

Митоз и мейоз.ppt
- Количество слайдов: 23
 Презентации к лекциям на тему: ЭУКАРИОТИЧЕСКАЯ ХРОМОСОМА, Митоз, Мейоз Гришанин А. К.
Презентации к лекциям на тему: ЭУКАРИОТИЧЕСКАЯ ХРОМОСОМА, Митоз, Мейоз Гришанин А. К.
 равноплечие; 2) явственно") С. Г. Навашин, в 1911 г. выделил 3 типа хромосом: 1) равноплечие; 2) явственно неравноплечие; 3) крючковидные. В 1924 г. Г. А. Левитский ввел понятие кариотип –
С. Г. Навашин, в 1911 г. выделил 3 типа хромосом: 1) равноплечие; 2) явственно неравноплечие; 3) крючковидные. В 1924 г. Г. А. Левитский ввел понятие кариотип –

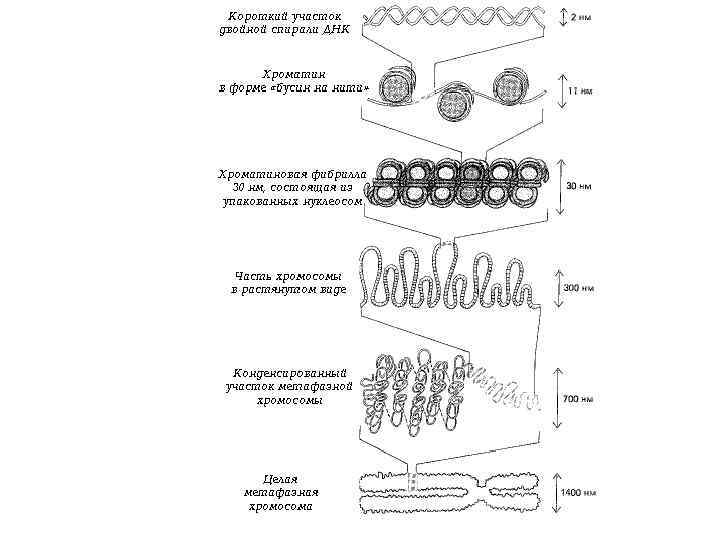
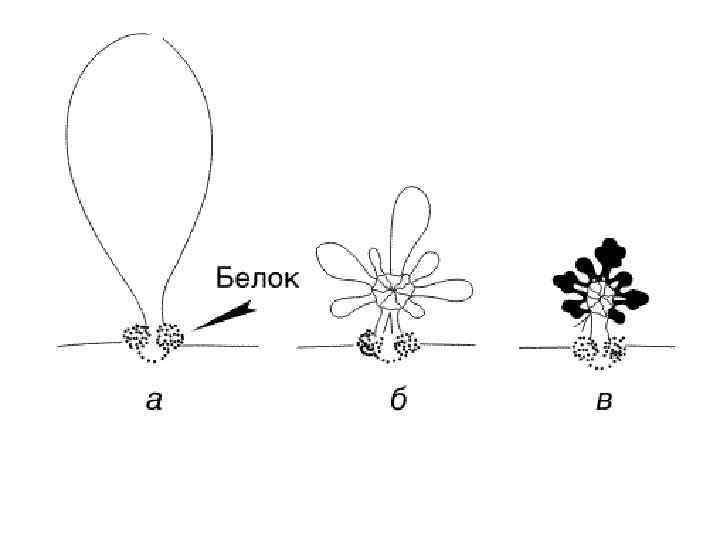
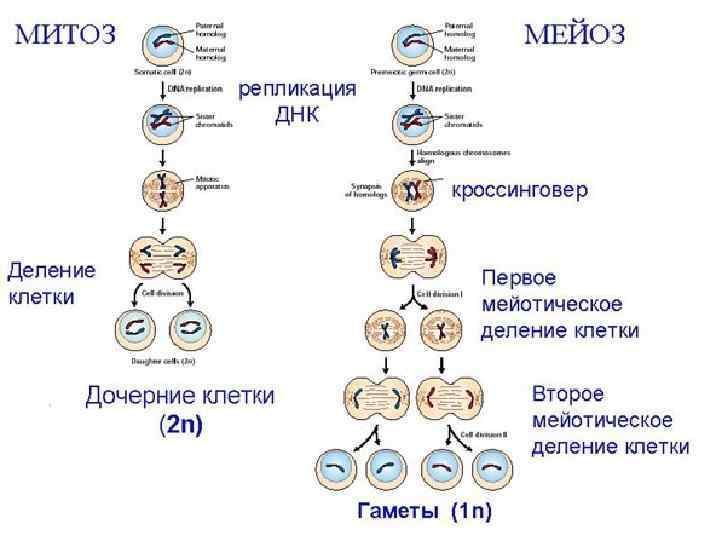
 Процесс деления, при котором исходно диплоидная клетка дает две дочерние, также диплоидные, клетки, называется митозом (Mitos лат. – нить). Основное свойство митоза- точное расхождение сестринских хроматид в дочерние ядра. На обеспечение этого процесса направлены все основные события митоза: 1)обособление и последующее расхождение сестринских центросом до полюсов веретена, в направление которых будут расходиться хромосомы в анафазе; 2) реорганизация сети интерфазных микротрубочек в полярные веретена, исходящие от центросомы; 3) реорганизация диффузного интерфазного хроматина в удобную для транспортировки структуру – конденсированную митотическую хромосому
Процесс деления, при котором исходно диплоидная клетка дает две дочерние, также диплоидные, клетки, называется митозом (Mitos лат. – нить). Основное свойство митоза- точное расхождение сестринских хроматид в дочерние ядра. На обеспечение этого процесса направлены все основные события митоза: 1)обособление и последующее расхождение сестринских центросом до полюсов веретена, в направление которых будут расходиться хромосомы в анафазе; 2) реорганизация сети интерфазных микротрубочек в полярные веретена, исходящие от центросомы; 3) реорганизация диффузного интерфазного хроматина в удобную для транспортировки структуру – конденсированную митотическую хромосому
