виверравиды. Миациды же дают начало подотряду псоподобных. • В конце палеогена некоторые хищные переходят к водному образу жизни и образуют семейства тюленей, ушастых тюленей и моржовых. Предковые формы тюленей близки к семейству куньих, а ушастых тюленей и моржовых – к медвежьим. Предковой формой настоящих тюленей считается Потамотерий – представитель семейства куньих.
 миасис Гиенодон Саркастодон Потамотерий
миасис Гиенодон Саркастодон Потамотерий