1МорфолУльтрастрКультивВирусов .ppt
- Количество слайдов: 101



Правила оформления тестового контроля На отдельном листе бумаги написать: Учебная дисциплина: Микробиология, вирусология, иммунология. Номер (или название) темы тестового контроля: Тема 1, или «Морфология и ультраструктура вирусов» . Группа: написать номер Студент: написать фамилию и имя Ответы на вопросы: написать номер вопроса и правильный вариант ответа. Например: 1 D. Под ответами: поставить дату (число, месяц, год) и личную подпись.

Модуль 2. Тестовый контроль по теме № 1: • Морфология и ультраструктура вирусов

1. Вирусы впервые открыты: A. Пастером Л. B. Мечниковым И. C. Кохом Р. D. Ивановским Д. E. Бейеринком М. 2. Первый научный доклад и публикация (1892 г. ) были посвящены вирусам, которые вызывают заболевания: А. Людей B. Животных C. Растений D. Грибов Е. Бактерий

3. Размеры вирусов определяются в нм с помощью: A. Люминисцентной микроскопии B. Электронной микроскопией C. Радиальной иммунодиффузии D. Световой микроскопии E. Фазово-контрастной микроскопии 4. Наиболее крупным вирусом является: A. Вирус полиомиелита B. Вирус натуральной оспы C. Вирус ящура D. Вирус гриппа E. Вирус кори

5. Какие признаки положены в основу современной классификации вирусов? A. Тип нуклеиновой кислоты B. Особенности структуры нуклеиновой кислоты C. Сложность строения и размер вирионов D. Форма капсида (тип симметрии и др. ) E. Круг восприимчивых хозяев 6. К поздним стадиям взаимодействия вируса с клеткой относятся: A. Адсорбция E. Сборка вирионов B. Проникновение F. Выход вирусов C. Раздевание D. Синтез вирусных макромолекул
 D.")
7. Вирион сложноустроенных вирусов содержит: A. Нуклеиновую кислоту B. Капсид C. Суперкапсид (пеплос) D. Ядро E. Митохондрии 8. Типы взаимодействия вирусов с клеткой хозяина: A. Продуктивный B. Абортивный C. Интегративный D. Активный E. Пассивный
 обусловлен наличием в их вирионе фермента:")
9. Специфический процесс репликации генома ретровирусов (онковирусы, ВИЧ) обусловлен наличием в их вирионе фермента: А. Эндонуклеазы D. ДНК - полимеразы В. Нейраминидазы Е. РНК -полимеразы С. Обратной транскриптазы 10. Культивирование (выращивание) вирусов осуществляют на: A. Твердых питательных средах B. Жидких питательных средах C. Лабораторных животных D. Куриных эмбрионах E. Культурах клеток

Тема практического занятия: Вирусология. Морфология и ультраструктура вирусов. Культивирование вирусов. С. И. Похил ©

ВИРУСОЛОГИЯ – НАУКА О МОРФОЛОГИИ, ФИЗИОЛОГИИ, ГЕНЕТИКЕ, ЭКОЛОГИИ И ЭВОЛЮЦИИ ВИРУСОВ. МЕДИЦИНСКАЯ ВИРУСОЛОГИЯ ИЗУЧАЕТ ВИРУСЫ-ПАРАЗИТЫ ЧЕЛОВЕКА, ИХ РОЛЬ В РАЗВИТИИ ЗАБОЛЕВАНИЙ, РАЗРАБАТЫВАЕТ МЕТОДЫ ДИАГНОСТИКИ, СПОСОБЫ ТЕРАПИИ И СПЕЦИФИЧЕСКОЙ ПРОФИЛАКТИКИ ВИРУСНЫХ ЗАБОЛЕВАНИЙ.
 Вирусы открыты ботаником Д. И. Ивановским (18641920 гг. ). Начало")
Virus (лат. – яд) Вирусы открыты ботаником Д. И. Ивановским (18641920 гг. ). Начало науки вирусологии 12 февраля 1892 г. Первая научная работа о вирусах : «О двух болезнях табака» .

Ивановский Д. И. доказал, что возбудитель табачной мозаики проходит сквозь бактериальный фильтр, который задерживал при фильтрации известные бактерии. Профильтрованный экстракт вызывает заболевание у здоровых растений при многочисленных повторных переносах, тоесть - вызывает заболевание и размножается как бактерии. Но, в отличие от бактерий, новый тип инфекционного агента не растет на питательных средах, а может жить и размножаться только в живых клетках.
 с")
Новый тип возбудителя был намного меньше известных бактерий, его не удавалось увидеть (описать) с помощью светового микроскопа и подтвердить «триадай Коха» . Датский ботаник М. Бейеринк в 1898 г. повторил и дополнил эксперименты Д. И. Ивановского и ввел термин – вирус, так как ошибочно считал вирусы не живым началом, а ядовитыми веществами.
 группа доклеточных прокариотических форм микроорганизмов, относятся")
1. Вирусы самостоятельная (отличная от бактерий и грибов) группа доклеточных прокариотических форм микроорганизмов, относятся к отдельному царству живого Vira. 2. Вирусы характеризуются малыми размерами, ввыраженным цитотропизмом, дизъюнктивным (разделенным) типом репликации, облигатным паразитизмом: они не имеют собственного аппарата синтеза биополимеров и энергообеспечения, поэтому не способны к самостоятельному росту и делению, но обладают кардинальными свойствами живого.
 в восприимчивых клетках животных,")
Основные признаки живого у вирусов: 1. Способность к размножению (репродукции) в восприимчивых клетках животных, растений и бактерий. 2. Наследственность (передача признаков потомству). 3. Изменчивость (способность к адаптации для выживания).
 ФОРМА (СТАДИЯ) ВИРУСОВ, ВЫПОЛНЯЮЩАЯ ФУНКЦИЮ ПЕРЕНОСА")
ФОРМЫ СУЩЕСТВОВАНИЯ ВИРУСОВ ВИРИОН ВИРУС ПРОВИРУС ВНЕКЛЕТОЧНАЯ (ПОКОЯЩАЯСЯ) ФОРМА (СТАДИЯ) ВИРУСОВ, ВЫПОЛНЯЮЩАЯ ФУНКЦИЮ ПЕРЕНОСА ГЕНОМА ИЗ ОДНОЙ КЛЕТКИ В ДРУГУЮ РЕПРОДУКТИВНАЯ ФОРМА (ВНУТРИКЛЕТОЧНАЯ СТАДИЯ РАЗВИТИЯ) ФОРМА ИНТЕГРАЦИИ ГЕНОМА ВИРУСА С ГЕНОМОМ КЛЕТКИХОЗЯИНА, РЕПРОДУКЦИЯ ВИРУСА НЕ ПРОИСХОДИТ, НО ОН СОХРАНЯЕТСЯ И МОЖЕТ ВЫЗЫВАТЬ ВИРУСИНДУЦИРУЕМЫЕ ИЗМЕНЕНИЯ В КЛЕТКЕ

Вирусы – облигатные внутриклеточные генетические паразиты. Для репродукции вирусы используют синтетический и энергетический аппарат клетки-мишени : • - рибосомы; • - мембраны; • - митохондрии; • - ферменты.

q. Геном среднего размера вируса в 106 раз меньше размера клетки, однако успешно конкурирует с последним и навязывает зараженной клетке реализацию собственной генетической информации с репликацией (размножением) вируса. q. Когда вирус находится вне клетки, в нём не происходит никаких метаболических процессов (инертен) и только сохраняется его генетическая информация (без ее изменения).