![Цитологи разделяют клеточное деление на 5 фаз: интерфазу(G 1, [G 0], S, G 2),](https://present5.com/presentation/-88870855_371531903/image-7.jpg "Цитологи разделяют клеточное деление на 5 фаз: интерфазу(G 1, [G 0], S, G 2),") Цитологи разделяют клеточное деление на 5 фаз: интерфазу(G 1, [G 0], S, G 2), профазу, метафазу, анафазу и телофазу. Интерфазное ядро имеет гомогенную структуру, является транскрипционно-активным, в интерфазном ядре происходит удвоение центриолей и репликация хромосом. «Машина» синтеза ДНК выбирает ядро с нереплицированными хромосомами. 1)происходит активация MPF (Maturation Promoting Factor), инициируется репликация, другой белковый фактор контролирует однократную репликацию. После репликации каждая хромосома становится как бы удвоенной. Каждая хромосома в этой паре называется хроматидой. До анафазы две сестринские хроматиды сцеплены вместе, благодаря молекулам белка когезина, который сшивает хроматиды. 2) Расходятся центросомы в ранней профазе. В конце профазы начале прометафазы обычно исчезают ядерная оболочка и ядрышко.
Цитологи разделяют клеточное деление на 5 фаз: интерфазу(G 1, [G 0], S, G 2), профазу, метафазу, анафазу и телофазу. Интерфазное ядро имеет гомогенную структуру, является транскрипционно-активным, в интерфазном ядре происходит удвоение центриолей и репликация хромосом. «Машина» синтеза ДНК выбирает ядро с нереплицированными хромосомами. 1)происходит активация MPF (Maturation Promoting Factor), инициируется репликация, другой белковый фактор контролирует однократную репликацию. После репликации каждая хромосома становится как бы удвоенной. Каждая хромосома в этой паре называется хроматидой. До анафазы две сестринские хроматиды сцеплены вместе, благодаря молекулам белка когезина, который сшивает хроматиды. 2) Расходятся центросомы в ранней профазе. В конце профазы начале прометафазы обычно исчезают ядерная оболочка и ядрышко.
