

6429f6eed48d695f9bf25ae2176b0d47.ppt
- Количество слайдов: 72
 Популяционная генетика - 5
Популяционная генетика - 5
") Факторы эволюции • мутации • отбор • половое размножение • генетико-автоматические процессы (генетический дрейф) • миграция
Факторы эволюции • мутации • отбор • половое размножение • генетико-автоматические процессы (генетический дрейф) • миграция
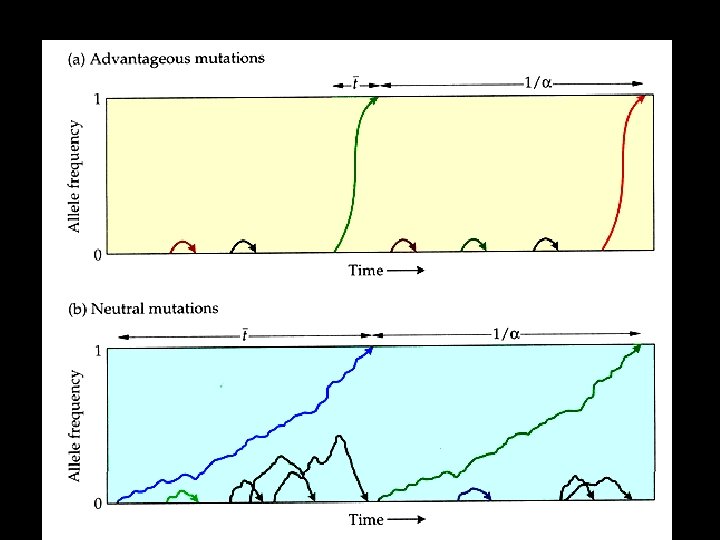
 Как отличить Дарвина от Кимуры?
Как отличить Дарвина от Кимуры?
 kотбор = Nus kнейт. = u
kотбор = Nus kнейт. = u
 kотбор = Nus kнейт. = u Преобладает отбор Преобладает дрейф s > 1/N s < 1/N
kотбор = Nus kнейт. = u Преобладает отбор Преобладает дрейф s > 1/N s < 1/N
 Положительный отбор
Положительный отбор
 Эпизодические адаптации
Эпизодические адаптации

 Dважн/Dнейтр = ?
Dважн/Dнейтр = ?
 human rat mouse
human rat mouse
 ожидаем наблюдаем 25% : 50% : 33% : 37% : 25% 30% Bazykin et al. Nature 2004
ожидаем наблюдаем 25% : 50% : 33% : 37% : 25% 30% Bazykin et al. Nature 2004
 взаимодействие между кодонами доля замен, происходящих в 0. 5 разных линиях 0. 4 0 100 расстояние, пар нуклеотидов Bazykin et al. Nature 2004
взаимодействие между кодонами доля замен, происходящих в 0. 5 разных линиях 0. 4 0 100 расстояние, пар нуклеотидов Bazykin et al. Nature 2004
 rat
rat
 rat
rat
 rat
rat
 rat
rat
 повсеместность «кластеров» Pseudomonas Muridae Pyrococcus Bacillus Salmonella Buchnera ожидание Chlamydia Escherichia Staphylococcus Streptococcus Helicobacter Vibrio Bazykin et al. Nature 2004
повсеместность «кластеров» Pseudomonas Muridae Pyrococcus Bacillus Salmonella Buchnera ожидание Chlamydia Escherichia Staphylococcus Streptococcus Helicobacter Vibrio Bazykin et al. Nature 2004
 В каких участках белка положительный отбор чаще и сильнее – в важных или в неважных? положительный отбор: Dважная > Dнейтр отрицательный отбор: Dважная < Dнейтр
В каких участках белка положительный отбор чаще и сильнее – в важных или в неважных? положительный отбор: Dважная > Dнейтр отрицательный отбор: Dважная < Dнейтр
 100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence mel-pse-vir in 21 bp window Bazykin et al. submitted
100% 90% 80% 70% 60% 50% 40% 30% 20% 10% 0% 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence mel-pse-vir in 21 bp window Bazykin et al. submitted
 Эволюция патогенов • ВИЧ-1 • грипп А
Эволюция патогенов • ВИЧ-1 • грипп А
 ВИЧ-1 • мало генов, сотни последовательностей • модельная система для изучения эволюции • основной подход: Dn/Ds по позициям образец 1 образец 2 … образец N ATG TCC CTA TAC GGA GCT ATG TCT CAT TAT AGA GAT ACG TCT CTT TAT AGG GCT Dn 1 Ds 1 Dn 2 Ds 2 Dn 3 Ds 3 Dn 4 Ds 4 Dn 5 Ds 5 Dn 6 Ds 6
ВИЧ-1 • мало генов, сотни последовательностей • модельная система для изучения эволюции • основной подход: Dn/Ds по позициям образец 1 образец 2 … образец N ATG TCC CTA TAC GGA GCT ATG TCT CAT TAT AGA GAT ACG TCT CTT TAT AGG GCT Dn 1 Ds 1 Dn 2 Ds 2 Dn 3 Ds 3 Dn 4 Ds 4 Dn 5 Ds 5 Dn 6 Ds 6
 Positive selection mapping of HIV-1 protease from 40, 000 patient samples. The Ka/Ks value represents the greatest selection pressure among all the individual amino acid mutations at each codon. The dotted line indicates the Ka/Ks value of 1 (Ka = d. N, Ks = d. S)
Positive selection mapping of HIV-1 protease from 40, 000 patient samples. The Ka/Ks value represents the greatest selection pressure among all the individual amino acid mutations at each codon. The dotted line indicates the Ka/Ks value of 1 (Ka = d. N, Ks = d. S)