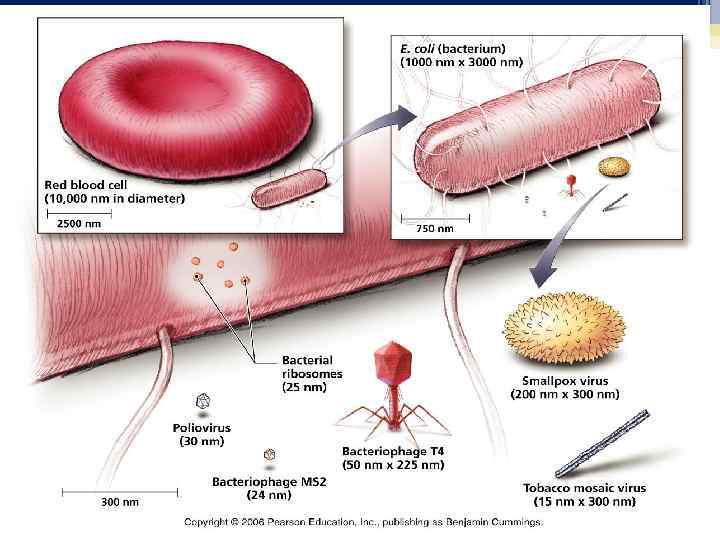
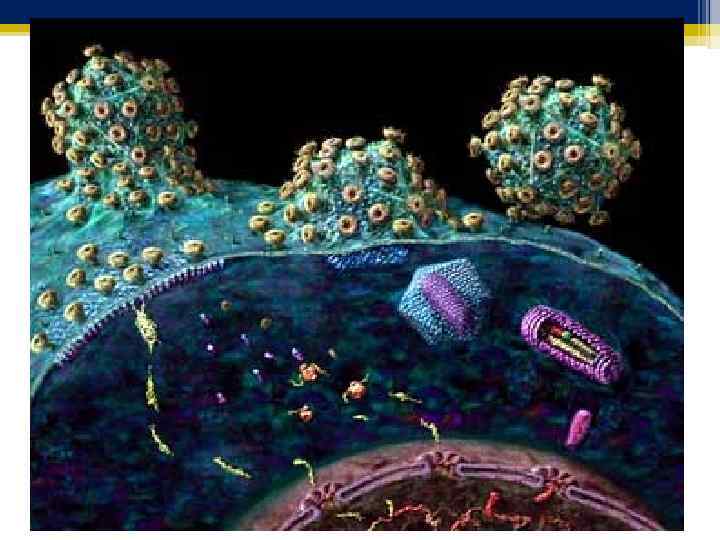
Созревшая полноценная частица вируса – вирион. Морфологию и структуру вирусов изучают с помощью электронного микроскопа. Размер вирусов измеряют в нм, применяя методы электронной микроскопии, ультрафильтрации и ультрацентрифугирования. Размеры вирусов колеблятся в широких пределах : от 1820 нм у парвовирусов (вирус В 19 вызывает инфекционную эритему) до 350 нм у поксивирусов (вирус натуральной оспы).

Форма вирионов чрезвычайно разнообразная, но у некоторых вирусов она может быть специфической Сферическая – вирусы полиомиелита, ротавирусы, ВИЧ. Палочковидная - вирус везикулярного стоматита, табачной мозаики. Пулевидная – вирус бешенства. Плеоморфная (неправильной формы) – коронавирусы (возбудитель SARS). Форма рыболовного крючка – вирус лихорадки Эбола.

Примеры формы вирионов
")
СХЕМА СТРОЕНИЕЯ ВИРИОНА БЕЛКОВАЯ ОБОЛОЧКАКАПСИД ГЛИКОПРОТЕИНОВЫЕ МОЛЕКУЛЫ – РЕЦЕПТОРЫ ФОСФОЛИПОПРОТЕИДНАЯ ОБОЛОЧКА - СУПЕРКАПСИД (ПЕПЛОС) ГЕНОМ (ДНК ИЛИ РНК) ГЕНОМ СВЯЗАН С ВНУТРЕННИМИ БЕЛКАМИ (КАПСИДОМ), ОБРАЗУЯ СЕРДЦЕВИНУ ВИРИОНА, ИЛИ НУКЛЕОКАПСИД

Структура вирионов может быть простой и сложной A – простого строения вирион содержит только нуклеокапсид: нуклеиновую кислоту (ДНК или РНК) и белковую оболочку – капсид (лат. сapsa – футляр); простые вирусы именуются «голыми» , так как они не имеют наружной оболочки. B – сложного строения вирион кроме нуклеокапсида имеет наружную липопротеиновую (мембраноподобную) оболочку, приобретаемая вирусом в момент выхода из клетки хозяина- суперкапсид (пеплос). Такие вирусы называют «одетыми» .

Геном вирусов q. Вирусы содержат только один тип нуклеиновой кислоты, ДНК или РНК, но не оба типа одновременно. Например, вирусы оспы, простого герпеса, Эпштейна-Барр — ДНКсодержащие, а тогавирусы, пикорнавирусы — РНК-содержащие. Геном вирусной частицы гаплоидный, за исключением ретровирусов, которые имеют диплоидный набор генов. q. Наиболее простой вирусный геном содержит 6 генов (кодирует 3 -4 белка), в геномах наиболее сложных вирусов имеется нескольких сотен генов (кодирует более 50 полипептидов).

q. Нуклеиновые кислоты представлены одно- нитевыми молекулами РНК (исключая реовирусы, у которых геном образован двумя нитями РНК) или двухнитевыми молекулами ДНК (исключая парвовирусы, у которых геном образован одной нитью ДНК). У вируса гепатита В нити двухнитевой молекулы ДНК неодинаковы по длине. q Вирусные ДНК образуют циркулярные, ковалентно-сцёпленные суперспирализованные (например, у паповавирусов) или линейные двухнитевые структуры (например, у герпес- и аденовирусов).

q. Вирусные РНК представлены одно- или двухнитевыми молекулами. Однонитевые молекулы могут быть сегментированными — от 2 сегментов у ареновирусов до 11 — у ротавирусов. Наличие сегментов ведёт к увеличению кодирующей ёмкости генома. q Вирусные РНК подразделяют на следующие группы полярности: плюс-нити РНК (+РНК), минуснити РНК (-РНК). У различных вирусов геном могут образовывать нити +РНК либо -РНК, а также двойные нити, одна из которых -РНК, другая (комплементарная ей) — +РНК. q Механизм репликации вирусов определяется особенностями строения его генома, что имеет значение при подборе препаратов для противовирусной терапии.

Форма молекул вирионных нуклеиновых кислот: ДНК 1. линейная 1 -цепочечная; 2. линейная 2 -цепочечная не замкнутая; 3. линейная 2 -цепочечная с замкнутыми концами; 4. кольцевые (цикличные) обычная и суперспиральная; 5. кольцевая с недостроенным участком в одной цепи. РНК 1. линейная 1 -цепочечная цельная; 2. линейная 2 - цепочечная сегментириванная; 3. линейная 1 - цепочечная сегментированная; 4. кольцевая сегментрированная; 5 двойная 1 -цепочечня.

