

ПРАКТИКА 3 Ф ЦНС 13-14 студентам.ppt
- Количество слайдов: 80
 Практическое занятие Частная физиология Спинной мозг. № 3. ЦНС.
Практическое занятие Частная физиология Спинной мозг. № 3. ЦНС.
 Группы нервов: I. Шейные нервы. II. Грудные нервы. III. Поясничные") Центральная нервная система (ЦНС) Группы нервов: I. Шейные нервы. II. Грудные нервы. III. Поясничные нервы. IV. Крестцовые нервы. V. Копчиковые нервы.
Центральная нервная система (ЦНС) Группы нервов: I. Шейные нервы. II. Грудные нервы. III. Поясничные нервы. IV. Крестцовые нервы. V. Копчиковые нервы.
 Регуляция функций • Основной принцип регуляции функций с участием нервной системы – рефлекс. • Структурной основой любого рефлекса является рефлекторная дуга.
Регуляция функций • Основной принцип регуляции функций с участием нервной системы – рефлекс. • Структурной основой любого рефлекса является рефлекторная дуга.
 Свойства рефлекторного центра • суммация • одностороннее проведение • задержка возбуждения • трансформация ритма • усвоение ритма
Свойства рефлекторного центра • суммация • одностороннее проведение • задержка возбуждения • трансформация ритма • усвоение ритма
 на изменение внешней и внутренней среды, осуществляемая и контролируемая") Рефлекс ответная реакция организма (органа) на изменение внешней и внутренней среды, осуществляемая и контролируемая ЦНС. Местный рефлекс осуществляется при участии периферических вегетативных ганглиев.
Рефлекс ответная реакция организма (органа) на изменение внешней и внутренней среды, осуществляемая и контролируемая ЦНС. Местный рефлекс осуществляется при участии периферических вегетативных ганглиев.
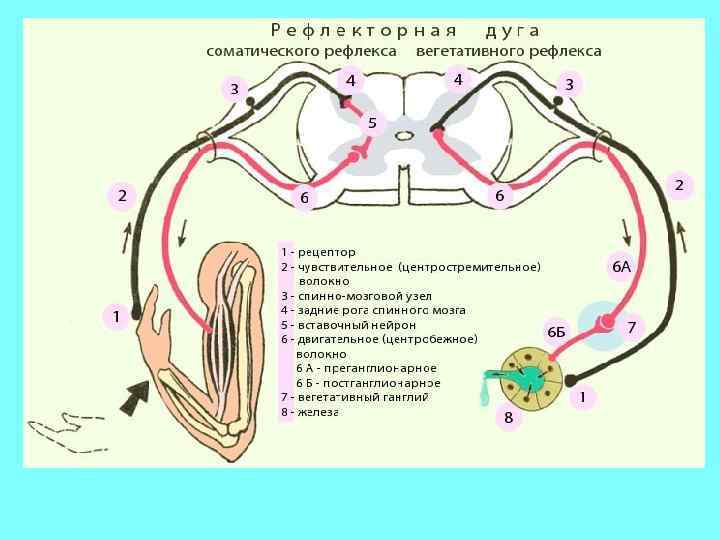
") Рефлекторная дуга 1 - рецепторы 2 - афферентный путь 3 - центр рефлекса (ЦНС) 4 - эфферентный путь 5 - эффектор Обратная связь. Рецептивное поле рефлекса - область расположения рецепторов, раздражение которых вызывает данный рефлекс.
Рефлекторная дуга 1 - рецепторы 2 - афферентный путь 3 - центр рефлекса (ЦНС) 4 - эфферентный путь 5 - эффектор Обратная связь. Рецептивное поле рефлекса - область расположения рецепторов, раздражение которых вызывает данный рефлекс.
, 2 - Рецептор в коже, 3 -") Рефлекторная дуга: 1 - Раздражитель (горячий объект), 2 - Рецептор в коже, 3 - Дендрит чувствительного (центростремительного) нейрона, 4 - Центростремительный нейрон, 5 - Аксон центростремительного нейрона, 6 - Чувствительный нейрон в спинномозговом ганглии, 7 - Бабочка спинного мозга, 8 - Вставочный нейрон, 9 - Двигательный нейрон в передних рогах спинного мозга, 10 - Аксон двигательного нейрона, 11 - Рабочий орган
Рефлекторная дуга: 1 - Раздражитель (горячий объект), 2 - Рецептор в коже, 3 - Дендрит чувствительного (центростремительного) нейрона, 4 - Центростремительный нейрон, 5 - Аксон центростремительного нейрона, 6 - Чувствительный нейрон в спинномозговом ганглии, 7 - Бабочка спинного мозга, 8 - Вставочный нейрон, 9 - Двигательный нейрон в передних рогах спинного мозга, 10 - Аксон двигательного нейрона, 11 - Рабочий орган
 Схема рефлекторной дуги с обратной связью
Схема рефлекторной дуги с обратной связью
 Рефлекторная дуга Взаимодействие рефлексов.
Рефлекторная дуга Взаимодействие рефлексов.
 Классификации рефлексов 1. По расположению рецепторов. 2. По типу рецепторов и ощущений. 3. По расположению жестких элементов центра в ЦНС. 4. По характеру эфферентного пути. 5. По характеру эффектора. 6. По биологическому значению. 7. По происхождению.
Классификации рефлексов 1. По расположению рецепторов. 2. По типу рецепторов и ощущений. 3. По расположению жестких элементов центра в ЦНС. 4. По характеру эфферентного пути. 5. По характеру эффектора. 6. По биологическому значению. 7. По происхождению.
 Классификации рефлексов ЦЕНТР РЕФЛЕКСА ПЛАСТИЧНЫЕ И ЖЕСТКИЕ ЭЛЕМЕНТЫ. СВОЙСТВА ЦЕНТРА: СВОЙСТВА НЕЙРОНОВ И СВОЙСТВА, ВОЗНИКШИЕ В РЕЗУЛЬТАТЕ ИХ ВЗАИМОДЕЙСТВИЯ.
Классификации рефлексов ЦЕНТР РЕФЛЕКСА ПЛАСТИЧНЫЕ И ЖЕСТКИЕ ЭЛЕМЕНТЫ. СВОЙСТВА ЦЕНТРА: СВОЙСТВА НЕЙРОНОВ И СВОЙСТВА, ВОЗНИКШИЕ В РЕЗУЛЬТАТЕ ИХ ВЗАИМОДЕЙСТВИЯ.
 Рефлекторная дуга СИНДРОМ БРОУН-СЕКАРА
Рефлекторная дуга СИНДРОМ БРОУН-СЕКАРА
 Спинной мозг • Является нижним уровнем системы управления движениями. • Состоит из сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов, выходы в виде передних корешков. • Деление на сегменты функционально. • Задние корешки афферентные (чувствительные, центростремительные). • Передние корешки эфферентные (двигательные, центробежные) (закон Белла. Мажанди).
Спинной мозг • Является нижним уровнем системы управления движениями. • Состоит из сегментов, имеющих входы в виде задних корешков, клеточную массу нейронов, выходы в виде передних корешков. • Деление на сегменты функционально. • Задние корешки афферентные (чувствительные, центростремительные). • Передние корешки эфферентные (двигательные, центробежные) (закон Белла. Мажанди).
 Строение сегмента спинного мозга: ядра серого вещества
Строение сегмента спинного мозга: ядра серого вещества
 Спинной мозг и его взаимосвязь с периферическими нервными волокнами - чувствительным, двигательным. 1 - серое вещество, 2 - белое вещество, 3 - дорсальный (задний) корешок, 4 - спинальный ганглий, 5 - рецептор, 6 - эффектор, 7 вентральный (передний) корешок, 8 - интернейрон. Стрелками показано направление распространения возбуждения.
Спинной мозг и его взаимосвязь с периферическими нервными волокнами - чувствительным, двигательным. 1 - серое вещество, 2 - белое вещество, 3 - дорсальный (задний) корешок, 4 - спинальный ганглий, 5 - рецептор, 6 - эффектор, 7 вентральный (передний) корешок, 8 - интернейрон. Стрелками показано направление распространения возбуждения.