 Bazykin et al. PNAS 2006") Отношение числа несинонимичных к числу синонимичных замен (Dn/Ds) Bazykin et al. PNAS 2006
Отношение числа несинонимичных к числу синонимичных замен (Dn/Ds) Bazykin et al. PNAS 2006
 ВИЧ-1 • мало генов, сотни последовательностей • модельная система для изучения эволюции • основной подход: Dn/Ds по позициям • … хорошо работает для поиска отбора, действовавшего равномерно в течение длительного времени … • … но плохо – для поиска переменного отбора.
ВИЧ-1 • мало генов, сотни последовательностей • модельная система для изучения эволюции • основной подход: Dn/Ds по позициям • … хорошо работает для поиска отбора, действовавшего равномерно в течение длительного времени … • … но плохо – для поиска переменного отбора.
 Другой подход: Эпизоды положительного отбора должны вызывать ВРЕМЕННОЕ увеличение скоростей замен – «пулеметные очереди» замен (множественные замены в быстрой последовательности)
Другой подход: Эпизоды положительного отбора должны вызывать ВРЕМЕННОЕ увеличение скоростей замен – «пулеметные очереди» замен (множественные замены в быстрой последовательности)
 AAC AAA ATC AAC GGG GCC 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
AAC AAA ATC AAC GGG GCC 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
 Пары замен 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
Пары замен 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
 Пары замен 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
Пары замен 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
 Расстояние между заменами в паре 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
Расстояние между заменами в паре 0. 1 нук. замен на сайт Bazykin et al. PNAS 2006
 Расстояние между заменами в паре несинонимичные расстояние между последовательными синонимичными заменами приблизительно соответствует ожиданию последовательные несинонимичные замены идут «очередями»
Расстояние между заменами в паре несинонимичные расстояние между последовательными синонимичными заменами приблизительно соответствует ожиданию последовательные несинонимичные замены идут «очередями»
 Расстояние между заменами в паре несинонимичные «сосредоточение» сильнее выражено в консервативных сайтах Bazykin et al. PNAS 2006
Расстояние между заменами в паре несинонимичные «сосредоточение» сильнее выражено в консервативных сайтах Bazykin et al. PNAS 2006
 Консервативные сайты под эпизодическим положительным отбором") Сайты под положительным отбором (Yamaguchi-Kabata and Gojobori, 2000) Консервативные сайты под эпизодическим положительным отбором Bazykin et al. PNAS 2006
Сайты под положительным отбором (Yamaguchi-Kabata and Gojobori, 2000) Консервативные сайты под эпизодическим положительным отбором Bazykin et al. PNAS 2006
 and synonymous (Lower) substitutions on the phylogenetic trees. The") Distances between successive nonsynonymous (Upper) and synonymous (Lower) substitutions on the phylogenetic trees. The mean distance between successive nonreversing substitutions is shown for codon sites with each total number of substitutions on the tree (dots). Solid lines present mean distances between successive substitutions within each sliding 30 -site window. Dashed lines show mean distances between independent substitutions obtained in simulations. Bazykin et al. PNAS 2006