Вирусный капсид q. Капсид – белковая оболочка, которая образована небольшими, повторяющимися белковыми субъединицами– капсомерами (составляющих одну или несколько полимерных макромолекул). q Капсомеры организованы в симметричные конструкции, тип которых постоянен также, как и число самих капсомеров у каждого вида вируса (у аденовирусов капсомеров 252, у вируса полиомиелита - 60, у вируса табачной мозаики – 2130). q Повторение субъединиц в капсидах облегчает их сборку и уменьшает количество генетической информации, необходимой для их синтеза.

Основные типы симметрии вирусных капсидов Спиральный тип Кубический, икосаэдсимметрии (вирусы рический, квазисферимозаичной болезни ческий тип симметрии табака, бешенства и (пикорнавиусы, др. ) аденовирусы и др. ) Смешаный тип симметрии имеют бактериофаги (головка - икосаэдр, отросток – спираль).
 q Наружная оболочка сложноустроенных вирусов, представляет собой липидный слой со встроенными белками,")
Суперкапсид (пеплос) q Наружная оболочка сложноустроенных вирусов, представляет собой липидный слой со встроенными белками, гликопротеидами. q Имеется не у всех вирусов (только у сложноустроенных). q Генетически суперкапсид не принадлежит вирусу - происходит из мембраны клеткихозяина. q Формируется суперкапсид при выходе вирусной частицы из инфицированной клетки (наиболее часто почкованием).
 или булавы.")
Суперкапсид снаружи усеян вирусспецифическими белками, которые по своему виду напоминают шипы (спикули) или булавы. На внутренней поверхности суперкапсида у многих семейств вирусов имеется гидрофобный матриксный белок, который стабилизирует оболочку. Липопротеиновая оболочка вирусов является фрагментом цитоплазматической мембраны или мембран внутренних органелл клеток хозяина и формируется при ввыходе вируса из клетки путем почкования.

Структура сложноустроенного вируса

Химический состав вирусов В 1935 г. У. Стенли получил вирус табачной мозаики в кристаллическом виде, изучил его химический состав и обнаружил, что простые вирусы состоят в основном из нуклеиновой кислоты и белка. В 1946 г. ученый получил за эту работу Нобелевскую премию по химии.

Химический состав вирусов состоит из: нуклеиновой кислоты, белкков, липидов, гликолипидов, гликопротеидов. Вирусы всегда содержат один тип нуклеиновой кислоты (ДНК или РНК), которая составляет от 1 % до 40 % массы вириона. Вирусные геномы содержат информацию, достаточную для синтеза лишь нескольких белков. Масса геномов вирусов достигает от (0, 7 – 1, 5) МД (у парвовирусов) до (150 -200) МД (у вируса натуральной оспы), что в 1 млн. раз меньше, чем в клетки, а длинна - до 0, 093 мм. Число нуклеотидных пар колеблется от 3150 (вирус гепатита В) до 230000 (вирус натуральной оспы).

q Белки вирусов составляют от 70 до 90% массы их вирионов и разделяются на структурные и неструктурные. Структурные – это белки, которые входят в состав зрелых внеклеточных вирионов (суперкапсидные, серцевинные). Они выполняют ряд важных функций: защищают нуклеиновую кислоту от внешнего повреждения, взаимодействуют с мембранами чувствительных клеток и обеспечивают проникновение вируса в клетку, имеют РНК- и ДНК-полимеразную активность и др. Неструктурные (функциональные) белки не входят в состав зрелых вирионов, однако образуются во время их репродукции. Эти белки обеспечивают регуляцию экспрессии вирусного генома, являются предшественниками вирусных белков, способны подавлять клеточный биосинтез.

q. В зависимости от расположения в вирионе, белки разделяются на суперкапсидные, матриксные, белки сердцевины - ассоциируемые с нуклеиновой кислотой. q Липиды содержатся в сложных вирусах и составляют 15 -35 % их массы, они входят в состав суперкапсидной оболочки, образовывая ее двойной липидный слой. Липиды стабилизируют вирусную оболочку, обеспечивают защиту внутренних слоев вирионов от гидрофильных веществ внешней среды, принимают участие в депротеинизации вирионов. Липопротеиды - комплекс липидов клеточной мембраны и вирусных суперкапсидных белков, каторые они приобретают при выделении из клетки во время репродукции.
, достигая 3,")
q Молекулы углеводов вирусов входят в состав гликопротеинов, гликолипидов (в основном суперкапсида), достигая 3, 5 -9 % массы вириона. Они играют важную роль, обеспечивая защиту соответствующих молекул от действия клеточных протеаз, выполняют функцию специфических рецепторов связывания (адсорбции) у вирусов. Состав липидов и углеводов определяется клеткой хозяина, но модифицируется структурными белками вируса.

Ферменты вирусов q. По происхождению вирусные ферменты делятся на три группы: 1. вирионные- входят в состав вирионов (например: ДНК-, РНК-полимеразы многих вирусов, обратная транскриптаза ретровирусов; экзо- и эндонуклеазы; АТФ-аза; нейраминидаза и др. ). 2. вирусиндуцированные - ферменты, структура которых закодирована в геноме вируса, а синтез происходит на рибосомах клетки-хозяина (например: РНК-полимераза пикорна-, тога-, орто- и парамиксовирусов; ДНК – полимераза покс- и герпесвирусов); 3. клеточные, модифицированные вирусом - это ферменты клетки-хозяина, которые не являются вирусспецифическими, но которые участвуют в репродукции вируса.

q По функциональному значению вирусные ферменты можно подразделить на две группы: 1. ферменты, участвующие в процессе репликации и транскрипции вирусной нуклеиновой кислоты (ДНК- полимераза, РНК-полимераза, обратная транскриптаза ретровирусов и др. ); 2. ферменты, участвующие в проникновении вирусного генома в клетку-хозяина и выходу образовавшихся вирионов (например: лизоцим, нейраминидаза, АТФ-аза и др. ).
 (International Committee on Taxonomy of Viruses, ICTV) в")
Международным Комитетом по Таксономии Вирусов (МКТВ) (International Committee on Taxonomy of Viruses, ICTV) в 1966 г. приняты основы принципов действующей сегодня системы таксонов вирусов: 1. Царство (Regnum) 2. Отдел (Division) 3. Класс (Classis) 4. Порядок (Order) 5. Семейство (Family) 6. Род (Genus) 7. Вид (Species) До настоящего времени используется иерархическая система таксонов вирусов не выше порядка.

На начало 2014 года МКТВ представил 7 порядков, 103 семейства, 23 подсемейства, около 450 родов и более 2650 видов классифицированных вирусов (всего описано около 4000 различных вирусов). МКТВ постепенно внедряет в науку и практику международную бинарную номенклатуру (наименование) вирусов, подобно названиям других живых существ: название рода + видовой эпитет. Окончание наименования: порядка – virales; семейства – viridae; подсемейства – virinae; рода – virus; вид – virus.

На современном развитии вирусологии одновременно из унифицированными бинарными наименованиями вирусов, внедряемыми МКТВ, продолжают широко фигурировать «английские тривиальные названия» . Например: Human adenovirus (НАd. V), серотип 12 в соответствии с правилами МКТВ; Herpes Simplex Virus тип 1 (HSV -1) – не по правилам МКТВ.