 на поверхности тела Буквы соответствуют отделам спинного мозга: С") Распределение дерматомов (областей чувствительной иннервации) на поверхности тела Буквы соответствуют отделам спинного мозга: С — шейному, Т — грудному, L — поясничному и S — крестцовому; цифрами показаны номера сегментов. От каждого участка кожи, обозначенного одинаковыми символами, чувствительные волокна отходят в один сегмент спинного мозга, например, чувствительность кожи в области СЗ шеи обеспечивается 3—м шейным сегментом, область L 5 на голени — 5—м поясничным сегментом.
Распределение дерматомов (областей чувствительной иннервации) на поверхности тела Буквы соответствуют отделам спинного мозга: С — шейному, Т — грудному, L — поясничному и S — крестцовому; цифрами показаны номера сегментов. От каждого участка кожи, обозначенного одинаковыми символами, чувствительные волокна отходят в один сегмент спинного мозга, например, чувствительность кожи в области СЗ шеи обеспечивается 3—м шейным сегментом, область L 5 на голени — 5—м поясничным сегментом.
 Основные функции спинного мозга • 1 - сенсорная: • на уровне нейронов спинного мозга происходит первичный анализ информации от проприорецепторов и кожных рецепторов туловища, конечностей и ряда висцерорецепторов. • К проприорецепторам относят мышечные рецепторы, рецепторы сухожилий, надкостницы, оболочек суставов. • Кожные рецепторы – это рецепторы, расположенные на поверхности и в толще кожного покрова: болевые, температурные, тактильные рецепторы и рецепторы давления.
Основные функции спинного мозга • 1 - сенсорная: • на уровне нейронов спинного мозга происходит первичный анализ информации от проприорецепторов и кожных рецепторов туловища, конечностей и ряда висцерорецепторов. • К проприорецепторам относят мышечные рецепторы, рецепторы сухожилий, надкостницы, оболочек суставов. • Кожные рецепторы – это рецепторы, расположенные на поверхности и в толще кожного покрова: болевые, температурные, тактильные рецепторы и рецепторы давления.
 Основные функции спинного мозга • 2 - ПРОВОДНИКОВАЯ: • восходящие и нисходящие волокна (белое вещество) образуют проводящие пути спинного мозга, по которым передается информация, поступающая от рецепторов и приходят импульсы от вышележащих отделов центральной нервной системы.
Основные функции спинного мозга • 2 - ПРОВОДНИКОВАЯ: • восходящие и нисходящие волокна (белое вещество) образуют проводящие пути спинного мозга, по которым передается информация, поступающая от рецепторов и приходят импульсы от вышележащих отделов центральной нервной системы.
 Проводниковая функция спинного мозга
Проводниковая функция спинного мозга
 Функции спинного мозга • 3 - рефлекторная • Спинальные соматические рефлексы • Проприоцептивные рефлексы • Кожно-мышечные рефлексы • Шейные позно-тонические рефлексы • Перекрестный разгибательный рефлекс • Ритмические рефлексы
Функции спинного мозга • 3 - рефлекторная • Спинальные соматические рефлексы • Проприоцептивные рефлексы • Кожно-мышечные рефлексы • Шейные позно-тонические рефлексы • Перекрестный разгибательный рефлекс • Ритмические рефлексы
 Группы афферентных входов • 1 – образована чувствительными волокнами от проприорецепторов – проприоцептивная чувствительность. • 2 – образована чувствительными волокнами от кожных рецепторов (болевых, температурных, тактильных) – кожная чувствительность. • 3 – образована чувствительными волокнами от висцеральных органов – висцеральная чувствительность.
Группы афферентных входов • 1 – образована чувствительными волокнами от проприорецепторов – проприоцептивная чувствительность. • 2 – образована чувствительными волокнами от кожных рецепторов (болевых, температурных, тактильных) – кожная чувствительность. • 3 – образована чувствительными волокнами от висцеральных органов – висцеральная чувствительность.
 Нейроны спинного мозга • Спинной мозг человека содержит около 13 млн. нейронов, из них 3% - мотонейроны, 97% вставочные. Функционально нейроны спинного мозга можно разделить на 4 группы: • 1. Мотонейроны – альфа- и гамма-мотонейроны клетки передних рогов спинного мозга, аксоны которых образуют передние рога. • 2. Интернейроны получают информацию от спинальных ганглиев, располагаются в задних рогах. Это чувствительные нейроны, реагируют на болевые, температурные, тактильные, вибрационные и проприоцептивные раздражения. • 3. Симпатические (боковые рога спинного мозга, тораколюмбальный отдел) и парасимпатические (сакральный отдел). • 4. Ассоциативные нейроны собственного аппарата спинного мозга, устанавливают связи внутри и между сегментами.
Нейроны спинного мозга • Спинной мозг человека содержит около 13 млн. нейронов, из них 3% - мотонейроны, 97% вставочные. Функционально нейроны спинного мозга можно разделить на 4 группы: • 1. Мотонейроны – альфа- и гамма-мотонейроны клетки передних рогов спинного мозга, аксоны которых образуют передние рога. • 2. Интернейроны получают информацию от спинальных ганглиев, располагаются в задних рогах. Это чувствительные нейроны, реагируют на болевые, температурные, тактильные, вибрационные и проприоцептивные раздражения. • 3. Симпатические (боковые рога спинного мозга, тораколюмбальный отдел) и парасимпатические (сакральный отдел). • 4. Ассоциативные нейроны собственного аппарата спинного мозга, устанавливают связи внутри и между сегментами.
 • • • Виды мотонейронов Мотонейроны делятся на α- и гамма-мотонейроны. Размер альфа-мотонейронов составляет от 40 -70 мкм, гаммамотонейронов – 30 -40 мкм. 1/3 от диаметра переднего корешка занимают аксоны гамма-мотонейронов. Аксон мотонейрона иннервирует мышечные волокна. Скелетные мышцы имеют 2 типа волокон: интрафузальные и экстрафузальные. Интрафузальное волокно находится внутри т. н. мышечного веретена – это специализированный мышечный рецептор, располагающийся в толще скелетной мышцы. Это волокно необходимо для регуляции чувствительности рецептора. Оно управляется гамма-мотонейроном. Все мышечные волокна, принадлежащие данной мышце, и не входящие в состав мышечного веретена, называются экстрафузальными – отвечают за сокращение мышцы. Альфа-мотонейроны иннервируют волокна скелетной мускулатуры (экстрафузальные волокна), обеспечивая мышечные сокращения. Гамма-мотонейроны иннервируют интрафузальные волокна, мышечные веретена являюшиеся рецепторами растяжения. Имеет место сочетанная активация альфа- и гамма-мотонейронов. Аксон альфа-мотонейрона является единственным каналом, соединяющим нервную систему со скелетной мышцей. Только возбуждение альфа-мотонейрона приводит к активации соответствующих мышечных волокон.
• • • Виды мотонейронов Мотонейроны делятся на α- и гамма-мотонейроны. Размер альфа-мотонейронов составляет от 40 -70 мкм, гаммамотонейронов – 30 -40 мкм. 1/3 от диаметра переднего корешка занимают аксоны гамма-мотонейронов. Аксон мотонейрона иннервирует мышечные волокна. Скелетные мышцы имеют 2 типа волокон: интрафузальные и экстрафузальные. Интрафузальное волокно находится внутри т. н. мышечного веретена – это специализированный мышечный рецептор, располагающийся в толще скелетной мышцы. Это волокно необходимо для регуляции чувствительности рецептора. Оно управляется гамма-мотонейроном. Все мышечные волокна, принадлежащие данной мышце, и не входящие в состав мышечного веретена, называются экстрафузальными – отвечают за сокращение мышцы. Альфа-мотонейроны иннервируют волокна скелетной мускулатуры (экстрафузальные волокна), обеспечивая мышечные сокращения. Гамма-мотонейроны иннервируют интрафузальные волокна, мышечные веретена являюшиеся рецепторами растяжения. Имеет место сочетанная активация альфа- и гамма-мотонейронов. Аксон альфа-мотонейрона является единственным каналом, соединяющим нервную систему со скелетной мышцей. Только возбуждение альфа-мотонейрона приводит к активации соответствующих мышечных волокон.