 Во время прометафазы формируется веретено деления за счет микротрубочек интерфазной клетки, хромосомы конденсируются") 3) Во время прометафазы формируется веретено деления за счет микротрубочек интерфазной клетки, хромосомы конденсируются и передвигаются в область экватора клетки. Конденсация хромосом идет от центромеры к теломерам. В конденсации хроматина участвует большое количество белков, основной среди которых – конденсин. На хромосоме образуется кинетохор, структура позволяющая полярным нитям веретена деления присоединиться к хромосоме. 4) В метафазе каждая хромосома располагается в экваториальной плоскости таким образом, что центромера каждой хромосомы находится точно в экваториальной плоскости. Во время метафазы – анафазы цитоплазма имеет наименьшую вязкость. 5) Во время анафазы разделяются центромеры хромосом, и сестринские хроматиды, которые уже называются дочерними хромосомами, растягиваются к полюсам клетки. В это время цитоплазма имеет наименьшую вязкость, что значительно облегчает движение хромосом в цитоплазме. Перед анафазой клетка проверяет число и состояние кинетохоров. 6)В телофазе дочерние хромосомы деспирализуются и утрачивают видимую индивидуальность. Образуется ядро, ядерная оболочка, ядрышко и все внутриядерные структуры, затем клетка делится (цитокинез).
3) Во время прометафазы формируется веретено деления за счет микротрубочек интерфазной клетки, хромосомы конденсируются и передвигаются в область экватора клетки. Конденсация хромосом идет от центромеры к теломерам. В конденсации хроматина участвует большое количество белков, основной среди которых – конденсин. На хромосоме образуется кинетохор, структура позволяющая полярным нитям веретена деления присоединиться к хромосоме. 4) В метафазе каждая хромосома располагается в экваториальной плоскости таким образом, что центромера каждой хромосомы находится точно в экваториальной плоскости. Во время метафазы – анафазы цитоплазма имеет наименьшую вязкость. 5) Во время анафазы разделяются центромеры хромосом, и сестринские хроматиды, которые уже называются дочерними хромосомами, растягиваются к полюсам клетки. В это время цитоплазма имеет наименьшую вязкость, что значительно облегчает движение хромосом в цитоплазме. Перед анафазой клетка проверяет число и состояние кинетохоров. 6)В телофазе дочерние хромосомы деспирализуются и утрачивают видимую индивидуальность. Образуется ядро, ядерная оболочка, ядрышко и все внутриядерные структуры, затем клетка делится (цитокинез).