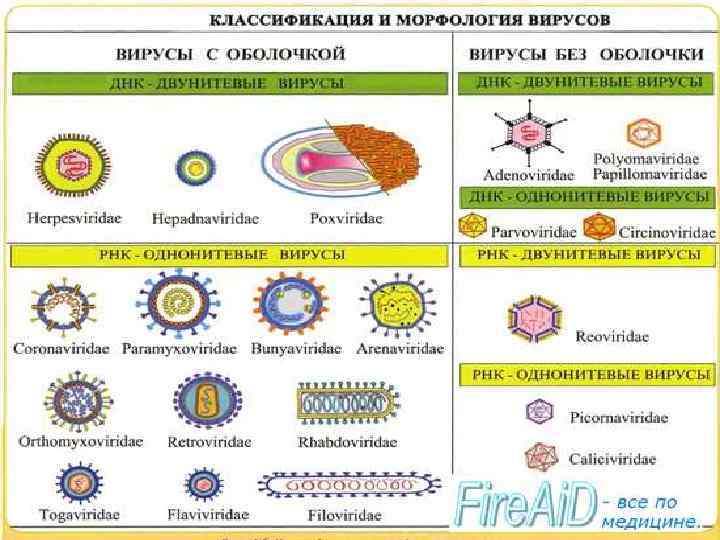
В основу классификации положены следующие основные свойства вирусов: 1. Тип нуклеиновой кислоты. В соответствии с этим все вирусы позвоночных подразделяют на два подтипа - ДНК- и РНК содержащие. 2. Количество и структурные особенности нитей ДНК и РНК: позитивная (+) или негативная (-) РНК; одинарная или двойная ДНК; линейная, кольцевая или сегментированная. 3. Наличие второй липопротеидной оболочки: простые вирусы без второй оболочки; сложные вирусы с наличием суперкапсида. 4. Чувствительность вирионов к органическим растворителям (эфиру, хлороформу).
 капсомеров в белковой оболочке вирионов: кубический, спиральный, смешанный. 6. Количество")
5. Тип укладки (симметрии) капсомеров в белковой оболочке вирионов: кубический, спиральный, смешанный. 6. Количество капсомеров в вирионах: для каждого семейства, рода вирусов количество определенно и постоянно. 7. Диаметр (размер) вирионов в нанометрах: размер вирионов у разных вирусов определенный и постоянный. 8. Организм-хозяин для репродукции вирусов: вирусы позвоночных (человека, животных и птиц), растений, насекомых, простейших, бактерий. 9. Способность агглютинировать эритроциты: с гемагглютинирующими свойствами и без таких свойств.
 Семейство вирусов Наличие суперкапсида Тип симмет рии Структура РНК Вирусы")
Классификация вирусов (РНК-содержащие ) Семейство вирусов Наличие суперкапсида Тип симмет рии Структура РНК Вирусы патогенные для человека Picornaviridae Нет Кубичес кий Однонитчастая, линейная, несегментированая, “плюс” Полиовирус, риновирус, вирус гепатита А Caliciviridae Нет Кубичес кий Однонитчастая, линейная, несегментированая, “плюс” Вирус Норвoлк, вирус гепатита Е Reoviridae Нет Кубичес кий Двухнитчастая, 10 сегментов Реовирус, ротавирус Flaviviridae Да Кубичес кий Однонитчастая, линейная, несегментированая, “плюс” Вирусы клещевого энцефалита, японского энцефалита, желтой лихорадки, лихорадки Западного Нила, гепатита С Togavirus Да Кубичес кий Однонитчастая, линейная, 2 сегменты, “плюс” Вирус краснухи
 Семейство вирусов Наличие супер -капсида Тип Симмет -рии Структура РНК")
Классификация вирусов (РНК- содержащие) Семейство вирусов Наличие супер -капсида Тип Симмет -рии Структура РНК Вирусы патогенные для человека Retroviridae Да Кубичес кий Однонитчастая, линейная, несегментированая, “плюс” ВИЧ, вирус Т-клеточной лейкемии человека Orthomyxovirus Да Спира льный Однонитчастая, линейная, 8 сегментов, “минус” Вирусы гриппа Paramyxoviridae Да Спира льный Однонитчастая, линейная, несегментированая, “минус” Rhabdoviridae Да Спира льный Однонитчастая, линейная, несегментированая, “минус” Вирусы парагрпипа, кори, паротита, респираторносинцитиальный вирус Вирус бешенства
 Семейство вирусов Наличие суперкапсида Тип Симметрии Структура РНК Вирусы патогенные")
Классификация вирусов (РНК- содержащие) Семейство вирусов Наличие суперкапсида Тип Симметрии Структура РНК Вирусы патогенные для человека Filoviridae Да Спира льный Однонитчастая, линейная, несегментирована я, “минус” Вирус Эбола, вирус Марбурга Coronaviridae Да Спира льный Однонитчастая, линейная, несегментирована я, “плюс” Коронавирусы Arenaviridae Да Спира льный Однонитчастая циркулярная, 2 сегменты, “минус” Вирус лимфоцитарного хориоменингита Bunyavirus Да Спира льный Однонитчастая циркулярная, 3 сегменты, “минус” Хантавирусы, вирус Кримской-Конго гемморагической лихорадки, вирус гемморагической лихорадки с почечным синдромом
 Семейство вирусов Наличие суперкапсида Тип Симметри и Структура РНК Вирусы")
Классификация вирусов (РНК- содержащие) Семейство вирусов Наличие суперкапсида Тип Симметри и Структура РНК Вирусы патогенные для человека Bornaviridae Да Кубиче ский Однонитчастая, линейная, несегментирована я, “минус” Вирусы болезни Борна Astroviridae Нет Кубиче ский Однонитчастая, линейная, несегментирована я, “плюс” Астровирусы человека Deltavirus (не классифициро ванный вирус Да неизве стный Однонитчастая, циркулярная, кольцо, “минус” Вирус гепатита Дельта
 Семейство вирусов Наличие суперкапсида Тип симмет рии Структура ДНК Вирусы")
Классификация вирусов (ДНК- содержащие) Семейство вирусов Наличие суперкапсида Тип симмет рии Структура ДНК Вирусы патогенные для человека Papovaviridae Нет Кубический Двухнитчастая, циркулярна Вирус папиломы Adenoviridae Нет Кубический Двухнитчастая, линейная Аденовирус Hepadnavirus Да Кубический Двониткова, дефектна, циркулярна Вирус гепатита В Herpesviridae Да Кубический Двухнитчастая, Вирус простого герпеса 1, 2, линейная опоясывающего герпеса-ветрянки, цитомегаловирус, Эпштейна-Барр вирус Poxviridae Да Смешанный Двухнитчастая, Вирус натуральной оспы, вирус линейная вакцины Parvoviridae Нет Кубичес- Однонитчастая, кий линейная Вирусы гастроэнтерита, инфекционной эритемы, гемолитической болезни

ВЗАИМОДЕЙСТВИЕ ВИРУСОВ С КЛЕТКОЙ ПОДГОТОВКА ВИРУСНОГО ГЕНОМА К ТРАНСКРИПЦИИ И ТРАНСЛЯЦИИ ДЕПРОТЕИНИЗАЦИЯ ВИРИОНА ПРОНИКНОВЕНИЕ ВИРИОНА В КЛЕТКУ РЕПЛИКАЦИЯ ВИРУСНОГО ГЕНОМА СИНТЕЗ ВИРУСНЫХ БЕЛКОВ СБОРКА ВИРИОНОВ АДСОРБЦИЯ ВИРИОНА НА ЦПМ КЛЕТКИ ВЫДЕЛЕНИЕ ВИРИОНОВ ИЗ КЛЕТКИ РАЗЛИЧНЫЕ СТАДИИ ВЗАИМОДЕЙСТВИЯ ВИРУСОВ С КЛЕТКОЙ ПРОИСХОДЯТ В РАЗНЫХ КОМПАРТМЕНТАХРАЗОБЩЕННЫЙ ТИП РЕПЛИКАЦИИ

q. К ранним стадиям взаимодействия вируса с клеткой принадлежат: адсорбция вируса с клеткой вирусов на поверхности клетки, их проникновение (пенетрация) внутрь клетки и «раздевание» (депротеинизация) вириона. q. Поздние стадии репродукции вирусов Поздние (стратегии вирусного генома) включают: синтез вирусных нуклеиновых кислот, синтез ранних (регуляторных) и поздних (регуляторных и структурных) белков, сборку вирионов и выход вирусных частиц из клетки.