 Мотонейроны !!! • Альфа-мотонейроны получают связь от чувствительных путей от экстрафузальных волокон мышцы. • Гамма-мотонейроны иннервируют интрафузальные мышечные волокна мышечного веретена. Сокращение интрафузального волокна не приводит к сокращению мышцы, но повышает частоту разрядов от рецепторов волокна в спинной мозг.
Мотонейроны !!! • Альфа-мотонейроны получают связь от чувствительных путей от экстрафузальных волокон мышцы. • Гамма-мотонейроны иннервируют интрафузальные мышечные волокна мышечного веретена. Сокращение интрафузального волокна не приводит к сокращению мышцы, но повышает частоту разрядов от рецепторов волокна в спинной мозг.
 (3%) иннервируют волокна скелетной мускулатуры. У них высокая скорость") Характеристика альфамотонейронов • Альфа-мотонейроны (α-мотонейроны) (3%) иннервируют волокна скелетной мускулатуры. У них высокая скорость проведения возбуждения: 70 -120 м/с. • Альфа-мотонейроны делятся на: • 1) альфа-мотонейроны-1 (быстрые) - идут к белым мышечным волокнам, их лабильность (максимальный ритм) - 30 импульсов в секунду. На одном альфа-мотонейроне бывает до 20 тысяч синапсов. Они получают возбуждение от кожных рецепторов, проприоцепторов и вышележащих отделов ЦНС, • 2) альфа-мотонейроны-2 (медленные) иннервируют красные мышечные волокна. Лабильность - 10 -15 импульсов в секунду.
Характеристика альфамотонейронов • Альфа-мотонейроны (α-мотонейроны) (3%) иннервируют волокна скелетной мускулатуры. У них высокая скорость проведения возбуждения: 70 -120 м/с. • Альфа-мотонейроны делятся на: • 1) альфа-мотонейроны-1 (быстрые) - идут к белым мышечным волокнам, их лабильность (максимальный ритм) - 30 импульсов в секунду. На одном альфа-мотонейроне бывает до 20 тысяч синапсов. Они получают возбуждение от кожных рецепторов, проприоцепторов и вышележащих отделов ЦНС, • 2) альфа-мотонейроны-2 (медленные) иннервируют красные мышечные волокна. Лабильность - 10 -15 импульсов в секунду.
 инервируют интрафузальные мышечные волокна мышечных рецепторов. • Гамма-мотонейроны изменяют") Характеристика гамма-мотонейронов • Гамма-мотонейроны (γ-мотонейроны) инервируют интрафузальные мышечные волокна мышечных рецепторов. • Гамма-мотонейроны изменяют активность мышечных рецепторов. • Сами мышечные рецепторы посылают импульсы на альфа-мотонейроны. • Они активизируют собственные альфамотонейроны и тормозят мотонейроны мышцантагонистов. • Лабильность высокая - 200 импульсов в секунду, но зато небольшая скорость проведения возбуждения - 10 -40 м/с.
Характеристика гамма-мотонейронов • Гамма-мотонейроны (γ-мотонейроны) инервируют интрафузальные мышечные волокна мышечных рецепторов. • Гамма-мотонейроны изменяют активность мышечных рецепторов. • Сами мышечные рецепторы посылают импульсы на альфа-мотонейроны. • Они активизируют собственные альфамотонейроны и тормозят мотонейроны мышцантагонистов. • Лабильность высокая - 200 импульсов в секунду, но зато небольшая скорость проведения возбуждения - 10 -40 м/с.
 Мышечные волокна • экстрафузальные - выполняют всю работу сокращения мышцы, • интрафузальные - специализированные для детектирования натяжения. • ИВ получают спиральные окончания чувствительных нервных волокон, образуя рецептор натяжения - мышечное веретено. • Фактически существует три типа ИВ: • два типа волокон с ядерной сумкой (динамические и статические) и • волокна с ядерной цепочкой.
Мышечные волокна • экстрафузальные - выполняют всю работу сокращения мышцы, • интрафузальные - специализированные для детектирования натяжения. • ИВ получают спиральные окончания чувствительных нервных волокон, образуя рецептор натяжения - мышечное веретено. • Фактически существует три типа ИВ: • два типа волокон с ядерной сумкой (динамические и статические) и • волокна с ядерной цепочкой.
 группа из трех интрафузальных волокон. In vivo веретено окружено экстрафузальными волокнами. Оконечности интрафузальных") (А) группа из трех интрафузальных волокон. In vivo веретено окружено экстрафузальными волокнами. Оконечности интрафузальных волокон содержат актомиозиновую сократительную машинерию и, следовательно имеет поперечно-полосатую структуру. Центральный участок ИВ несколько расширен и окружен соединительнотканной капсулой. Он иннервируется двумя типами гамма-двигательных волокон. Эти волокна входят в капсулу и направляются к оконечностям ИВ, где образуют с ними соединение. М - гамма-двигательное волокно; К - капсула; С - 1 а и II чувствительные волокна; ИФ - интрафузальные волокна; НМС - нервномышечное соединение. (Б) Схематический рисунок показывает пять типов ИВ, обнаруженных в мышечном веретене млекопитающих. ДВС - динамическое волокно с ядерной сумкой; Ду. ДВ - динамическое двигательное волокно; ВЦ - волокно с ядерной цепочкой; ПО - первичное окончание; СВС - статическое волокно с ядерной сумкой; СО - статическое окончание; Су. ДВ - статическое гамма-двигательное волокно. По Kandel, Schwartz and Jessell, 1991
(А) группа из трех интрафузальных волокон. In vivo веретено окружено экстрафузальными волокнами. Оконечности интрафузальных волокон содержат актомиозиновую сократительную машинерию и, следовательно имеет поперечно-полосатую структуру. Центральный участок ИВ несколько расширен и окружен соединительнотканной капсулой. Он иннервируется двумя типами гамма-двигательных волокон. Эти волокна входят в капсулу и направляются к оконечностям ИВ, где образуют с ними соединение. М - гамма-двигательное волокно; К - капсула; С - 1 а и II чувствительные волокна; ИФ - интрафузальные волокна; НМС - нервномышечное соединение. (Б) Схематический рисунок показывает пять типов ИВ, обнаруженных в мышечном веретене млекопитающих. ДВС - динамическое волокно с ядерной сумкой; Ду. ДВ - динамическое двигательное волокно; ВЦ - волокно с ядерной цепочкой; ПО - первичное окончание; СВС - статическое волокно с ядерной сумкой; СО - статическое окончание; Су. ДВ - статическое гамма-двигательное волокно. По Kandel, Schwartz and Jessell, 1991
 Мотонейрон спинного мозга позвоночных. Указаны функции разных его частей
Мотонейрон спинного мозга позвоночных. Указаны функции разных его частей
 Интернейроны • Промежуточные нейроны. Генерируют импульсы с частотой до 1000 в секунду, фоновоактивные. На их дендритах имеется до 500 синапсов. • Устанавливают связи между структурами спинного мозга и обеспечивают влияния восходящих и нисходящих проводящих путей на клетки отдельных сегментов спинного мозга. • Способны тормозить активность нейронов для сохранения направленности пути возбуждения (клетки Реншоу).
Интернейроны • Промежуточные нейроны. Генерируют импульсы с частотой до 1000 в секунду, фоновоактивные. На их дендритах имеется до 500 синапсов. • Устанавливают связи между структурами спинного мозга и обеспечивают влияния восходящих и нисходящих проводящих путей на клетки отдельных сегментов спинного мозга. • Способны тормозить активность нейронов для сохранения направленности пути возбуждения (клетки Реншоу).