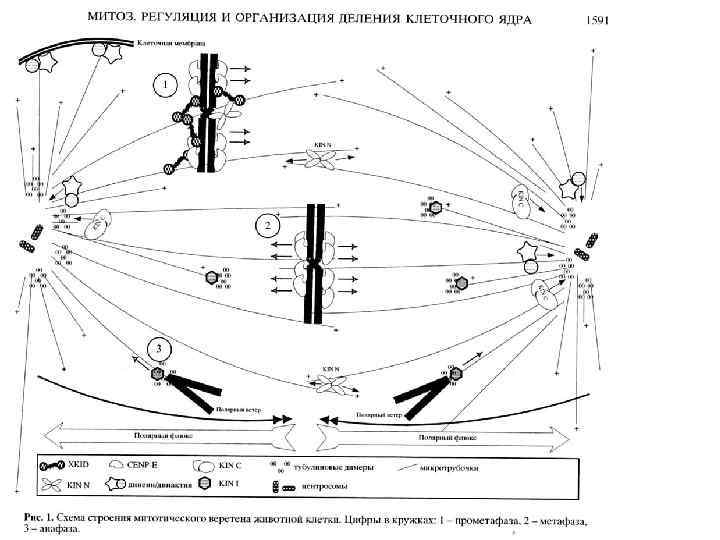
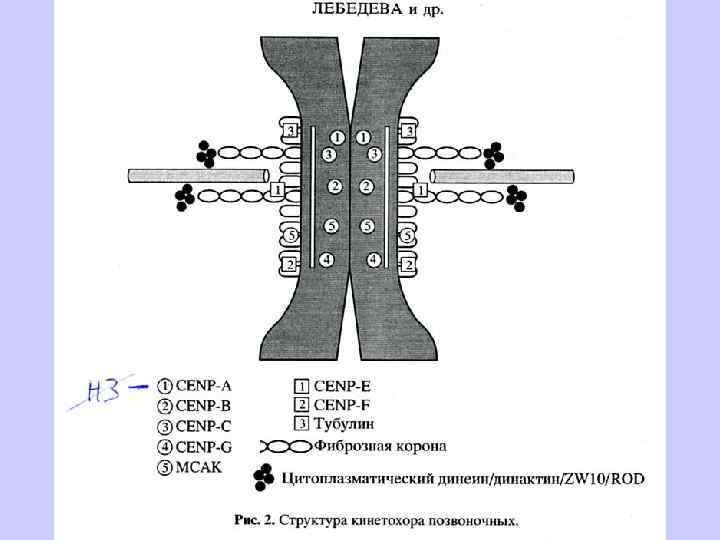
 Кинетохор формируется на центромере только во время митоза и на время митоза. Кинетохор представляет собой трехслойную дископодобную структуру из белков и ДНК. Петли хроматина выходят на поверхность кинетохора, связывая его внутренние и внешние слои. Кинетохор позвоночных состоит из множества функциональных единиц. У дрожжей одна, у позвоночных от 10 до 45. У человека -30. Функции кинетохора: 1) присоединение к микротрубочкам, 2) транспортировка хромосом, 3) задержка митотического деления в случае повреждения веретена или несцепления микротрубочек с кинетохором. ДНК центромерного хроматина упакована в нуклеосомоподобные частицы, в которых гистон Н 3 замещен гистон-подобным белком CENP-A, который в отличие от Н 3 не фосфорилируется, что препятствует конденсации хроматина.
Кинетохор формируется на центромере только во время митоза и на время митоза. Кинетохор представляет собой трехслойную дископодобную структуру из белков и ДНК. Петли хроматина выходят на поверхность кинетохора, связывая его внутренние и внешние слои. Кинетохор позвоночных состоит из множества функциональных единиц. У дрожжей одна, у позвоночных от 10 до 45. У человека -30. Функции кинетохора: 1) присоединение к микротрубочкам, 2) транспортировка хромосом, 3) задержка митотического деления в случае повреждения веретена или несцепления микротрубочек с кинетохором. ДНК центромерного хроматина упакована в нуклеосомоподобные частицы, в которых гистон Н 3 замещен гистон-подобным белком CENP-A, который в отличие от Н 3 не фосфорилируется, что препятствует конденсации хроматина.