Адсорбция вирусов на клетки q. Прикрепление вирусов к поверхности клетки обеспечивается двумя механизмами: неспецифическим и специфическим. Неспецифический определяется силами электростатического взаимодействия, что возникает между химическими группами на поверхности вирусов и клеток, которые несут разные заряды. Специфический механизм (обратимая и необратимая адсорбция) предопределяется комплементарними вирусными и клеточными специфическими рецепторами. Число рецепторов на участках адсорбции может достигать 3000. Они могут иметь белковую, углеводную, липидную природу. Например, рецептором для вирусов гриппа является сиаловая кислота, ацетилхолиновый рецептор – для вируса бешенства, СD 3 рецептор для вируса герпеса,
 СЛИЯНИЕ ОБОЛОЧЕК ВИРИОНА И КЛЕТКИ При")
ВАРИАНТЫ ПРОНИКНОВЕНИЯ ВИРИОНА В КЛЕТКУ РЕЦЕПТОРНЫЙ ЭНДОЦИТОЗ (виропексис) СЛИЯНИЕ ОБОЛОЧЕК ВИРИОНА И КЛЕТКИ При проникновении вируса в клетку способом эндоцитоза определяющую роль играет клеточный поверхностный белок клатрин, а при способе слияния оболочек вирусный белок слияния (F- от fusion - слияние).

«Раздевание» - депротеинизация вирусов q. Внутри клетки-хозяина защитные оболочки вируса (которые тормозят его репликацию) капсид и суперкапсид разрушаются под действием клеточных и вирусных ферментов, геном и внутренние белки вируса освобождаются. Происходит «раздевание» в специализированных участок - лизосомах, аппарате Гольджи.

Стадия синтеза вирусных макромолекул q Реализация генетической информации вирусов по синтезу вирусных макромолекул (нуклеиновых кислот, белков) осуществляется в соответствии с процессами: транскрипции – синтеза информационных РНК (и-РНК), или матричных РНК (м-РНК); трансляции – синтеза белка на рибосомах клетки с участием и-РНК; репликации – синтеза молекул нуклеиновой кислоты, гомологичных/идентичных геному вируса. Механизм репликации (образование вирусных геномов, которые являются точной копией предшественника) зависит от особенностей нуклеиновой кислоты. У разных видов вирусов он неодинаковый. Выделяют 6 основных классов репликации геномов вирусов.

Транскрипция генома вирусов q Транскрипция ДНК-вирусов и вирусов с двунитчатой РНК происходит в ядре клетки, а однонитчатых РНК-вирусов происходит в цитоплазме У двухнитчастых ДНК-содержащих вирусов (герепесвирусы, аденовирусы, вирусы натуральной оспы) сначала происходит деспирилизация ДНК и расхождение ее нитей. На одной из них по принципу комплементарности синтезируется новая нить ДНК, на второй - информационная (матричная) РНК (и-РНК или м-РНК). Этот процесс длится, пока в клетках не образуется достаточное количество нуклеиновых кислот.

q У однонитчастых ДНК-геномных вирусов процесс происходит при условии образования промежуточной двунитчатой формы ДНК. q У двунитчатых РНК-вирусов (реовирусы) их геномная РНК транскрибируется непосредственно в м-РНК. q Среди четырех групп однонитчатых РНК вирусов особым процессов транскрипции характеризуются группы вирусов с плюс-нитями РНК (+РНК) и минус-нитями РНК (-РНК). +РНК - одиночные цепочки, имеющие характерные окончания ( «шапочки» ) для распознавания рибосом, а -РНК – нити не имеют таких распознавательной последовательности (10 -15) нуклеотидов.

Поэтому молекулы +РНК способны непосредственно транслировать генетическую информацию на рибосомах заражённой вирусом клетки, то есть выполнять функции м-РНК для синтеза структурных белков и репликации вирусной РНК. А -РНК – нити не способны транслировать генетическую информацию непосредственно на рибосомах, то есть они не могут функционировать как м-РНК. Для –РНК-вирусов необходима промежуточная транскрипция в +РНК форму.
.")
q Существенно отличается от предыдущих групп РНК содержащих вирусов механизм репликации ретровирусов (онковирусы, ВИЧ). Их процесс репродукции тесно связан с репродукцией ДНК клетки хозяина. Сначала на РНК содержащем геноме вируса происходит синтез гомологичных нитей ДНК с помощью РНК-зависимой ДНКполимеразы (обратной транскриптазы). Двойные нити ДНК вируса в дальнейшем интегрируются в геном клетки-хозяина. В таком состоянии вирус (провирус) длительное время сохраняется в клетке. Позже эта нить ДНК служит матрицей для образования вирусной РНК.
 — американский биохимик,")
Дэвид Балтимор (англ. David Baltimore; род. 7 марта 1938, Нью. Йорк) — американский биохимик, молекулярный биолог и вирусолог. В 1971 году предложил классификацию вирусов в зависимости от типа геномной нуклеиновой кислоты и способа её репликации. Нобелевская премия по физиологии и медицине за 1975 г. За открытия, касающиеся взаимодействия между онкогенными вирусами и генетическим материалом клетки.
 нуклеиновых кислот геномов вирусов и их репликация по Д. Балтимору")
Классы (типы) нуклеиновых кислот геномов вирусов и их репликация по Д. Балтимору

Трансляция – синтез вирусных белков q. Синтез вирусных белков осуществляется в клеточных рибосомах с участием м-РНК. На стадии трансляции происходит считывание генетической информации с м-РНК и перевод ее в последовательность аминокислот белков. Молекулы м-РНК продвигаются в рибосомах в соответствии с последовательностью триплетного кода, который распознают транспортные РНК (т-РНК). Последние несут на специальных участках аминокислоты.

Репликация копий вирусных геномных ДНК и РНК q. Репликацию ДНК-вирусов осуществляют клеточные и вирусные белки ДНКполимеразы (как правило в ядре, за исключением геномной ДНК поксвирусов, которая реплицируется в цитоплазме) а репликацию РНК-вирусов осуществляют вирусспецифические РНК-полимеразы (как привило в цитоплазме, за исключением вирусов с минус-РНК-нитями, которые реплицируются в двух местах: в ядре и цитоплазме).
 осуществляется вирусными ферментами. В основе")
Самосборка вирионов q. Самосборка вирусных частиц (геном + капсид) осуществляется вирусными ферментами. В основе лежит возможность специфического распознавания нуклеиновых кислот и вирусных белков при достижении их определенной концентрации. Подсчитано, что для образования одной вирусной частицы необходимо около 10 тысяч молекул белков, липидов, углеводов, нуклеиновых кислот. Простые вирионы складываются на мембранах эндоплазматичного ретикулуму. У сложных вирусов он начинается в аналогичных участках, а заканчивается на цитоплазматической мембране клетки.