 Вставочные соматические нейроны • Обеспечивают связь между чувствительными афферентными нейронами и эфферентными мотонейронами спинного мозга. Также вставочные нейроны связывают нейроны спинного мозга с ядрами ствола мозга а через них с корой больших полушарий. Лабильность у них высокая - 1000 имп/сек. • Вставочные нейроны вегетативной НС • Симпатическая НС (в боковых рогах грудного, поясничного и шейного отделов). • Парасимпатическая НС (в крестцовом отделе спинного мозга). • Ассоциативные нейроны • Это внутренний аппарат спинного мозга, обеспечивающий связь между его сегментами, а также внутри сегментов. Ассоциативные нейроны участвуют в координации позы, тонуса мышц и движений. • Нейроны ретикулярной формации спинного мозга - образуют тонкие перекладины серого вещества, идущего в разных направлениях. Сосредоточены в шейных и верхнегрудных сегментах. Обеспечивают поддержание тонуса спинного мозга.
Вставочные соматические нейроны • Обеспечивают связь между чувствительными афферентными нейронами и эфферентными мотонейронами спинного мозга. Также вставочные нейроны связывают нейроны спинного мозга с ядрами ствола мозга а через них с корой больших полушарий. Лабильность у них высокая - 1000 имп/сек. • Вставочные нейроны вегетативной НС • Симпатическая НС (в боковых рогах грудного, поясничного и шейного отделов). • Парасимпатическая НС (в крестцовом отделе спинного мозга). • Ассоциативные нейроны • Это внутренний аппарат спинного мозга, обеспечивающий связь между его сегментами, а также внутри сегментов. Ассоциативные нейроны участвуют в координации позы, тонуса мышц и движений. • Нейроны ретикулярной формации спинного мозга - образуют тонкие перекладины серого вещества, идущего в разных направлениях. Сосредоточены в шейных и верхнегрудных сегментах. Обеспечивают поддержание тонуса спинного мозга.
 Вегетативные нейроны: • симпатического отдела располагаются в боковых рогах серого вещества нижних шейных, грудных и верхних поясничных сегментов спинного мозга; фоновоактивные; • парасимпатического отдела располагаются в крестцовом отделе спинного мозга; фоновоактивные.
Вегетативные нейроны: • симпатического отдела располагаются в боковых рогах серого вещества нижних шейных, грудных и верхних поясничных сегментов спинного мозга; фоновоактивные; • парасимпатического отдела располагаются в крестцовом отделе спинного мозга; фоновоактивные.
 и вегетативные. •") Рефлекторная функция спинного мозга • Спинальные рефлексы подразделяют на соматические (двигательные) и вегетативные. • Двигательные рефлексы в свою очередь делятся на тонические (направлены на поддержание мышечного тонуса, поддержание конечностей и всего туловища в определенном статическом положении) и фазические (обеспечивают движение конечностей и туловища). • К тоническим относятся: миотатический рефлекс, шейные тонические рефлексы положения, рефлекс опоры (впервые их описание дал голландский физиолог Рудольф Магнус, 1924 г. ), сгибательный тонический рефлекс. • К фазическим рефлексам относят: сухожильные рефлексы, рефлексы на укорочение с телец Гольджи, подошвенные, брюшные, сгибательные защитные, разгибательный перекрестный, ритмические.
Рефлекторная функция спинного мозга • Спинальные рефлексы подразделяют на соматические (двигательные) и вегетативные. • Двигательные рефлексы в свою очередь делятся на тонические (направлены на поддержание мышечного тонуса, поддержание конечностей и всего туловища в определенном статическом положении) и фазические (обеспечивают движение конечностей и туловища). • К тоническим относятся: миотатический рефлекс, шейные тонические рефлексы положения, рефлекс опоры (впервые их описание дал голландский физиолог Рудольф Магнус, 1924 г. ), сгибательный тонический рефлекс. • К фазическим рефлексам относят: сухожильные рефлексы, рефлексы на укорочение с телец Гольджи, подошвенные, брюшные, сгибательные защитные, разгибательный перекрестный, ритмические.
 Спинальные соматические рефлексы • • Проприоцептивные рефлексы Кожно-мышечные рефлексы Шейные позно-тонические рефлексы Перекрестный разгибательный рефлекс • Ритмические рефлексы
Спинальные соматические рефлексы • • Проприоцептивные рефлексы Кожно-мышечные рефлексы Шейные позно-тонические рефлексы Перекрестный разгибательный рефлекс • Ритмические рефлексы
 Миотатический рефлекс • рефлекс растяжения, например, когда человек занимает вертикальное положение, то за счет гравитационных сил он может упасть (сгибание в суставах нижних конечностей), но с участием миотатических рефлексов это не происходит, т. к. при растяжении мышцы активируются мышечные веретена, которые расположены параллельно экстрафузальным волокнам скелетной мышцы. • Импульсация от мышечных рецепторов идет через афферентный нейрон и попадает на альфа-мотонейроны данной мышцы. • В результате происходит укорочение экстрафузальных водокон. • Тем самым длина мышцы возвращается к исходной. • Миотатический рефлекс свойственен всем мышцам, хорошо выражен и легко вызывается у мышц сгибателей, направлен против гравитационных сил, для поддержания равновесия, мышечного тонуса. • Надо отметить, что импульсация от рецепторов одновременно через вставочные тормозные клетки Реншоу попадает на альфамотонейроны антагониста этой мышцы, поэтому при укорочении агониста мышца-антагонист не препятствует этому процессу.
Миотатический рефлекс • рефлекс растяжения, например, когда человек занимает вертикальное положение, то за счет гравитационных сил он может упасть (сгибание в суставах нижних конечностей), но с участием миотатических рефлексов это не происходит, т. к. при растяжении мышцы активируются мышечные веретена, которые расположены параллельно экстрафузальным волокнам скелетной мышцы. • Импульсация от мышечных рецепторов идет через афферентный нейрон и попадает на альфа-мотонейроны данной мышцы. • В результате происходит укорочение экстрафузальных водокон. • Тем самым длина мышцы возвращается к исходной. • Миотатический рефлекс свойственен всем мышцам, хорошо выражен и легко вызывается у мышц сгибателей, направлен против гравитационных сил, для поддержания равновесия, мышечного тонуса. • Надо отметить, что импульсация от рецепторов одновременно через вставочные тормозные клетки Реншоу попадает на альфамотонейроны антагониста этой мышцы, поэтому при укорочении агониста мышца-антагонист не препятствует этому процессу.
 Миотатический рефлекс Рефлекс с мышечных веретен, быстрый, моносинаптический. Физиологическое значение – механизм стабилизации длины мышцы.
Миотатический рефлекс Рефлекс с мышечных веретен, быстрый, моносинаптический. Физиологическое значение – механизм стабилизации длины мышцы.
 Характеристика рефлексов: шейные тонические рефлексы • Рецептивным полем шейных тонических рефлексов положения являются проприорецепторы мышц шеи и фасций, покрывающих шейный участок позвоночника. • Центральная часть рефлекторной дуги имеет полисинаптический характер, т. е. включает вставочные нейроны. • Рефлекторная реакция вовлекает мышцы туловища и конечностей. • Кроме спинного мозга, в ней участвуют и моторные ядра мозгового ствола, иннервирующие мышцы глазных яблок. • Шейные тонические рефлексы возникают при поворотах и наклонах головы, что вызывает растяжение мышц шеи и активирует рецептивное поле рефлекса.
Характеристика рефлексов: шейные тонические рефлексы • Рецептивным полем шейных тонических рефлексов положения являются проприорецепторы мышц шеи и фасций, покрывающих шейный участок позвоночника. • Центральная часть рефлекторной дуги имеет полисинаптический характер, т. е. включает вставочные нейроны. • Рефлекторная реакция вовлекает мышцы туловища и конечностей. • Кроме спинного мозга, в ней участвуют и моторные ядра мозгового ствола, иннервирующие мышцы глазных яблок. • Шейные тонические рефлексы возникают при поворотах и наклонах головы, что вызывает растяжение мышц шеи и активирует рецептивное поле рефлекса.