Distances between successive nonsynonymous (Upper) and synonymous (Lower) substitutions on the phylogenetic trees. The mean distance between successive nonreversing substitutions is shown for codon sites with each total number of substitutions on the tree (dots). Solid lines present mean distances between successive substitutions within each sliding 30 -site window. Dashed lines show mean distances between independent substitutions obtained in simulations. Bazykin et al. PNAS 2006
 dn/ds тест для отдельных ветвей филогенетического дерева Bazykin et al. PNAS 2006
dn/ds тест для отдельных ветвей филогенетического дерева Bazykin et al. PNAS 2006
 Grenfell et al. Science 2004
Grenfell et al. Science 2004
 Bush et al. Science 1999
Bush et al. Science 1999
 Plotkin et al. PNAS 2000
Plotkin et al. PNAS 2000

 Тест Макдональда-Крейтмана
Тест Макдональда-Крейтмана
 kполезная ~ 2 Nus kнейтральная ~ u
kполезная ~ 2 Nus kнейтральная ~ u
 Bazykin et al. Biology Direct 2007
Bazykin et al. Biology Direct 2007

 Тест Макдональда-Крейтмана Основное объяснение: • движущий отбор Альтернативное объяснение: • усиление отрицательного отбора (напр. из-за увеличения размера популяции) этот тест чувствителен к особенностям популяционной демографии
Тест Макдональда-Крейтмана Основное объяснение: • движущий отбор Альтернативное объяснение: • усиление отрицательного отбора (напр. из-за увеличения размера популяции) этот тест чувствителен к особенностям популяционной демографии
 Simpson’s paradox Shapiro et al. PNAS 2007
Simpson’s paradox Shapiro et al. PNAS 2007
 0. 2 0. 18 0. 16 0. 14 0. 12 ratio 0. 1 0. 08 0. 06 0. 04 0. 02 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window Dn/Ds
0. 2 0. 18 0. 16 0. 14 0. 12 ratio 0. 1 0. 08 0. 06 0. 04 0. 02 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window Dn/Ds
 0. 2 0. 18 0. 16 0. 14 0. 12 ratio 0. 1 0. 08 0. 06 0. 04 0. 02 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window Dn/Ds Pn/Ps (with singletons)
0. 2 0. 18 0. 16 0. 14 0. 12 ratio 0. 1 0. 08 0. 06 0. 04 0. 02 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window Dn/Ds Pn/Ps (with singletons)
 1 0. 2 0. 18 0. 16 0. 7 0. 14 0. 6 0. 12 0. 5 0. 1 0. 4 0. 08 0. 3 0. 06 0. 2 0. 04 0. 1 0. 02 0 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window ratio 0. 9 (with singletons) Dn/Ds Pn/Ps (with singletons)
1 0. 2 0. 18 0. 16 0. 7 0. 14 0. 6 0. 12 0. 5 0. 1 0. 4 0. 08 0. 3 0. 06 0. 2 0. 04 0. 1 0. 02 0 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window ratio 0. 9 (with singletons) Dn/Ds Pn/Ps (with singletons)
 Smith and Eyre-Walker 2002: 45% Bierne and Eyre-Walker 2004: 25% Shapiro et al. 2007: 30% 1 0. 2 0. 18 0. 16 0. 7 0. 14 0. 6 0. 12 0. 5 0. 1 0. 4 0. 08 0. 3 0. 06 0. 2 0. 04 0. 1 0. 02 0 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window ratio 0. 9 (with singletons) Dn/Ds Pn/Ps (with singletons)
Smith and Eyre-Walker 2002: 45% Bierne and Eyre-Walker 2004: 25% Shapiro et al. 2007: 30% 1 0. 2 0. 18 0. 16 0. 7 0. 14 0. 6 0. 12 0. 5 0. 1 0. 4 0. 08 0. 3 0. 06 0. 2 0. 04 0. 1 0. 02 0 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window ratio 0. 9 (with singletons) Dn/Ds Pn/Ps (with singletons)
 1 0. 2 0. 18 0. 16 0. 7 0. 14 0. 6 0. 12 0. 5 0. 1 0. 4 0. 08 0. 3 0. 06 0. 2 0. 04 0. 1 0. 02 0 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window ratio 0. 9 (with singletons) Dn/Ds Pn/Ps (no singletons) Pn/Ps (with singletons)