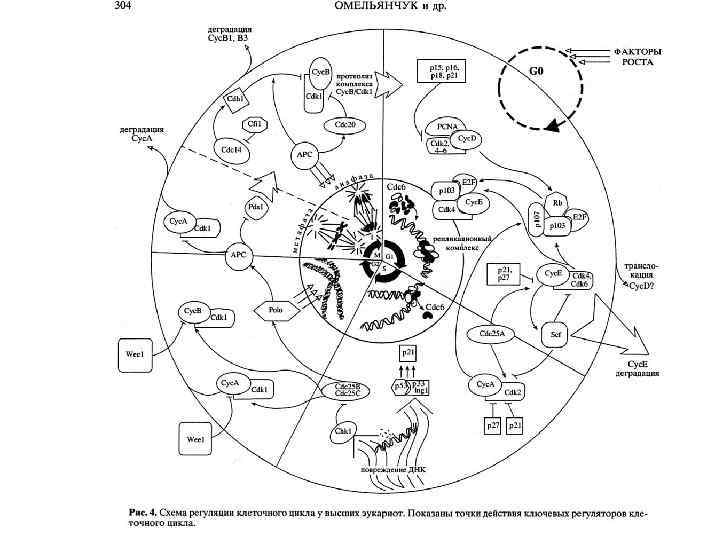
 Регуляция событий митоза организована так, чтобы гарантировать точную сегрегацию хромосом. Циклины белкирегуляторы клеточного цикла обеспечивают программы: пролиферации, дифференцировки, апоптоза. Подсчитано, что примерно около 100 генов эукариот вовлечены в организацию митоза, что составляет 1/6 всех генов клеточного цикла. Нарушение точной сегрегации контролируется системой Checpoints, приводящей к задержке митоза. Задержка может быть обратимой, если за счет репарационных или компенсаторных механизмов произошло восстановление процесса митоза. В случае аномальной сегрегации хромосом происходят необратимые нарушения, и дочерние клетки останавливаются в интерфазе следующего цикла. Если система контроля не срабатывает, а деление клеток не блокируется, то возможна трансформация делящихся клеток в раковые.
Регуляция событий митоза организована так, чтобы гарантировать точную сегрегацию хромосом. Циклины белкирегуляторы клеточного цикла обеспечивают программы: пролиферации, дифференцировки, апоптоза. Подсчитано, что примерно около 100 генов эукариот вовлечены в организацию митоза, что составляет 1/6 всех генов клеточного цикла. Нарушение точной сегрегации контролируется системой Checpoints, приводящей к задержке митоза. Задержка может быть обратимой, если за счет репарационных или компенсаторных механизмов произошло восстановление процесса митоза. В случае аномальной сегрегации хромосом происходят необратимые нарушения, и дочерние клетки останавливаются в интерфазе следующего цикла. Если система контроля не срабатывает, а деление клеток не блокируется, то возможна трансформация делящихся клеток в раковые.
 Ассиметричный митоза - сестринские клетки имеют разные размеры, содержат неравное количество цитоплазмы и имеют разную судьбу. Например яйцеклетки животных при созревании. Амитоз представляет собой прямое деление интерфазного ядра путем перетяжки без образования хромосом. Эндомитоз, способ деления клетки, при котором происходит умножение числа хромосом без деления клетки или деления ядра. Хромосомы репродуцируются в интерфазе, расхождение их происходит внутри ядра без разрушения ядерной оболочки. Эндорепродукция - удвоение одной или нескольких хромосом без преобразований ядра. Митоз с задержкой цитокинеза происходит в результате деления ядра в отсутствие цитокинеза, что приводит к появлению многоядерных клеток вроде синцития. При политении происходит редупликация хромосом без их расхождения. Хромосомы при этом не расходятся и примыкают друг к другу по всей длине.
Ассиметричный митоза - сестринские клетки имеют разные размеры, содержат неравное количество цитоплазмы и имеют разную судьбу. Например яйцеклетки животных при созревании. Амитоз представляет собой прямое деление интерфазного ядра путем перетяжки без образования хромосом. Эндомитоз, способ деления клетки, при котором происходит умножение числа хромосом без деления клетки или деления ядра. Хромосомы репродуцируются в интерфазе, расхождение их происходит внутри ядра без разрушения ядерной оболочки. Эндорепродукция - удвоение одной или нескольких хромосом без преобразований ядра. Митоз с задержкой цитокинеза происходит в результате деления ядра в отсутствие цитокинеза, что приводит к появлению многоядерных клеток вроде синцития. При политении происходит редупликация хромосом без их расхождения. Хромосомы при этом не расходятся и примыкают друг к другу по всей длине.
