Проблемы видов-двойников УУде.ppt
- Количество слайдов: 51



Проблемы видов-двойников и методы их решения l «Биоразнообразие: глобальные и региональные процессы» Картавцева Ирина Васильевна, Биолого-почвенный институт ДВО РАН, Улан-Удэ, Россия, 16 -21 сентября 2013 г.
. n морфологический критерий (выявляющий")
ТИПЫ ВИДОВ Выделение видов происходит на основании двух принципов (критериев). n морфологический критерий (выявляющий различия между видами) и n репродуктивной изоляции (оценивающий степень их генетической обособленности). Процедура описания новых видов зачастую сопряжена с определенными сложностями, связанных как с неоднозначным соответствием критериев вида другу, так и с постепенностью и незавершенностью процесса видообразования. В зависимости от того, какого рода сложности возникали при выделении видов и каким образом они были решены, выделяют так называемые "типы видов".

Монотипический вид n Часто никаких сложностей при описании новых видов не возникает. Такие виды обычно обладают обширным неразорванным ареалом, на протяжении которого географическая изменчивость выражена слабо.

Политипический вид. Нередко с помощью морфологического критерия выделяется целая группа близкородственных форм, обитающих, как правило, в сильно расчлененной местности (в горах, либо на островах). Каждая из этих форм имеет свой, обычно довольно ограниченный ареал. Если между сравниваемыми формами существует географический контакт, то возможно применение критерия репродуктивной изоляции: если гибриды не возникают, либо относительно редки, этим формам придается статус самостоятельных видов; в противном случае описывают разные подвиды одного вида. Вид, включающий в себя несколько подвидов называется политипическим. n

Когда же анализируемые формы географически изолированы, оценка их статуса довольно субъективна и происходит только на основании морфологического критерия: если различия между ними "существенны", то перед нами разные виды, если нет – подвиды. Не всегда в группе близкородственых форм удается однозначно определить статус каждой формы. Иногда группа популяций замыкается в кольцо, охватывающее горный массив или земной шар. В таком случае может оказаться, что "хорошие" (обитающие совместно и не гибридизирующие) виды связаны друг с другом цепью подвидов. l

"Комплекс форм“ больших белоголовых чаек l Ареал комплекса больших белоголовых чаек охватывает широким кольцом северное полушарие земного шара. В пределах комплекса выделяют до 15 различных форм, различающихся по окраске спины и крыльев, цветом ног и окологлазничного кольца. Их таксономический статус которых невозможно установить, так как обитающие совместно и негибридизирующие виды (серебристая чайка и чайка-клуша) оказываются связанными друг с другом цепью гибридизирующих подвидов. Кольцевые перекрывания являются наиболее убедительными доказательствами географического видообразования.

Полиморфный вид l Адаптивный полиморфизм богомола У богомола существует зеленая и бурая морфы. Первая плохо заметна на зеленых частях растений, вторая на ветках деревьев и сухой траве. В экспериментах по пересаживанию богомолов на фон не соответствующий их окраске удалось показать, что полиморфизм в данном случае мог возникнуть и поддерживается благодаря естественному отбору: зеленая и бурая окраска богомолов является защитой от хищников и позволяет этим насекомым меньше конкурировать друг с другом.

Гибридогенный полиморфизм испанской каменки Каменка-плешанка • Испанская каменка белогорлая морфа Испанская каменка черногорлая морфа У самцов испанской каменки существуют белогорлая и черногорлая морфы. Характер соотношения этих морф в разных частях ареала позволяет предположить, что черногорлая морфа образовалась в результате гибридизации с близкородственным видом - каменкой-плешанкой.
 и плешанки (внизу):")
Испанская каменка и каменка-плешанка Диагностические признаки окраски самок испанской каменки (вверху) и плешанки (внизу): а - черноватый цвет, б -бурый, в - рыжий ЗООЛОГИЧЕСКИЙ ЖУРНАЛ 1992, том 71, вып. 1 © 1992 г. В. И. ГРАБОВСКИЙ, Е. Н. ПАНОВ, А. С. РУБЦОВ ФЕНОТИПИЧЕСКИЙ СОСТАВ И УСПЕХ РАЗМНОЖЕНИЯ В ГИБРИДНОЙ ПОПУЛЯЦИИ ПЛЕШАНКИ OENANTHE PLESCHANKA И ИСПАНСКОЙ КАМЕНКИ О. HISPANICA В ВОСТОЧНОМ АЗЕРБАЙДЖАНЕ