 • Эффекторным звеном шейных рефлексов могут быть не только мышцы конечностей, но и глазная мускулатура. Рефлекторные влияния на глазные мышцы обеспечивают компенсаторную установку глазных яблок при изменении положения головы. Любое отклонение головы вызывает движение глазных яблок в противоположном направлении, и таким образом рефлекторно сохраняется правильная зрительная ориентация животного. • Все шейные рефлексы являются полисинаптическими. Импульсы от проприоцепторов шейных мышц идут в соответствующие центры продолговатого мозга, а оттуда нисходящие команды поступают или к мотонейронам спинного мозга, или к двигательному ядру отводящего нерва, и производят определенное влияние на позный тонус и на глазную мускулатуру. • Вестибулярные рефлексы неразрывно связаны с шейными тоническими рефлексами и в естественных условиях дополняют их. Вестибулярные рефлексы не зависят от положения головы относительно туловища и в чистом виде могут быть получены при фиксации головы по отношению к туловищу или при выключении проприоцепторов шейных мышц новокаиновой блокадой. • Вестибулярные рефлексы, согласно классификации Магнуса, подразделяются на статические и статокинетические.
• Эффекторным звеном шейных рефлексов могут быть не только мышцы конечностей, но и глазная мускулатура. Рефлекторные влияния на глазные мышцы обеспечивают компенсаторную установку глазных яблок при изменении положения головы. Любое отклонение головы вызывает движение глазных яблок в противоположном направлении, и таким образом рефлекторно сохраняется правильная зрительная ориентация животного. • Все шейные рефлексы являются полисинаптическими. Импульсы от проприоцепторов шейных мышц идут в соответствующие центры продолговатого мозга, а оттуда нисходящие команды поступают или к мотонейронам спинного мозга, или к двигательному ядру отводящего нерва, и производят определенное влияние на позный тонус и на глазную мускулатуру. • Вестибулярные рефлексы неразрывно связаны с шейными тоническими рефлексами и в естественных условиях дополняют их. Вестибулярные рефлексы не зависят от положения головы относительно туловища и в чистом виде могут быть получены при фиксации головы по отношению к туловищу или при выключении проприоцепторов шейных мышц новокаиновой блокадой. • Вестибулярные рефлексы, согласно классификации Магнуса, подразделяются на статические и статокинетические.
 – при стоянии на поверхности усиливается тонус мышц") Характеристика рефлексов • Рефлекс опоры (отталкивания) – при стоянии на поверхности усиливается тонус мышц разгибателей. • Сгибательный тонический рефлекс наблюдается, например, у лягушки или у кролика, при котором характерно подогнутое положение конечностей. Этот рефлекс направлен на поддержание определенной позы, что возможно при наличии определенного тонуса мышц. • Сухожильный рефлекс – рефлекс на укорочение с телец Гольджи • Подошвенный рефлекс – раздражение кожи стопы приводит к подошвенному сгибанию пальцев и стопы нижней конечности.
Характеристика рефлексов • Рефлекс опоры (отталкивания) – при стоянии на поверхности усиливается тонус мышц разгибателей. • Сгибательный тонический рефлекс наблюдается, например, у лягушки или у кролика, при котором характерно подогнутое положение конечностей. Этот рефлекс направлен на поддержание определенной позы, что возможно при наличии определенного тонуса мышц. • Сухожильный рефлекс – рефлекс на укорочение с телец Гольджи • Подошвенный рефлекс – раздражение кожи стопы приводит к подошвенному сгибанию пальцев и стопы нижней конечности.
 Характеристика рефлексов • Брюшные рефлексы – напряжение брюшных мышц, возникающее при ноцицептивных афферентных влияниях. Это защитный рефлекс. • Сгибательные защитные рефлексы - возникают при раздражении болевых рецепторов кожи, мышц и внутренних органов направлены на избежание различных повреждающих воздействий. • Разгибательный перекрестный рефлекс: рефлекторное сгибание одной из конечностей нередко сопровождается сокращением контрлатеральной конечности, на которую в естественных условиях (при ходьбе) переносится дополнительный вес тела.
Характеристика рефлексов • Брюшные рефлексы – напряжение брюшных мышц, возникающее при ноцицептивных афферентных влияниях. Это защитный рефлекс. • Сгибательные защитные рефлексы - возникают при раздражении болевых рецепторов кожи, мышц и внутренних органов направлены на избежание различных повреждающих воздействий. • Разгибательный перекрестный рефлекс: рефлекторное сгибание одной из конечностей нередко сопровождается сокращением контрлатеральной конечности, на которую в естественных условиях (при ходьбе) переносится дополнительный вес тела.
 Характеристика рефлексов • К ритмическим рефлексам у млекопитающих относится чесательный рефлекс. • Его аналогом у земноводных является потирательный рефлекс. • Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, устанавливающих конечность в определенное положение к кожной поверхности.
Характеристика рефлексов • К ритмическим рефлексам у млекопитающих относится чесательный рефлекс. • Его аналогом у земноводных является потирательный рефлекс. • Ритмические рефлексы характеризуются координированной работой мышц конечностей и туловища, правильным чередованием сгибания и разгибания конечностей наряду с тоническим сокращением приводящих мышц, устанавливающих конечность в определенное положение к кожной поверхности.
 Характеристика рефлексов • Шагательный рефлекс – согласованная двигательная активность верхних и нижних конечностей. Для осуществления этого рефлекса необходимо межсегментарное взаимодействие мышц рук, ног и туловища. Механизмы шагательных движений заложены в спинном мозге, но включение спинального механизма производится со среднего мозга. • Вегетативные спинальные рефлексы: сосудистые, потоотделение, мочеиспускание, дефекация. • Обеспечивают реакцию внутренних органов, сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов.
Характеристика рефлексов • Шагательный рефлекс – согласованная двигательная активность верхних и нижних конечностей. Для осуществления этого рефлекса необходимо межсегментарное взаимодействие мышц рук, ног и туловища. Механизмы шагательных движений заложены в спинном мозге, но включение спинального механизма производится со среднего мозга. • Вегетативные спинальные рефлексы: сосудистые, потоотделение, мочеиспускание, дефекация. • Обеспечивают реакцию внутренних органов, сосудистой системы на раздражение висцеральных, мышечных, кожных рецепторов.
 Рефлексы спинного мозга А - коленный; Б - ахиллов; В - подошвенный; Г – сгибательно-локтевой; Д – разгибательнолоктевой; Е - брюшной.
Рефлексы спинного мозга А - коленный; Б - ахиллов; В - подошвенный; Г – сгибательно-локтевой; Д – разгибательнолоктевой; Е - брюшной.
 Вестибулярные рефлексы • неразрывно связаны с шейными тоническими рефлексами и в естественных условиях дополняют их; • не зависят от положения головы относительно туловища и в чистом виде могут быть получены при фиксации головы по отношению к туловищу или при выключении проприоцепторов шейных мышц новокаиновой блокадой. • Вестибулярные рефлексы, согласно классификации Магнуса, подразделяются на статические и статокинетические.
Вестибулярные рефлексы • неразрывно связаны с шейными тоническими рефлексами и в естественных условиях дополняют их; • не зависят от положения головы относительно туловища и в чистом виде могут быть получены при фиксации головы по отношению к туловищу или при выключении проприоцепторов шейных мышц новокаиновой блокадой. • Вестибулярные рефлексы, согласно классификации Магнуса, подразделяются на статические и статокинетические.
 Статические и статокинетические рефлексы • Шейные тонические рефлексы запускаются при возбуждении проприоцепторов мышц шеи. Впервые они описаны голландским физиологом Р. Магнусом на кошках с перерезанным на уровне шатра мозжечка стволом мозга. • В чистом виде шейные рефлексы можно наблюдать при разрушении вестибулярного аппарата, дающего дополнительную информацию о положении головы в пространстве. • Как показали опыты Р. Магнуса, запрокидывание головы кошки назад в сагиттальной плоскости вызывает возрастание тонуса мышц-разгибателей передних конечностей и одновременно ослабление экстензорного (разгибательного) тонуса задних. • Напротив, наклон головы вниз вызывает обратный процесс, когда тонус разгибателей передних конечностей снижается, а задних - возрастает. • Нарушение равновесия за счет поворота головы вправо или влево относительно продольной оси вызывает компенсаторное усиление тонуса разгибателей тех конечностей, в сторону которых повернута голова. Это так называемый рефлекс вращения по Магнусу.
