Молекулярная биология ч.4 Мессенджеры.pptx
- Количество слайдов: 25



Постсинаптическая трансформация сигнала: первичные и вторичные посредники, G-белки и протеинкиназы. Молекулярные механизмы передачи импульса в мембранах нейронов, ионные каналы и рецепторы.








, нейромедиатора и гормона, известного под названием серотонин, взаимодействующие")
мембранные рецепторы 5 -гидрокситриптамина (5 -HT), нейромедиатора и гормона, известного под названием серотонин, взаимодействующие также с множеством медицинских препаратов и психоактивных веществ. Активация рецепторов запускает внутриклеточные процессы, влияющие на активность других медиаторных систем — глутаматной, дофаминовой и ГАМК.






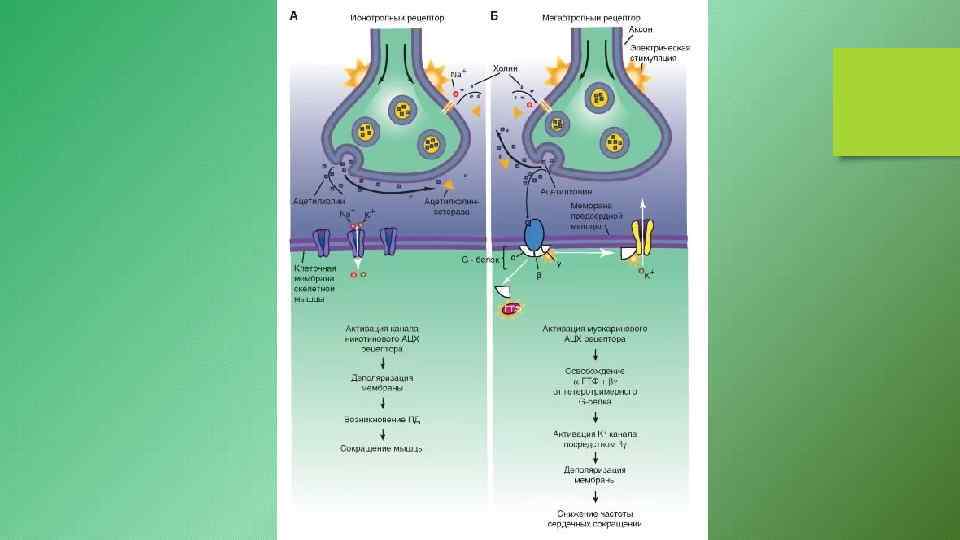


1. Связывание агониста индуцирует открывание канала, образуемого рецептором. Примерами могут служить никотиновый ацетилхолиновый рецептор (n. ACh. R), рецептор γ – аминомасляной кислоты (ГАМК) и глициновый рецептор. Все эти рецепторы нейромедиаторов, которые структурно объединены в малое суперсемейство. 2. При связывании лиганда активируется тирозиновая протеинкиназа, представляющая собой цитоплазматический домен рецептора. Обычно сам рецептор и является мишенью (происходит аутофосфорилирование), но данные о том, какие именно белки фосфорилируются и как они влияют на клетку, практически отсутствуют. Этот механизм используют рецепторы митогенных пептидных гормонов и фактора роста, причём многие из этих рецепторов структурно родственны между собой. Каждый из этих рецепторных полипептидов имеет по одному трансмембранному сегменту. 3. Рецептор образует комплекс с одной из групп мембраносвязанных GTP– связывающих белков, называемых G– белками. При связывании лиганда с рецептором в комплексе рецептор/G– белок происходит конформационное изменение, в результате чего облегчается обмен связанного GDP и GTP на G– белке. G– белок активируется на короткое время, будучи связанным с GTP, и в этом состоянии может отделиться от рецептора, причём одна или больше субъединиц G– белка могут связаться с другими мембранными белками. Это так называемая «мишень» . На них и оказывается воздействие. К этим мишеням относятся ионные каналы, аденилатциклаза, с. GMP– фосфодиэстераза и фосфолипаза С. К суперсемейству рецепторов, чьё действие опосредуется G– белками, принадлежат также β – адренергические рецепторы, мускариновые ацетилхолиновые рецепторы и опсины (световые рецепторы). Каждый из этих рецепторов имеет семь трансмембранных сегментов.