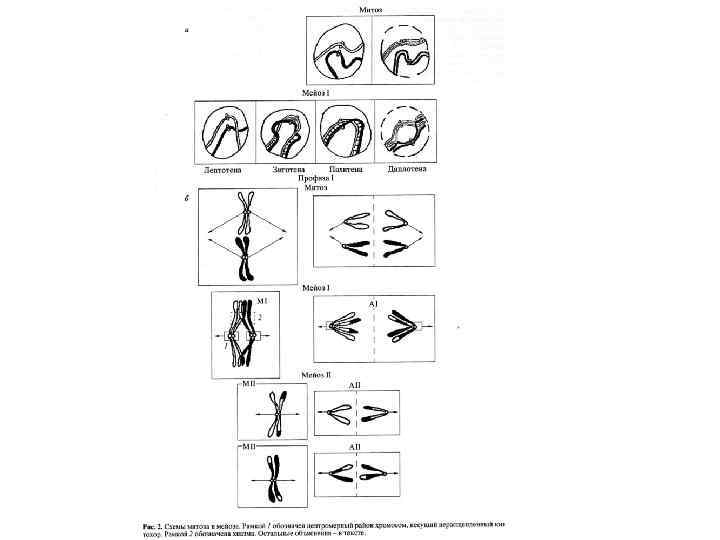
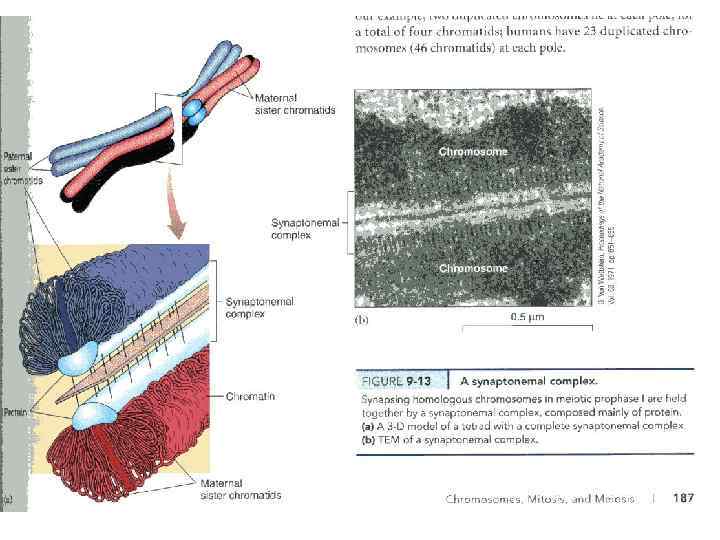
 Характерная особенность мейоза заключается в том, что процесс редупликации ДНК происходит только в интерфазе первого деления мейоза, а второе деление мейоза проходит без синтеза ДНК, поэтому во время 2 -го мейотического деления количество ДНК в клетке уменьшается вдвое. Из исходных клеток с диплоидным набором хромосом возникают гаметы с гаплоидным набором. Из одной диплоидной клетки образуются четыре гаплоидные. Основные этапы мейоза: 1) в премейотической интерфазе и ранней лептотене под действием специфических для мейоза эндонуклеаз возникают двойные разрывы ДНК. Одновременно наблюдается локальное взаимное узнавание отдельных локусов гомологичных хромосом и их попарное соединение. 2) На стадии зиготены происходит тесное сближение гомологичных хромосом по всей их длине – синапсис. Хромосомы состоящие из двух хроматид образуются биваленты, в состав которых входит 4 хроматиды. Начинается формирование синаптонемных комплексов.