ПРОБЛЕМНЫЕ ВИДЫ • Обыкновенная и белошапочная овсянки - два хорошо различающихся вида. Область их совместного обитания простирается на 2, 5 тыс. км. с запада на восток (от Уральских гор до Байкала) и на 2 тыс. км. с севера на юг. При этом между ними существует регулярная гибридизация: на разных участках зоны перекрывания ареалов гибриды составляют от 3 до 50% численности популяции.

"Полувиды" • Видообразование - длительный процесс, и поэтому можно столкнуться с такими формами, статус которых нельзя оценить объективно. Они еще не являются самостоятельными видами, поскольку гибридизируют в природе, но это уже и не подвиды, так как морфологические различия между ними весьма существенны. Такие формы называются "пограничными случаями", "проблемными видами" или "полувидами". Формально им приписываются бинарные латинские названия, как у "нормальных" видов, и в таксономических списках они помещаются рядом друг с другом. "Полувиды" встречаются не так уж и редко, и мы сами зачастую не подозреваем, что окружающие нас виды являются типичными примерами "пограничных случаев".

Виды-двойники виды, обитающие совместно и не скрещивающиеся друг с другом, но очень слабо отличающиеся морфологически. Сложность различения таких видов связана со сложностью выделения или неудобстве использования их диагностических признаков - ведь сами видыдвойники прекрасно разбираются в собственной "таксономии". Чаще видыдвойники встречаются среди групп животных, использующих для поиска полового партнера запах (насекомые, грызуны) и реже - у тех, которые пользуются зрительной и акустической сигнализацией (птицы).
 используют принятые основные критерии: •")
Виды-двойники • В систематике для определения вида (биологическая концепция) используют принятые основные критерии: • Морфологический (включая цитогенетический), • Экологический, • Физиологический (включая генетический, гибридизационный), • Географический и • этологический. Тщательное исследование морфологических признаков позволило выявить виды-двойники – виды, сходные морфологически (сестринские «sibling» , скрытые «cryptic» , двойники «twin» ), но в природе надежно разделенные репродуктивной или географической изоляцией.

виды-двойники • Термин «виды-двойники» введен Эрнстом Майром в 1942 с определением: «пары или группы видов, которые морфологически не различимы или различимы с трудом» , для синиц рода Parus [P. 151]. Позже видыдвойники были обнаружены Добжанским у плодовых мушек рода Drosophila, а определение для этого термина было уточнено – «морфологически сходные или идентичные популяции, которые репродуктивно изолированы • “Sibling species are species that are very similar in appearance, in behavior and in other characteristics, but they are reproductively isolated”. » [ P. 34].
 и курганчиковая (Mus spicilegus) мыши. • Домовая и курганчиковая мыши внешне")
Домовая (Mus musculus) и курганчиковая (Mus spicilegus) мыши. • Домовая и курганчиковая мыши внешне неразличимы. Тем не менее их образ жизни сильно различается, и в природе они не скрещиваются. Домовая мышь строго территориальна, курганчиковая мышь живет поселениями, формируя "курганчики" с многочисленными норами. Домовая мышь терпима к человеку и часто проникает в дома, курганчиковая мышь не может жить по соседству с человеком.