ВЫХОД ВИРИОННОГО ПОТОМСТВА ИЗ КЛЕТКИ ПУТЕМ «ПОЧКОВАНИЯ» С СОХРАНЕНИЕМ ЖИЗНЕСПОСОБНОСТИ КЛЕТКИ-ХОЗЯИНА ПУТЕМ РАЗРУШЕНИЯ (ЛИЗИСА, «взрыва» ) КЛЕТКИ-ХОЗЯИНА
 способ репродукции. Последний (разорванный) заключается в том,")
Особенность репродукции вирусов. Вирусам присущий дизъюнктивный (разорванный) способ репродукции. Последний (разорванный) заключается в том, что синтез генома и белков вируса разорван в пространстве и времени: нуклеиновые кислоты реплицируются в ядре и цитоплазме клетки, белки - в цитоплазме, а сбор целых вирионов может происходить на внутренней поверхности цитоплазматической мембраны. Репродукция вирусов - уникальная система воссоздания чужеродной информации в клетках эукариотов и обеспечивает абсолютное подчинение клеточных структур потребностям вирусов.

Примеры схем циклов репродукции РНК– и ДНК-вирусов

Схема цикла репродукции РНК-вируса
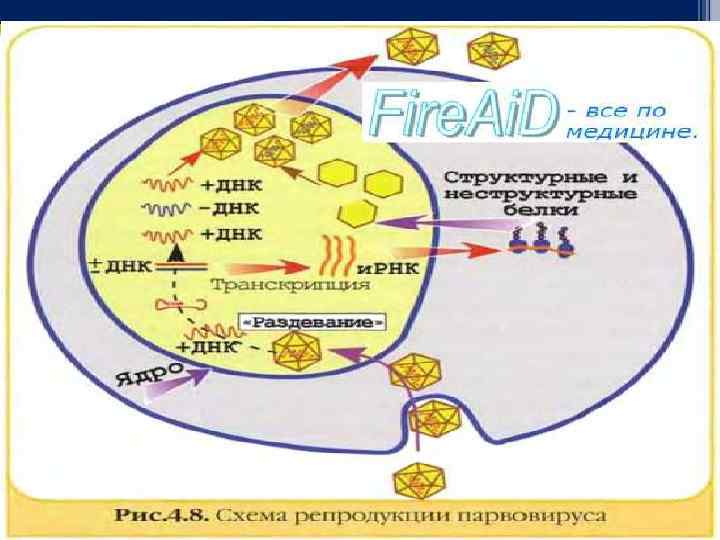
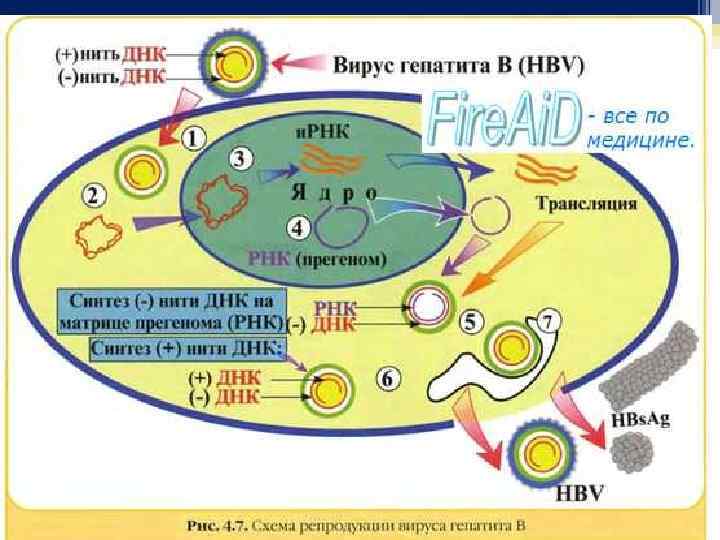

Схема цикла репродукции РНКвируса

ФОРМЫ ВИРУСНОЙ ИНФЕКЦИИ КЛЕТКИ ПРОДУКТИВНАЯ - вирус функционирует в клетке автономно, его репродукция происходит независимо от репродукции клеточного генетического аппарата, при этом образуется новое поколение вирулентных вирусов. АБОРТИВНАЯ – процесс репродукции вируса преры- вается на определенном этапе, «созревание» полноценных вирионов не происходит, но могут образовываться дефектные вирусные частицы, которые потеряли часть своего генома (дефектные интерферирующие частицы, интегрированные геномы, вирусы-спутники и псевдовирионы). ИНТЕГРАТИВНАЯ – геном вируса встраивается в геном клетки и существует там в виде провируса. В результате могут меняться наследственные свойства клетки – вирогения (процесс характерен для РНК и ДНК содержащих онковирусов, ВИЧ).
 (от др. -греч. φᾰγω — \"пожираю\") — вирусы, избирательно поражающие бактериальные")
q. Бактериофаги (фаги) (от др. -греч. φᾰγω — "пожираю") — вирусы, избирательно поражающие бактериальные клетки. Чаще всего бактериофаги размножаются внутри бактерий и вызывают их лизис. Бактериофаг был открыт канадским микробиологом Феликсом д'Эрелем в 1917 году, он же выдвинул предположение, что бактериофаги имеют корпускулярную природу. Однако только после изобретения электронного микроскопа удалось увидеть и изучить ультраструктуру фагов.

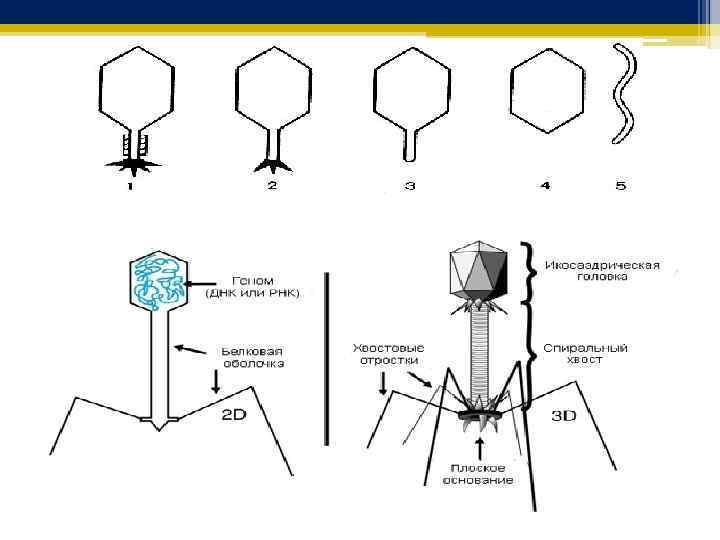
Основные структырные элементы бактериофага • У бактериофагов смешанный тип симметрии: • головка имеет вид кубического (икосаэдр) капсида • от нее отходит длинный (спиральная симметрия) трубчатый отросток – «хвост»

Морфологические типы бактериофагов q. В зависимости от наличия и характера основных структурных компонентов – головки и отростка – бактериофаги подразделяются на морфологические типы, в пределах которых, в свою очередь, возможны различные варианты, что делает мир бактериофагов очень разнообразным. q Бактериофаг может иметь только один из основных структурных компонентов (головку или отросток). Такие бактериофаги иногда называют простыми. 1. Нитчатые бактериофаги не имеют головки и представлены только отростком – это I морфологический тип бактериофагов.