Статические и статокинетические рефлексы • Шейные тонические рефлексы запускаются при возбуждении проприоцепторов мышц шеи. Впервые они описаны голландским физиологом Р. Магнусом на кошках с перерезанным на уровне шатра мозжечка стволом мозга. • В чистом виде шейные рефлексы можно наблюдать при разрушении вестибулярного аппарата, дающего дополнительную информацию о положении головы в пространстве. • Как показали опыты Р. Магнуса, запрокидывание головы кошки назад в сагиттальной плоскости вызывает возрастание тонуса мышц-разгибателей передних конечностей и одновременно ослабление экстензорного (разгибательного) тонуса задних. • Напротив, наклон головы вниз вызывает обратный процесс, когда тонус разгибателей передних конечностей снижается, а задних - возрастает. • Нарушение равновесия за счет поворота головы вправо или влево относительно продольной оси вызывает компенсаторное усиление тонуса разгибателей тех конечностей, в сторону которых повернута голова. Это так называемый рефлекс вращения по Магнусу.
 Статические рефлексы • К числу статических вестибулярных рефлексов относят рефлексы выпрямления, направленные на переход животного из неестественной позы в обычное для него положение. • Пример: переворачивание кошки в воздухе при падении спиной вниз. За очень короткое время падения животное успевает занять нормальное по отношению к гравитационному полю положение и упасть сразу на все четыре лапы. • В процессе переворачивания выпрямительные рефлексы совершаются в определенной последовательности. Сначала за счет вестибулярного выпрямительного рефлекса восстанавливается нормальное положение головы - мордой вниз. Затем изменение положения головы возбуждает проприоцепторы шейных мышц и они запускают шейный выпрямительный рефлекс, в результате которого вслед за головой туловище также возвращается в нормальное положение. • Таким образом, в естественных условиях вестибулярные рефлексы выпрямления дополняются шейными, причем ведущую роль в их осуществлении играет положение головы, где расположены дистантные рецепторы.
Статические рефлексы • К числу статических вестибулярных рефлексов относят рефлексы выпрямления, направленные на переход животного из неестественной позы в обычное для него положение. • Пример: переворачивание кошки в воздухе при падении спиной вниз. За очень короткое время падения животное успевает занять нормальное по отношению к гравитационному полю положение и упасть сразу на все четыре лапы. • В процессе переворачивания выпрямительные рефлексы совершаются в определенной последовательности. Сначала за счет вестибулярного выпрямительного рефлекса восстанавливается нормальное положение головы - мордой вниз. Затем изменение положения головы возбуждает проприоцепторы шейных мышц и они запускают шейный выпрямительный рефлекс, в результате которого вслед за головой туловище также возвращается в нормальное положение. • Таким образом, в естественных условиях вестибулярные рефлексы выпрямления дополняются шейными, причем ведущую роль в их осуществлении играет положение головы, где расположены дистантные рецепторы.
 Различают 3 способа связи волокон нисходящих путей с альфа-мотонейронами: • 1. Прямое нисходящее влияние на альфамотонейрон • 2. Опосредованно через вставочный нейрон • 3. Активация гамма-мотонейрона и через интрафузальное волокно к альфамотонейрону. • Гамма-мотонейроны активируют инрафузальные мышечные волокна, в результате чего активируются афферентные нервные волокна и поток импульсов идет на альфа-мотонейроны или на вставочные мотонейроны, а от них к альфамотонейронам – это называется гамма-петля.
Различают 3 способа связи волокон нисходящих путей с альфа-мотонейронами: • 1. Прямое нисходящее влияние на альфамотонейрон • 2. Опосредованно через вставочный нейрон • 3. Активация гамма-мотонейрона и через интрафузальное волокно к альфамотонейрону. • Гамма-мотонейроны активируют инрафузальные мышечные волокна, в результате чего активируются афферентные нервные волокна и поток импульсов идет на альфа-мотонейроны или на вставочные мотонейроны, а от них к альфамотонейронам – это называется гамма-петля.
 В состав гамма-петли входят: - супраспинальные моторные пути, - вставочные нейроны, - нисходящая ретикулярная система, - альфа- и гамма-нейроны.
В состав гамма-петли входят: - супраспинальные моторные пути, - вставочные нейроны, - нисходящая ретикулярная система, - альфа- и гамма-нейроны.
 Функции γ-петли Периферическая регуляция осуществляется с участием гамма-петли, в состав которой входят супраспинальные моторные пути, вставочные нейроны, нисходящая ретикулярная система, альфа- и гамма-нейроны. • Активация γпетли обеспечивает укорочение мышцы с минимальными ошибками (обычно имеет место α-γ-коактивация) γ
Функции γ-петли Периферическая регуляция осуществляется с участием гамма-петли, в состав которой входят супраспинальные моторные пути, вставочные нейроны, нисходящая ретикулярная система, альфа- и гамма-нейроны. • Активация γпетли обеспечивает укорочение мышцы с минимальными ошибками (обычно имеет место α-γ-коактивация) γ
 α-γ-коактивация • За счет супраспинальных влияний и с участием интернейронов практически одновременно активируются α-мотонейроны и γ-мотонейроны
α-γ-коактивация • За счет супраспинальных влияний и с участием интернейронов практически одновременно активируются α-мотонейроны и γ-мотонейроны
 сгибательный, 2) реципрокного торможения мышцыантагониста, 3) перекрестный разгибательный") Рефлексы: 1) сгибательный, 2) реципрокного торможения мышцыантагониста, 3) перекрестный разгибательный
Рефлексы: 1) сгибательный, 2) реципрокного торможения мышцыантагониста, 3) перекрестный разгибательный
 Коленный рефлекс Стимул автоматические, повторяющиеся, стереотипные ответы. Простые рефлексы реализуются нейронами спинного мозга.
Коленный рефлекс Стимул автоматические, повторяющиеся, стереотипные ответы. Простые рефлексы реализуются нейронами спинного мозга.
 Рефлекторная дуга коленного рефлекса
Рефлекторная дуга коленного рефлекса
 Гольджи") Рецепторы двигательных систем • Мышечные веретена • Сухожильные органы (рецепторы) Гольджи
Рецепторы двигательных систем • Мышечные веретена • Сухожильные органы (рецепторы) Гольджи
 Сухожильные рецепторы Гольджи • Рецепторы Гольджи являются датчиками силы сокращения мышцы При их активации запускается рефлекс, тормозящий дальнейшую активацию данной мышцы
Сухожильные рецепторы Гольджи • Рецепторы Гольджи являются датчиками силы сокращения мышцы При их активации запускается рефлекс, тормозящий дальнейшую активацию данной мышцы
 Мышечное веретено – датчик длины мышцы При укорочении мышцы частота ПД от мышечного веретена уменьшается, Генерация ПД может прекратиться. Figure 13 -5: Gamma motor neurons
Мышечное веретено – датчик длины мышцы При укорочении мышцы частота ПД от мышечного веретена уменьшается, Генерация ПД может прекратиться. Figure 13 -5: Gamma motor neurons
 Рефлекс с рецепторов Гольджи Осуществляются при увеличении напряжения мышцы Адресуется группе мышц-антагонистов Приводит к расслаблению мышцы, с сухожилия которой начался рефлекс и сокращение мышцыантагониста.
Рефлекс с рецепторов Гольджи Осуществляются при увеличении напряжения мышцы Адресуется группе мышц-антагонистов Приводит к расслаблению мышцы, с сухожилия которой начался рефлекс и сокращение мышцыантагониста.
 Мышечные веретена • Сенсорная иннервация – аннулоспиральные окончания в средней части мышечного веретена. • Двигательная иннервация – γ-волокна от γ-мотонейронов заканчиваются на сократительных частях мышечного веретена.
Мышечные веретена • Сенсорная иннервация – аннулоспиральные окончания в средней части мышечного веретена. • Двигательная иннервация – γ-волокна от γ-мотонейронов заканчиваются на сократительных частях мышечного веретена.