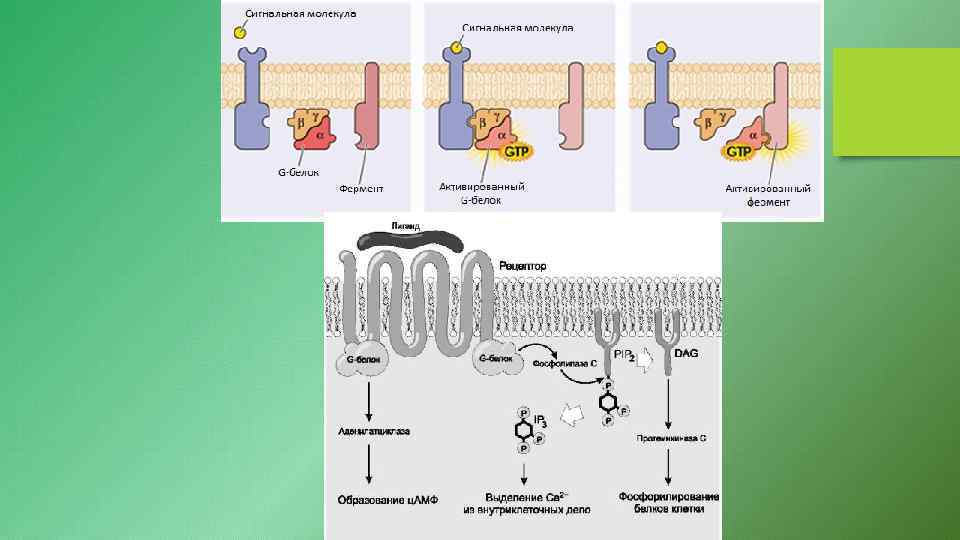

Примеры молекулярной организации некоторых структур, участвующих в путях сигнальной трансдукции. А - рецептор мембраны клетки, связывающий на внешней поверхности лиганд, а внутри гетеротримерный G-белок. Б - молекулярная организация аденилатциклазы. В - структура фосфатидилинози- тол-4, 5 -дифосфата и образованных под действием фосфолипазы С инозитол-1, 4, 5 трифосфата и диацилглицерола. Г - структура 3', 5'циклического АМФ (активатора протеинкиназы А). Д структура ГМФ. Е - структура 3', 5'-циклического ГМФ (активатора протеинкиназы G)
 - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной")
G-белки (ГТФ-связывающие белки) - универсальные посредники при передаче сигналов от рецепторов к ферментам клеточной мембраны, катализирующим образование вторичных посредников гормонального сигнала. G-белки - олигомеры, состоящие из α, β и γсубъединиц. Состав димеров βγ незначительно различаются в разных тканях, но в пределах одной клетки все G-белки, как правило, имеют одинаковый комплект βγ-субъединиц. Поэтому G-белки принято различать по их α-субъединицам. Выявлено 16 генов, кодирующих различные α-субъединицы G-белков. Некоторые из генов имеют более одного белка, вследствие альтернативного сплайсинга РНК. Каждая а-субъединица в составе G-белка имеет специфические центры: связывания ГТФ или ГДФ; взаимодействия с рецептором; связывания с βγ-субъединицами; фосфорилирования под действием протеинкиназы С; взаимодействия с ферментом аденилатциклазой или фосфолипазой С. В структуре G-белков отсутствуют α-спиральные, пронизывающие мембрану домены. G-белки относят к группе "заякоренных" белков.
 Принцип работы мономерных ГТФ-связывающих белков (мономерных G-белков).")
Принцип работы гетеротримерных ГТФсвязывающих белков (гетеротримерных G-белков) Принцип работы мономерных ГТФ-связывающих белков (мономерных G-белков).


Молекулярная биология ч.4 Мессенджеры.pptx