Характерная особенность мейоза заключается в том, что процесс редупликации ДНК происходит только в интерфазе первого деления мейоза, а второе деление мейоза проходит без синтеза ДНК, поэтому во время 2 -го мейотического деления количество ДНК в клетке уменьшается вдвое. Из исходных клеток с диплоидным набором хромосом возникают гаметы с гаплоидным набором. Из одной диплоидной клетки образуются четыре гаплоидные. Основные этапы мейоза: 1) в премейотической интерфазе и ранней лептотене под действием специфических для мейоза эндонуклеаз возникают двойные разрывы ДНК. Одновременно наблюдается локальное взаимное узнавание отдельных локусов гомологичных хромосом и их попарное соединение. 2) На стадии зиготены происходит тесное сближение гомологичных хромосом по всей их длине – синапсис. Хромосомы состоящие из двух хроматид образуются биваленты, в состав которых входит 4 хроматиды. Начинается формирование синаптонемных комплексов.
 пахитена: завершение построения СК по всей длине бивалентов), происходят заключительные события рекомбинации -") 3) пахитена: завершение построения СК по всей длине бивалентов), происходят заключительные события рекомбинации - в виде кроссинговера. 4) диплотена начинается с момента расхождения гомологичных хромосом. На стадии диплотены СК поэтапно разрушается, исчезает контакт (когезия) сестринских хроматид во всех локусах кроме хиазм, и хиазмы -результат кроссинговера - становятся видимыми в микроскоп. Число хиазм обычно соответствует количеству рекомбинантных событий в пахитене. 5)Диакинез - дальнейшее укорочение хромосом вследствие компактизации. 6) метафаза-I: гомологичные хромосомы, соединенные хиазмами в виде бивалентов, коориентируются к разным полюсам на экваторе веретена клеточного деления. (7) Во время анафазы-I гомологичные хромосомы расходятся. Полная сегрегация рекомбинировавших рецессивных и доминантных аллелей совершается только в анафазе.
3) пахитена: завершение построения СК по всей длине бивалентов), происходят заключительные события рекомбинации - в виде кроссинговера. 4) диплотена начинается с момента расхождения гомологичных хромосом. На стадии диплотены СК поэтапно разрушается, исчезает контакт (когезия) сестринских хроматид во всех локусах кроме хиазм, и хиазмы -результат кроссинговера - становятся видимыми в микроскоп. Число хиазм обычно соответствует количеству рекомбинантных событий в пахитене. 5)Диакинез - дальнейшее укорочение хромосом вследствие компактизации. 6) метафаза-I: гомологичные хромосомы, соединенные хиазмами в виде бивалентов, коориентируются к разным полюсам на экваторе веретена клеточного деления. (7) Во время анафазы-I гомологичные хромосомы расходятся. Полная сегрегация рекомбинировавших рецессивных и доминантных аллелей совершается только в анафазе.
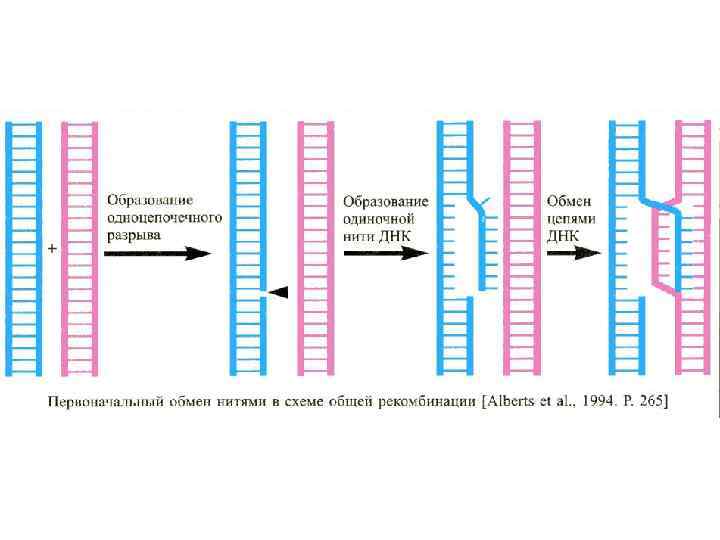
 Рекомбинация между гомологичными хромосомами приводит к интенсивной перетасовке отцовских и материнских генов во время мейоза. Это событие происходит на стадии четырех хроматид у самцов в ходе сперматогенеза, а у самок в ходе оогенеза. В обмене участвуют две из четырех хроматид. Гомологичная рекомбинация происходит между двумя дуплексными молекулами ДНК. Действия генов, управляющие стадиями мейоза, приводят к тому , что: гены синапсиса, гены рекомбинации гомологичных хромосом в профазе-I мейоза и гены, обеспечивающие расхождение хромосом в метафазе – I, обусловливают расхождение в анафазе I не сестринских хроматид (как в митозе), а гомологичных хромосом, каждая из которых остается состоящей из двух сестринских хроматид, которые могут быть кроссоверными.