 Сухожильный рефлекс и рефлекс реципрокного торможения мышцы-антагониста
Сухожильный рефлекс и рефлекс реципрокного торможения мышцы-антагониста
 Формы координации рефлексов Координация – оптимальное взаимоотношение центров, направленное на достижение полезного результата. 1. ОТНОСИТЕЛЬНАЯ РЕФЛЕКСОВ. 2. СИНЕРГИЗМ. 3. АНТАГОНИЗМ. 4. ДОМИНАНТА. НЕЗАВИСИМОСТЬ
Формы координации рефлексов Координация – оптимальное взаимоотношение центров, направленное на достижение полезного результата. 1. ОТНОСИТЕЛЬНАЯ РЕФЛЕКСОВ. 2. СИНЕРГИЗМ. 3. АНТАГОНИЗМ. 4. ДОМИНАНТА. НЕЗАВИСИМОСТЬ
 Относительная независимость рефлексов
Относительная независимость рефлексов
 Синергизм Эмоциональное поведение
Синергизм Эмоциональное поведение
 Антагонизм Соматические рефлексы Динамический переход: антагонизм ↔ синергизм. Вегетативные рефлексы Жестко закреплённый антагонизм. 1. Рефлекс глотания и дыхания.
Антагонизм Соматические рефлексы Динамический переход: антагонизм ↔ синергизм. Вегетативные рефлексы Жестко закреплённый антагонизм. 1. Рефлекс глотания и дыхания.
 может состояться или не состояться,") Принцип вероятности При действии адекватного стимула рефлекторная реакция 1) может состояться или не состояться, 2) может быть различной по силе и времени возникновения.
Принцип вероятности При действии адекватного стимула рефлекторная реакция 1) может состояться или не состояться, 2) может быть различной по силе и времени возникновения.
 Строение сегмента спинного мозга
Строение сегмента спинного мозга
 спинномозговых нервов,") Строение спинномозгового нерва Отрезок спинного мозга, соответствующий каждой паре (правой и левой) спинномозговых нервов, образует сегмент спинного мозга. В шейном отделе они расположены на один позвонок выше соответствующего им по счету позвонка, в среднегрудном — выше на 2 позвонка, а нижнегрудном - выше на 3 позвонка (правило Шило). 1 - задний корешок; чувствительный; 2 - спинномозговой узел; 3 - передний корешок; двигательный; 4 - спинномозговой нерв или канатик - смешанный, выходит из позвоночного канала через межпозвоночное отверстие; 5 - ветвь мозговой оболочки - чувствительная, иннервирует оболочки мозга; 6 - задняя ветвь спинномозгового нерва - смешанная, иннервирует кожу и аутохтонные мышцы дорсальной поверхности туловища; 7 - передняя ветвь спинномозгового нерва - смешанная, иннервирует кожу и мышцы вентральной поверхности туловища; 8 - боковое промежуточное вегетативное ядро; симпатическое; 9 - белая соединительная ветвь; 10 - серая соединительная ветвь - обе соединительные ветви являются вегетативными симпатическими; 11 - вегетативный узел пограничного симпатического ствола.
Строение спинномозгового нерва Отрезок спинного мозга, соответствующий каждой паре (правой и левой) спинномозговых нервов, образует сегмент спинного мозга. В шейном отделе они расположены на один позвонок выше соответствующего им по счету позвонка, в среднегрудном — выше на 2 позвонка, а нижнегрудном - выше на 3 позвонка (правило Шило). 1 - задний корешок; чувствительный; 2 - спинномозговой узел; 3 - передний корешок; двигательный; 4 - спинномозговой нерв или канатик - смешанный, выходит из позвоночного канала через межпозвоночное отверстие; 5 - ветвь мозговой оболочки - чувствительная, иннервирует оболочки мозга; 6 - задняя ветвь спинномозгового нерва - смешанная, иннервирует кожу и аутохтонные мышцы дорсальной поверхности туловища; 7 - передняя ветвь спинномозгового нерва - смешанная, иннервирует кожу и мышцы вентральной поверхности туловища; 8 - боковое промежуточное вегетативное ядро; симпатическое; 9 - белая соединительная ветвь; 10 - серая соединительная ветвь - обе соединительные ветви являются вегетативными симпатическими; 11 - вегетативный узел пограничного симпатического ствола.
 Проводниковая организация спинного мозга • Аксоны спинальных ганглиев и серого вещества спинного мозга идут в его белое вещество, а затем в другие структуры ЦНС, создавая тем самым так называемые проводящие пути, функционально подразделяющиеся на проприоцептивные, спиноцеребральные (восходящие) и цереброспинальные (нисходящие).
Проводниковая организация спинного мозга • Аксоны спинальных ганглиев и серого вещества спинного мозга идут в его белое вещество, а затем в другие структуры ЦНС, создавая тем самым так называемые проводящие пути, функционально подразделяющиеся на проприоцептивные, спиноцеребральные (восходящие) и цереброспинальные (нисходящие).
 Проприоспинальные пути • связывают между собой нейроны одного или разных сегментов спинного мозга. • Функция таких связей ассоциативная и заключается в координации позы, тонуса мышц, движений различных метамеров тела. • Один метамер включает себя 1 пару спинномозговых нервов и участок тела иннервируемый им.
Проприоспинальные пути • связывают между собой нейроны одного или разных сегментов спинного мозга. • Функция таких связей ассоциативная и заключается в координации позы, тонуса мышц, движений различных метамеров тела. • Один метамер включает себя 1 пару спинномозговых нервов и участок тела иннервируемый им.
 Спиноцеребральные пути • соединяют сегменты спинного мозга со структурами головного мозга. • Они представлены проприоцептивным, спиноталамическим, спиномозжечковым и спиноретикулярным путями.
Спиноцеребральные пути • соединяют сегменты спинного мозга со структурами головного мозга. • Они представлены проприоцептивным, спиноталамическим, спиномозжечковым и спиноретикулярным путями.
 • начинается от рецепторов глубокой") Проприоцептивный путь (тонкий пучок Голля и клиновидный пучок Бурдаха) • начинается от рецепторов глубокой чувствительности надкостницы, оболочек суставов, сухожилий и мышц. • Через спинальный ганглий он идет в задние корешки спинного мозга, в белое вещество задних канатиков и, не переключаясь на новый нейрон на уровне спинного мозга, поднимается в ядра Голля и Бурдаха продолговатого мозга. • Здесь происходит переключение на новый нейрон, далее путь идет в латеральные ядра таламуса противоположного полушария мозга, здесь переключается на новый нейрон (второе переключение). • От таламуса путь поднимается к нейронам соматосенсорной коры. • По ходу волокна этих трактов отдают коллатерали в каждом сегменте спинного мозга, что создает возможность коррекции позы всего туловища.
Проприоцептивный путь (тонкий пучок Голля и клиновидный пучок Бурдаха) • начинается от рецепторов глубокой чувствительности надкостницы, оболочек суставов, сухожилий и мышц. • Через спинальный ганглий он идет в задние корешки спинного мозга, в белое вещество задних канатиков и, не переключаясь на новый нейрон на уровне спинного мозга, поднимается в ядра Голля и Бурдаха продолговатого мозга. • Здесь происходит переключение на новый нейрон, далее путь идет в латеральные ядра таламуса противоположного полушария мозга, здесь переключается на новый нейрон (второе переключение). • От таламуса путь поднимается к нейронам соматосенсорной коры. • По ходу волокна этих трактов отдают коллатерали в каждом сегменте спинного мозга, что создает возможность коррекции позы всего туловища.
 Спиноталамический путь • начинается от болевых, температурных, барорецепторов кожи. • Сигнал от рецепторов кожи идет в спинальный ганглий, далее через задний корешок к заднему рогу спинного мозга, здесь переключается на новый нейрон (первое переключение). • Чувствительные нейроны задних рогов посылают аксоны на противоположную сторону спинного мозга и поднимаются по боковому канатику к таламусу. • Здесь происходит второе переключение и поднимаются в сенсорную кору. • Часть волокон кожных рецепторов идет к таламусу по переднему канатику спинного мозга.