Strophurus intermedius ящерицы • Strophurus williamsi
 и сосновик (Loxia pytyopsittacus) • Эти два вида клестов являются")
Клесты еловик (Loxia curvirostra) и сосновик (Loxia pytyopsittacus) • Эти два вида клестов являются одним из немногочисленных примеров видов-двойников среди птиц. Обитая совместно на значительной территории, охватывающую Северную Европу и Скандинавский полуостров, эти виды не скрещиваются друг с другом. Морфологические различия между ними, незначительные и весьма ненадежные, выражаются в размерах клюва: у сосновика он несколько толще, чем у еловика.
. Черногрудый воробей (Paster hispaniolensis). В Средней Азии домовый")
• Домовый воробей (Paster domesticus). Черногрудый воробей (Paster hispaniolensis). В Средней Азии домовый воробей обитает совместно с другим близкородственным видом - черногрудым воробьем, от которого хорошо отличается по окраске. Гибридизация между ними в этом районе отсутствует. Их систематический статус как самостоятельных видов не вызывал бы сомнений, если бы не существовала вторая зона контакта в Европе. Италию населяет особая форма воробьев, возникшая в результате гибридизации домового и испанского. При этом в Испании, где домовый и испанский воробьи также обитают совместно, гибриды встречаются редко.
 n 3.")
Методы: генетический 1. Хромосомный n 2. Аллозимный (подвижность белков в электрофорезном геле) n 3. Молекулярно-генетический n

Хромосомный

Молекулярно-генетический ДНК баркодинг начинается с публикации в 2003 группой Пола Хеберта (Paul Hebert’s research group at the University of Guelph) статьи n «Биологическая идентификация с помощью ДНК штрихкодирования» "Biological identifications through DNA barcodes". n Они предложили новую систему идентификации видов и открыли использование короткого участка ДНК стандартного района генома мт ДНК n
 n CCTATACCTAATCTTCGGAGCATGAGC GGGCATGGTAGGC. . . COI мт ДНК")
Арктическая славка (Phylloscopus borealis) n CCTATACCTAATCTTCGGAGCATGAGC GGGCATGGTAGGC. . . COI мт ДНК

mt. DNA sequencing in the Barcode of Life project Building the Barcode Library n 2, 012, 391 specimens 309, 803 barcode clusters/species Target for 2015: 5, 000 specimens 500, 000 species
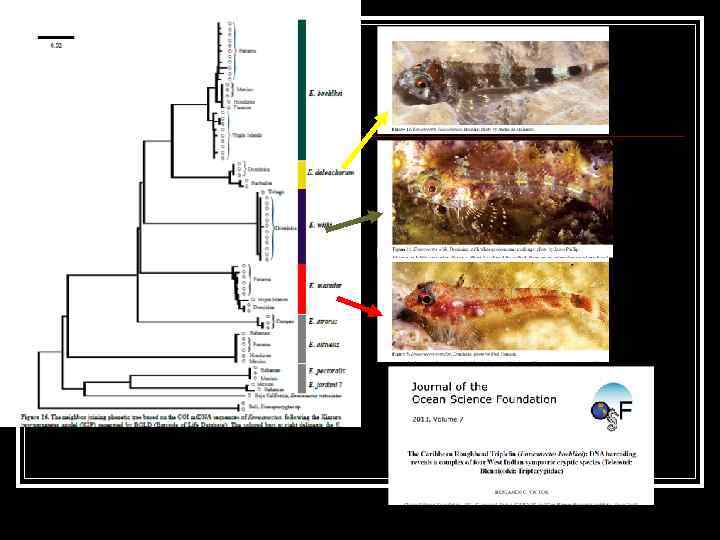

Международная конференция по баркодингу, Автралия, Аделаида, декабрь 2011 г.

p-дистанции в различных таксономических группах Kartavtsev, Lee, 2006 Cyt-b and Co-1 генв мт ДНК. Группы: 1. внутривидовые, особи одного вида; 2. 2. подвиды и виды двойники, 3. 3. виды одного рода; 4. 4. роды одного семейства

Генетические дистанции в разных группах животных

n Виды-двойники хомячков группы «barabensis»

Cricetulus barabensis sensu lato

Распространение хромосомных форм Распр “barabensis” 2 n=20, NF=38 “pseudogriseus” 2 n=24, NF=38 “griseus” 2 n=22, NF=38 “sokolovi” 2 n=20, NF=40

Внешний вид хомячков из Poplavskaja et al. , 2011
")
Морфологический анализ (Лебедев Лисовский, 2008)