1 0. 2 0. 18 0. 16 0. 7 0. 14 0. 6 0. 12 0. 5 0. 1 0. 4 0. 08 0. 3 0. 06 0. 2 0. 04 0. 1 0. 02 0 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window ratio 0. 9 (with singletons) Dn/Ds Pn/Ps (no singletons) Pn/Ps (with singletons)
 1 0. 2 0. 18 0. 16 0. 7 0. 14 0. 6 0. 12 0. 5 0. 1 0. 4 0. 08 0. 3 0. 06 0. 2 0. 04 0. 1 (no singletons) (with singletons) Dn/Ds Pn/Ps (no singletons) Pn/Ps (with singletons) 0. 02 0 ratio 0. 9 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window Bazykin et al. submitted
1 0. 2 0. 18 0. 16 0. 7 0. 14 0. 6 0. 12 0. 5 0. 1 0. 4 0. 08 0. 3 0. 06 0. 2 0. 04 0. 1 (no singletons) (with singletons) Dn/Ds Pn/Ps (no singletons) Pn/Ps (with singletons) 0. 02 0 ratio 0. 9 0 0 1 2 3 -5 6 -8 9 -10 11 -15 16 -20 divergence yak-pse-vir in 21 bp window Bazykin et al. submitted
 Ometto et al. Mol. Biol. Evol. 2005
Ometto et al. Mol. Biol. Evol. 2005
 «Выметание» отбором
«Выметание» отбором
 «Выметание» отбором
«Выметание» отбором
 «Выметание» отбором
«Выметание» отбором
 «Выметание» отбором Вокруг сайта под отбором изменчивости не будет!
«Выметание» отбором Вокруг сайта под отбором изменчивости не будет!
 «Выметание» отбором Hamblin and Di Rienzo, 2000 Am. J. Hum. Genet. 66: 1669– 1679
«Выметание» отбором Hamblin and Di Rienzo, 2000 Am. J. Hum. Genet. 66: 1669– 1679
 «Выметание» отбором
«Выметание» отбором
 «Выметание» отбором
«Выметание» отбором
 «Выметание» отбором
«Выметание» отбором
 «Выметание» отбором
«Выметание» отбором
 «Выметание» отбором Вокруг сайта под отбором изменчивость понижена
«Выметание» отбором Вокруг сайта под отбором изменчивость понижена
 Неравновесие по сцеплению Sabeti et al. , 2002 Nature 419: 832 -837
Неравновесие по сцеплению Sabeti et al. , 2002 Nature 419: 832 -837
 Насколько адаптивна эволюция?
Насколько адаптивна эволюция?
 kполезная ~ 2 Nus kнейтральная ~ u
kполезная ~ 2 Nus kнейтральная ~ u
 Bazykin et al. Biology Direct 2007
Bazykin et al. Biology Direct 2007
 Lynch et al. Science 2003
Lynch et al. Science 2003
 Lynch et al. PNAS 2007
Lynch et al. PNAS 2007
 Отбор: выводы • отрицательный отбор: самая распространенная форма отбора. Легко засечь. Под отбором ~10% нуклеотидов в геноме человека, ~50% нуклеотидов в геноме мухи. • положительный отбор: многие адаптации могут быть получены с помощью разных мутаций. Вероятность закрепления каждой такой мутации невелика, поэтому адаптивная эволюция непредсказуема. Доли замен под отбором и (тем более) силы отбора не знаем. • постоянный: отдельные истории • переменный: по-видимому, широко распространён
Отбор: выводы • отрицательный отбор: самая распространенная форма отбора. Легко засечь. Под отбором ~10% нуклеотидов в геноме человека, ~50% нуклеотидов в геноме мухи. • положительный отбор: многие адаптации могут быть получены с помощью разных мутаций. Вероятность закрепления каждой такой мутации невелика, поэтому адаптивная эволюция непредсказуема. Доли замен под отбором и (тем более) силы отбора не знаем. • постоянный: отдельные истории • переменный: по-видимому, широко распространён
 Общие выводы • эпистатические взаимодействия играют большую роль и не изучены • признаки, адаптивные сейчас, не обязательно были адаптивными при возникновении • в эволюции геномов огромную роль играют нейтральные (а не адаптивные) процессы
Общие выводы • эпистатические взаимодействия играют большую роль и не изучены • признаки, адаптивные сейчас, не обязательно были адаптивными при возникновении • в эволюции геномов огромную роль играют нейтральные (а не адаптивные) процессы