Спиноталамический путь • начинается от болевых, температурных, барорецепторов кожи. • Сигнал от рецепторов кожи идет в спинальный ганглий, далее через задний корешок к заднему рогу спинного мозга, здесь переключается на новый нейрон (первое переключение). • Чувствительные нейроны задних рогов посылают аксоны на противоположную сторону спинного мозга и поднимаются по боковому канатику к таламусу. • Здесь происходит второе переключение и поднимаются в сенсорную кору. • Часть волокон кожных рецепторов идет к таламусу по переднему канатику спинного мозга.
 Спиномозжечковые пути • начинаются от рецепторов мышц, связок, внутренних органов и представлены неперекрещивающимся пучком Говерса и дважды перекрещивающимся пучком Флексига. • Следовательно, правый и левый мозжечок получают информацию только со своей стороны тела. • Эта информация идет от рецепторов Гольджи сухожилий, проприорецепторов, рецепторов давления и прикосновения. • Спиноретикулярный путь – начинается от интернейронов спинного мозга и доходит до РФ ствола мозга. Несет информацию от висцерорецепторов.
Спиномозжечковые пути • начинаются от рецепторов мышц, связок, внутренних органов и представлены неперекрещивающимся пучком Говерса и дважды перекрещивающимся пучком Флексига. • Следовательно, правый и левый мозжечок получают информацию только со своей стороны тела. • Эта информация идет от рецепторов Гольджи сухожилий, проприорецепторов, рецепторов давления и прикосновения. • Спиноретикулярный путь – начинается от интернейронов спинного мозга и доходит до РФ ствола мозга. Несет информацию от висцерорецепторов.
 Цереброспинальные пути • начинаются от нейронов структур головного мозга и заканчиваются на нейронах сегментов спинного мозга. • К ним относятся пути: • кортикоспинальный путь, обеспечивающий регуляцию произвольных движений, • руброспинальный, • вестибулоспинальный и • ретикулоспинальный пути, регулирующие тонус мускулатуры. • Объединяющим для этих путей является то, что конечным пунктом для них являются мотонейроны передних рогов спинного мозга.
Цереброспинальные пути • начинаются от нейронов структур головного мозга и заканчиваются на нейронах сегментов спинного мозга. • К ним относятся пути: • кортикоспинальный путь, обеспечивающий регуляцию произвольных движений, • руброспинальный, • вестибулоспинальный и • ретикулоспинальный пути, регулирующие тонус мускулатуры. • Объединяющим для этих путей является то, что конечным пунктом для них являются мотонейроны передних рогов спинного мозга.
 • возникает после полной перерезки спинного мозга. • Он заключается в") Спинальный шок (шок-удар) • возникает после полной перерезки спинного мозга. • Он заключается в том, что все центры ниже перерезки перестают организовывать присущие им рефлексы. • Спинальный шок характеризуется временным исчезновением рефлекторных функций спинного мозга. • Нарушение рефлекторной деятельности после пересечения спинного мозга у разных животных длится разное время. • У обезьян первые признаки восстановления рефлексов после перерезки спинного мозга появляются через несколько суток; у лягушки – через минуты, у человека первые спинальные рефлексы восстанавливаются через несколько недель, а то и месяцев.
Спинальный шок (шок-удар) • возникает после полной перерезки спинного мозга. • Он заключается в том, что все центры ниже перерезки перестают организовывать присущие им рефлексы. • Спинальный шок характеризуется временным исчезновением рефлекторных функций спинного мозга. • Нарушение рефлекторной деятельности после пересечения спинного мозга у разных животных длится разное время. • У обезьян первые признаки восстановления рефлексов после перерезки спинного мозга появляются через несколько суток; у лягушки – через минуты, у человека первые спинальные рефлексы восстанавливаются через несколько недель, а то и месяцев.
 Причина шока • нарушение регуляции рефлексов со стороны вышележащих структур ЦНС. • При травме спинного мозга у человека может появиться группа двигательных спинальных рефлексов, которые имеются в норме лишь в первые дни и месяцы постнатального развития. • Растормаживание этих примитивных рефлексов является клиническим признаком нарушений работы спинного мозга.
Причина шока • нарушение регуляции рефлексов со стороны вышележащих структур ЦНС. • При травме спинного мозга у человека может появиться группа двигательных спинальных рефлексов, которые имеются в норме лишь в первые дни и месяцы постнатального развития. • Растормаживание этих примитивных рефлексов является клиническим признаком нарушений работы спинного мозга.
 Спинальное животное • это животное, у которого спинной мозг отделен от головного мозга, перерезка спинного мозга производится ниже 3 -го шейного позвонка. • Перерезка выше 3 -го шейного позвонка несовместима с жизнью, потому что на уровне 1 -2 шейных позвонков лежат нервные центры дыхательной мускулатуры и, если их разрушить, животное погибнет от паралича дыхательных мышц, т. е. асфиксии.
Спинальное животное • это животное, у которого спинной мозг отделен от головного мозга, перерезка спинного мозга производится ниже 3 -го шейного позвонка. • Перерезка выше 3 -го шейного позвонка несовместима с жизнью, потому что на уровне 1 -2 шейных позвонков лежат нервные центры дыхательной мускулатуры и, если их разрушить, животное погибнет от паралича дыхательных мышц, т. е. асфиксии.
 Повреждение спинного мозга у человека • • При травмах у человека в ряде случаев происходит полное или половинное пересечение спинного мозга. При половинном латеральном повреждении спинного мозга развивается синдром Броун-Секара - на половине поражения (ниже места поражения) развивается паралич двигательной системы вследствие повреждения пирамидных путей. На противоположной стороне движения сохраняются. На стороне поражения (ниже места поражения) нарушается проприоцептивная чувствительность (от рецепторов глубокой чувствительности надкостницы, оболочек суставов, сухожилий и мышц). Это обусловлено тем, что восходящие пути глубокой чувствительности идут по своей стороне спинного мозга до продолговатого мозга, где происходит их перекрест (пучок Голля и Бурдаха). На противоположной стороне туловища (относительно повреждения) нарушается болевая и температурная чувствительность (спиноталамический путь), т. к. восходящие пути глубокой чувствительности идут от спинального ганглия в задний рог спинного мозга, где переключаются на новый нейрон, аксон которого переходит на противоположную сторону. В итоге, если повреждена левая половина спинного мозга, то исчезает болевая и температурная чувствительность правой половины туловища ниже повреждения. После травмы спинного мозга у человека наблюдается извращение спинальных рефлексов: ослабление миотатических и кожно-мышечных двигательных рефлексов, усиление сухожильных рефлексов, извращение подошвенного рефлекса.
Повреждение спинного мозга у человека • • При травмах у человека в ряде случаев происходит полное или половинное пересечение спинного мозга. При половинном латеральном повреждении спинного мозга развивается синдром Броун-Секара - на половине поражения (ниже места поражения) развивается паралич двигательной системы вследствие повреждения пирамидных путей. На противоположной стороне движения сохраняются. На стороне поражения (ниже места поражения) нарушается проприоцептивная чувствительность (от рецепторов глубокой чувствительности надкостницы, оболочек суставов, сухожилий и мышц). Это обусловлено тем, что восходящие пути глубокой чувствительности идут по своей стороне спинного мозга до продолговатого мозга, где происходит их перекрест (пучок Голля и Бурдаха). На противоположной стороне туловища (относительно повреждения) нарушается болевая и температурная чувствительность (спиноталамический путь), т. к. восходящие пути глубокой чувствительности идут от спинального ганглия в задний рог спинного мозга, где переключаются на новый нейрон, аксон которого переходит на противоположную сторону. В итоге, если повреждена левая половина спинного мозга, то исчезает болевая и температурная чувствительность правой половины туловища ниже повреждения. После травмы спинного мозга у человека наблюдается извращение спинальных рефлексов: ослабление миотатических и кожно-мышечных двигательных рефлексов, усиление сухожильных рефлексов, извращение подошвенного рефлекса.
 изображена на рис. и подпишите структуры, обозначенные") Определите, дуга какого рефлекса (соматического или вегетативного) изображена на рис. и подпишите структуры, обозначенные цифрами • А • Б
Определите, дуга какого рефлекса (соматического или вегетативного) изображена на рис. и подпишите структуры, обозначенные цифрами • А • Б
 Спасибо за внимание
Спасибо за внимание