Рекомбинация между гомологичными хромосомами приводит к интенсивной перетасовке отцовских и материнских генов во время мейоза. Это событие происходит на стадии четырех хроматид у самцов в ходе сперматогенеза, а у самок в ходе оогенеза. В обмене участвуют две из четырех хроматид. Гомологичная рекомбинация происходит между двумя дуплексными молекулами ДНК. Действия генов, управляющие стадиями мейоза, приводят к тому , что: гены синапсиса, гены рекомбинации гомологичных хромосом в профазе-I мейоза и гены, обеспечивающие расхождение хромосом в метафазе – I, обусловливают расхождение в анафазе I не сестринских хроматид (как в митозе), а гомологичных хромосом, каждая из которых остается состоящей из двух сестринских хроматид, которые могут быть кроссоверными.

 • Хромосомная теория наследственности • В 1902 -1904 г. г. Теодор Бовери и Уолтер Сэттон высказали идею о параллелизме между независимым расщеплением признаков при их наследовании и поведением гомологичных хромосом в мейозе, что послужило первым шагом к формированию хромосомной теории наследственности. В 1910 г. Томасом Морганом и его студентом Кальвином Бриджесом в 1916 г. , была показана четкая взаимосвязь между отдельными генами и отдельными хромосомами. Их опыты показывали, что два аллеля находящиеся в разных гомологичных хромосомах расходятся в разные гаметы вместе с гомологичными хромосомами во время мейоза. Те аллельные гены, которые находятся в парных гомологичных хромосомах, попадают в разные гаметы независимо от других аллельных генов локализующихся в других парах гомологичных хромосом и комбинируются вместе с хромосомами, в которых они находятся. Линейное расположение генов в хромосоме также стало понятным после работы Т. Моргана и его сотрудников А. Стертеванта, Г. Меллера, К. Бриджеса и др. , после того как Морган показал, что гены локализованы в хромосомах и расположены в них линейно.
• Хромосомная теория наследственности • В 1902 -1904 г. г. Теодор Бовери и Уолтер Сэттон высказали идею о параллелизме между независимым расщеплением признаков при их наследовании и поведением гомологичных хромосом в мейозе, что послужило первым шагом к формированию хромосомной теории наследственности. В 1910 г. Томасом Морганом и его студентом Кальвином Бриджесом в 1916 г. , была показана четкая взаимосвязь между отдельными генами и отдельными хромосомами. Их опыты показывали, что два аллеля находящиеся в разных гомологичных хромосомах расходятся в разные гаметы вместе с гомологичными хромосомами во время мейоза. Те аллельные гены, которые находятся в парных гомологичных хромосомах, попадают в разные гаметы независимо от других аллельных генов локализующихся в других парах гомологичных хромосом и комбинируются вместе с хромосомами, в которых они находятся. Линейное расположение генов в хромосоме также стало понятным после работы Т. Моргана и его сотрудников А. Стертеванта, Г. Меллера, К. Бриджеса и др. , после того как Морган показал, что гены локализованы в хромосомах и расположены в них линейно.