2 n=24 Хромосомный анализ 2 n=22 2 n=20 B. Kral et al COMPARISON OF KARYOTYPES, G-BANDS AND NORS IN THREE CRICETULUS SPP. (CRICETIDAE, RODENTIA) FOLIA ZOOLOGICA - 33(1): 85 -96 (1984) n 2 n=20
")
n. Молекулярный анализ (Neumann et al. , 2006)
. 2")
Распределение гаплотипов цитохрома b хомячков рода Cricetulus (по Лебедев и др. , 2007). 2 n=22 2 n=24 2 n=20 Cyt b, различие между «кариоморфами» 24% Divergence rate ~8% per My 2 n=20
 0, 15 0, 32 “barabensis” –")
Генетическое разнообразие хомячков в Забайкалье 0, 04 (!) 0, 15 0, 32 “barabensis” – 2 n=20 0, 2 0, 45 0, 42 0, 37 0, 52 0, 35 0, 68 0, 37 0, 61 0, 57 0, 58 0, 75 0, 36 “pseudogriseus” – 2 n=24 0, 69 0, 39 0, 22 0, 21
 r. Selenga Селенга “barabensis” “pseudogriseus” r.")
Зона потенциального контакта в Бурятии (данные группы Лебедева) r. Selenga Селенга “barabensis” “pseudogriseus” r. Чикой Chikoy
 2 n n ( is” s e rab a “b ”")
Река Чикой (Бурятия) 2 n n ( is” s e rab a “b ” us e ris g do u se “p 4) 2 n= (2 ) 0 =2
 самец Cr. b. barabensis (2 n=20) x самка Cr. b.")
ГИБРИДНЫЙ КАРИОТИП (2 n=21) самец Cr. b. barabensis (2 n=20) x самка Cr. b. griseus (2 n=22) (по данным московской группы исследователей)
")
Точки хромосомного исследования (из Вакурин и др. , 2014 Зоол. Журн, в печати)
 Cr. barabensis (2 n=20)")
Окрашивание на структурный гетерохроматин (из Вакурин и др. , 2014) Cr. barabensis (2 n=20) Cr. griseus (2 n=22)
 Cr. griseus (2")
Схема различий по количеству и локализации С-блоков Cr. barabensis (2 n=20) Cr. griseus (2 n=22)

Дифференциальная С-окраска 2 -й пары аутосом барабинских хомячков C. barabensis из различных географических регионов НВ – Новосибирская область, БР – Республика Бурятия, ЗБ 1 – Забайкальский край, ЗБ 2 – Сохондинский заповедник, АМ – Амурская область, ЕВ – Еврейская Автономная область, ПР – Приморский край.

Полевки группы «maximowiczii» M. fortis 2 n=52 M. sachalinensis 2 n=50 M. maximowiczii 2 n=36 -44 M. gromovi 2 n=44 M. evoronenesis 2 n=38 -42 M. mujanensis 2 n=38

Хромосомы и перестройки видов-двойников M. mujanensis 2 n=38 - инверсии M. evoronenesis 2 n=38 -40 - слияния 2 n=40 2 n=38

M. m. ungurensis 2 n=39 -44 Nf=52 -62 M. m. maximowiczii 2 n=40 -42 Nf=56 -58, 60 2 n=38 -43 Nf=54 -57 2 n=36 -41 Nf=51 -58 2 n=40 -43 Nf=54 -56, 58

Microtus maximowiczii gromovi M. m. ungurensis M. maximowiczii ungurensis Из Мейер и др. , 1996. M. m. ungurensis M. m. gromovi 2 n=44, Nf=60
 cit b ДНК")
Генетическое исследование видов рода Microtus Аллозимный (Фрисман и др. , 2008) cit b ДНК (Sheremetyeva et al. , 2008)

Microtus gromovi stat. nov. gromovi Sheremetyeva , Kartavtseva, Voyta , Kryukov and Haring Morphometric analysis of intraspecific variation in Microtus maximowiczii (Rodentia, Cricetidae) in relation to chromosomal differentiation with reinstatement of Microtus gromovi Vorontsov, Journal of Zoological Systematics and Evolutionary Research 2009 Vol. 47, 1, P. 42 -48. gromovi

Выводы Таким образом, применение комплексного анализа методов, как правило, позволяет решить проблему видов-двойников. n Для каждого отдельно взятого вида используют свой набор методов, т. к. универсальных методов нет n

n Мы просто похожи
Проблемы видов-двойников УУде.ppt