2. Бактериофаги, имеющие только головку, без отростка, составляют II морфологический тип. q Б. Бактериофаги, имеющие и головку и отросток, иногда называют сложными. 1. Отросток у бактериофагов может быть: рудиментным, коротким и без чехла – такие бактериофаги относятся к III морфологическому типу. 2. Отросток у бактериофагов может быть длинным и с чехлом. а. Если отросток при этом не может сокращаться, то такие бактериофаги относятся к IV морфологическому типу. б. Бактериофаги, обладающие сократительным отростком, относятся к V морфологическому типу.

Стадии взаимодействия бактериофага с клеткой бактерии: 1. адсорбция; 2. присоединение; 3. сокращение хвостика; 4. прокалывание клеточной стенки; 5. инъекция ДНК; 6. репродукция/редукция бактериофагов. Репродукция – размножение в клетке фагов, лизис клетки и выход фагов наружу. Редукция – встраивание генома бактериофага в геном клетки (профаг), жизнеспособность клетки сохраняется и профаг размножается синхронно с клеткой.

Типы взаимодействия бактериофагов с клетками бактерий

q Бактериофагов делят на вирулентные, вызывающие лизис клетки с образованием новых частиц, и умеренные (симбиотические), которые адсорбируются клеткой и проникают в неё, но лизиса не вызывают, а остаются в геноме клетки в латентной (скрытой) неинфекционной форме (профаг). Культуры, содержащие латентный фаг, называются лизогенными. Лизогения передаётся потомству бактерии. Воздействуя на лизогенную культуру ультрафиолетовыми или рентгеновскими лучами, перекисью водорода и некоторыми другими веществами, можно значительно увеличить количество клеток, освобождающих фаг (т. н. индукция бактериофагов).

Лизогения широко распространена среди всех видов бактерий и актиномицетов. В ряде случаев многие свойства лизогенной культуры (токсичность, подвижность бактерий, устойчивость к лекарственным препаратам и др. ) зависят от наличия в ней определённых профагов. У возбудителя дифтерии (Сorynebacterium diphtheria) способность продуцировать экзотоксин связана с наличием профага (лизогенией культуры умеренным бактериофагом). Практическое применение бактериофагов в медицине Бактериофаги в медицине применяют для: диагностики заболеваний – фагодиагностика; лечения заболеваний – фаготерапия; профилактики заболеваний - фагопрофилактика; в лабораторной практике для идентификации и типировании возбудителей инфекционных заболеваний – фаготипирование.

q. Фагодиагностика - заключается в выделении фага из организма больного, что косвенно свидетельствует о наличии в материале соответствующих микроорганизмов. Например, обнаружение фагов бактерий Klebsiella rhinoscleromatis (возбудитель заболевания – склеромы) в отделяемом носа при наличии соответствующих признаков болезни подтверждает ее этиологию. q Фаготерапия - лечение фагами некоторых инфекци- онных заболеваний, вызванных, например, шигеллами, сальмонеллами, стафилококком, протеем. Бактериофаги с целью терапии применяют местно путем аппликации на раневую или ожоговую поверхность, введением в полости (брюшную, плевральную, суставную, мочевой пузырь), через рот, а также ректально. Соответственно способу применения препараты бактериофагов выпускают в различных лекарственных формах – жидком виде, в таблетках, мазях, свечах, аэрозолях.
")
q Фагопрофилактика - предупреждения некоторых заболеваний (например, дизентерии, сальмонеллеза, брюшного тифа и др. ) в эпидемичес-ком очаге путем применения бактериофагов лицам с высоким риском заражения. q Фаготипирование - разделение микробов одного и того же вида на различные фаготипы по лизису штаммов микробов типоспецифическими фагами, что важно в эпидемиологической практике, например, при выявлении источника инфекции. Существуют типовые фаги в отношении сальмонелл тифа (96 типов), паратифа В (11 типов), сальмонелл тифимуриум (Бреслау) (13 типов), холерного вибриона (5 типов) , стафилококков (22 типа).

q Определение фагочувствительности является одним из методов дополнительного определения вида и типа микроорганизма, вызвавшего заболевание. В основе ее лежит специфичность действия фага на определенные возбудители. Например, лизис выделенной у больного культуры с помощью специфического фага используют для дифференциации возбудителей чумы от сходных с ним возбудителей псевдотуберкулеза грызунов, холерного вибриона—от других вибрионов, возбудителя гонореи — от сходных микробов, возбудителя склеромы — от палочки Klebsiella pneumoniae. На основании лизиса специфическим фагом определяют родовую и видовую принадлежность шигелл, сальмонелл, бруцелл, холерных вибрионов, которые не агглютинируются специфическими иммунными сыворотками.

q Инфекционную активность бактериофагов определяют с помощью титрования. Титрование в жидкой среде – метод Аппельмана: делают серию разведений бактериофага в жидкой среде, после чего вносят культуру чувствительных к данному фагу бактерий. Титр фага – наибольшее его разведение, где происходит лизис бактерий. Более точным является метод титрования в агаровых слоях по Грациа. Принцип метода состоит в том, что при оптимальном соотношении каждая фаговая частица, размножаясь на бактериях, дает потомство, которое лизирует все бактерии, находящееся на определенном участке плотной среды; указанный процесс проявляется в образовании негативных колоний.

Грациа метод (A. Gratia, 1936 г. , совр. бельг. микробиолог; син. двухслойный метод, метод агаровых слоев) — метод титрования фага, заключающийся в определении количества активных фаговых частиц в 1 мл субстрата путем внесения образца в полужидкий агар (0, 7 %), содержащий чувствительную к фагу культуру, с последующим наслоением смеси на плотный агар (1, 5 %) в чашке Петри, термостатированием и подсчетом количества негативных колоний. Количество колониеобразующих единиц (КОЕ) фага в 1 мл суспензии называют титром фага.

Методы диагностики вирусных инфекций q Методы лабораторной диагностики вирусных инфекций имеют свою специфику, учитывая особенности биологии вирусов. В медицине используются методы лабораторной диагностики вирусных инфекций: § вирусоскопический; § вирусологический; § серологический (или иммунологический); § молекулрногенетический.

q. Вирусоскопический метод - заключается в обнаружении вируса в исследуемом материале под микроскопом. Чаще используют электронный микроскоп, реже – люминесцентный, световая микроскопия из-за очень малых размеров вирусов применяется очень редко. С помощью электронного микроскопа удается определить морфологические особенности вириона, что имеет диагностическое значение. А с помощью светового микроскопа можно выявить внутриклеточные включения, которые образуются в пораженных клетках при некоторых инфекциях.

q. Вирусологический метод - заключается в заражении исследуемым материалом чувствительной биологической модели (лабораторные животные, куриные эмбрионы или культуры клеток), индикации (выявлении) вируса и его последующей идентификации (определении вида). Вирусологический метод по-прежнему остаётся “золотым стандартом” диагностики вирусных инфекций человека, поскольку характеризуется высокой чувствительностью (85 -100 %) и специфичностью (100 %), позволяет получить чистую культуру вируса для дальнейшего изучения, в частности испытания чувствительности к противовирусным препаратам. Из-за длительности выполнения исследования (2 -14 суток) не пригоден для диагностики неотложных состояний (например, при вирусном энцефалите или сепсисе).
 методов – заключается в выявлении специфических вирусных антигенов (Аг) в клиническом")
q Серологический (иммунологический) методов – заключается в выявлении специфических вирусных антигенов (Аг) в клиническом материале от пациента и/или в определении титра противовирусных антител в сыворотке крови больного в динамике болезни с помощью разнообразных серологических (иммунологических) реакций (ИФА, РИФ, РНИФ, РПГА, РТГА, РСК и др. ). При использовании методов серодиагностики обязательным является исследование «парных сывороток» (отобраных с интервалом 10 -14 суток). При этом четырехкратное нарастание титра антител во второй сыворотке в большинстве случаев служит показателем протекающей или недавноперенесенной инфекции. При исследовании одной сыворотки, взятой в острой стадии болезни, диагностическое значение имеют антитела класса Ig М, свидетельствующее об острой инфекции.

q Молекулрно-генетический метод – заключается в непосредственном выявлении геномов возбудителей вирусных инфекций в образцах клинического материала от пациента. Первоначально классическим методом выявления вирусного генома считался высокоспецифичный метод гибридизации нуклеиновых кислот (ДНК/ДНК -гибридизация, ДНК/РНК –гибридизация), но в настоящее время все шире используется для выявления геномов вирусов полимеразная цепная реакция (ПЦР). С помощью молекулярногенетических методов удается диагностировать персистирующие вирусные инфекции, которые с трудом или вообще не удается обнаружить другими методами.
 q Неспецифическая (достигается с помощью")
Этиотропная терапия вирусных инфекций q. Специфическая (использование иммуноглобулинов, вакцин) q Неспецифическая (достигается с помощью интерферонов, их индукторов и иммуномодуляторов) q Химиотерапия (применение химиопрепаратов, подавляющие репродукцию вирусов)

Специфическая иммунотерапия § Активная Вакцины § Вакцина против бешенства, кори, паротита, краснухи, полиомиелита, герпеса и т. д. • Инактивированная вакцина против вируса гриппа (инфлувак, § Пассивная Препараты иммуноглобулинов • Иммуноглобулины человеческие: цитомегаловирусный (против TORCH-инфекций), против герпеса простого. ваксигрипп, флюарикс и др. ) В/в, для лечения и профилактики гриппа, гепатита, кори, краснухи, полиомиелита, бешенства, желтой лихорадки и др. Антитела, в препарате, нейтрализуют вирусы и препятствуют присоединению вирусов к клеткам Ig. G Нормальный иммуноглобулин человека – содержит Ig. G из крови не менее 1000 здоровых доноров

КЛАССИФИКАЦИЯ ПРОТИВОВИРУСНЫХ СРЕДСТВ ШИРОКОГО НЕСПЕЦИФИЧЕСКОГО ДЕЙСТВИЯ Интерфероны Иммуномодуляторы • • Диуцифон – стимуляция ИЛ-2; • Левамизол, Тактивин, Тимозин – нормализация функции Т-лимфоцитов. ИФ лейкоцитар- ный человеческий • α-ИФ (реаферон, роферон, интрон, виферон) • β-ИФ (авонекс, бетасерон) • γ-ИФ (гаммаферон, иммуноферон) Интерфероногены (индукторы ИФ) • Тилорон (амиксин) • Амизон • Дибазол • Теофиллин • Изопринозин (инозиплекс) • Циклоферон

Этапы репродукции вирусов – «мишени» действия основных противовирусных средств Адгезия вируса 1 2 3 «прилипание» вируса к клетке-хозяина проникновение вируса внутрь клетки-хозяина Адсорбция вируса «Раздевание» вируса в клетке хозяина Синтез НК вируса из генетического материала клетки-хозяина 4 Синтез «ранних» вирусных белков Синтез «поздних» вирусных белков Структурные белки Белкиферменты Сборка дочерних популяций Выход новых вирусов из клетки хозяина высвобождение вирусного генома 1. Прямые вирулоциды (оксолин, бонафтон и др. ) 2. Адамантамины (ремантадин) 3. Аналоги нуклеозидов (ацикловир, зидовудин, индинавир и др. ) 4. Интерфероны и индукторы ИФ Основные группы противовирусных средств
 Ингибиторы адгезии, пенетрации синтеза и депротеинизации нуклеиновых кислот (")
НАПРАВЛЕННОСТЬ ДЕЙСТВИЯ ПРОТИВОВИРУСНЫХ СРЕДСТВ (I) Ингибиторы адгезии, пенетрации синтеза и депротеинизации нуклеиновых кислот ( «раздевания» ) будущего вирусов вируса • Ремантадин Аналоги нуклеозидов (римантадин, (пуринов и альгирем) пиримидинов), [взаимодействуют с которые белком встраиваются М 2 вируса гриппа, что в блокирует синтезируемую образование вирусную ионных каналов нуклеиновую в липидной кислоту – оболочке и вход «ложная» ДНК/РНК протонов вируса внутрь вирионов и (летальный для последующее его вируса синтез) «раздевание» ] Ингибиторы синтеза белков структурных и ферментных будущего вируса Препараты интерферонов Прямые вирулоциды = убивают вирусы вне клеток • Оксолин • Госиптол • Риолоксол • Бонафтон • Хелепин и др
 Ингибиторы герпевирусной ДНК-полимеразы • Видарабин • Цитарабин • Индоксуридин")
НАПРАВЛЕННОСТЬ ДЕЙСТВИЯ ПРОТИВОВИРУСНЫХ СРЕДСТВ (II) Ингибиторы герпевирусной ДНК-полимеразы • Видарабин • Цитарабин • Индоксуридин (офтан) Избирательно активируемые тимидинкиназой, блокируют ДНК-полимеразу Ингибиторы обратной транскриптазы ретровирусов, включая ВИЧ: • Ацикловир (зовиракс) • Фамцикловир (фамвир) • Ганцикловир (цимевен) • Зидовудин (азидотимидин) • Диданозин, • Ламивудин • Ставудин Ингибиторы протеаз молекул полипротеинов ( «ранних» белков) Индинавир (криксиван) • Ритонавир (норвир) • Саквинавир (фортоваза) • . Ингибиторы нейраминазы • Зинамивир (реленца) • Оселтамивир (тамифлю)
 – низкомолекулярные индуцибельные белки-цитомены,")
Интерфероны (система ИФ – важнейших факторов неспецифической резистентности организма человека) – низкомолекулярные индуцибельные белки-цитомены, выполняющие в организме контрольно-регуляторную функцию Природа Синтезируют В-лимфоциты α-ИФ периферической крови β-ИФ Образуют фибробласты Продукт γ-ИФ стимулированных Т-лимфоцитов, NKклеток, макрофагов Препараты ИФ: природные рекомбинантные Направление на сохранение клеточного гомеостаза NB! Основное биологическое предназначение ИФ – индукция (вызывание) АНТИВИРУСНОГО состояния клетки Функции Противо. ВИРУСНАЯ Противо. ОПУХОЛЕВАЯ Иммуно. МОДУЛИРУЮЩАЯ Радио. ПРОТЕКТОРНАЯ • п/вирусные ЛС ê п/опухолевые ЛС ê иммунотропные ЛС

Спасибо за внимание!
1МорфолУльтрастрКультивВирусов .ppt