

Лекция Обмен нуклеиновых кислот.ppt
- Количество слайдов: 52


 Переваривание нуклеотидов и всасывание продуктов их распада осуществляется в желудочно-кишечном тракте Под влиянием ферментов желудка и кишечника, частично НСl, нуклеопротеиды пищи распадаются на белок и белок нуклеиновые кислоты Распад нуклеиновых кислот осуществляется в основном гидролитическим путем в тонком кишечнике под действием ДНК-азы и РНК-азы поджелудочного сока. ДНК-азы РНК-азы Продуктами реакции при действии РНК-азы являются: • пиримидиновые мононуклеотиды • смесь ди- и тринуклеотидов • олигонуклеотиды Продуктами реакции при действии ДНК-азы являются: • динуклеотиды • олигонуклеотиды • небольшое количество мононуклеотидов
Переваривание нуклеотидов и всасывание продуктов их распада осуществляется в желудочно-кишечном тракте Под влиянием ферментов желудка и кишечника, частично НСl, нуклеопротеиды пищи распадаются на белок и белок нуклеиновые кислоты Распад нуклеиновых кислот осуществляется в основном гидролитическим путем в тонком кишечнике под действием ДНК-азы и РНК-азы поджелудочного сока. ДНК-азы РНК-азы Продуктами реакции при действии РНК-азы являются: • пиримидиновые мононуклеотиды • смесь ди- и тринуклеотидов • олигонуклеотиды Продуктами реакции при действии ДНК-азы являются: • динуклеотиды • олигонуклеотиды • небольшое количество мононуклеотидов
 Полный гидролиз нуклеиновых кислот осуществляется ферментами слизистой оболочки кишечника – фосфодиэстеразами Мононуклеотиды в кишечнике расщепляются под Мононуклеотиды действием неспецифических фосфатаз (кислой и фосфатаз щелочной). Частично распад осуществляется в клетках слизистой оболочки кишечника под действием нуклеотидаз Нуклеозиды могут быть использованы для синтеза нуклеиновых кислот организма. Если же происходит дальнейший распад, то гуанин не используется для синтетических целей. Аденин и урацил могут включаться в состав нуклеиновых кислот.
Полный гидролиз нуклеиновых кислот осуществляется ферментами слизистой оболочки кишечника – фосфодиэстеразами Мононуклеотиды в кишечнике расщепляются под Мононуклеотиды действием неспецифических фосфатаз (кислой и фосфатаз щелочной). Частично распад осуществляется в клетках слизистой оболочки кишечника под действием нуклеотидаз Нуклеозиды могут быть использованы для синтеза нуклеиновых кислот организма. Если же происходит дальнейший распад, то гуанин не используется для синтетических целей. Аденин и урацил могут включаться в состав нуклеиновых кислот.
 Таким образом, синтез нуклеиновых кислот определяется скоростью синтеза пуриновых и пиримидиновых нуклеотидов Источником рибозы и дезоксирибозы является глюкоза или ее метаболиты в глюкоза пентозофосфатном пути. Фосфорная кислота поступает достаточном количестве с пищей. в
Таким образом, синтез нуклеиновых кислот определяется скоростью синтеза пуриновых и пиримидиновых нуклеотидов Источником рибозы и дезоксирибозы является глюкоза или ее метаболиты в глюкоза пентозофосфатном пути. Фосфорная кислота поступает достаточном количестве с пищей. в
 Пути распада нуклеиновых кислот Полимерные молекулы нуклеиновых кислот расщепляются в тканях преимущественно гидролитическим путем при участии путем специфических ферментов – нуклеаз Ускоряют реакцию разрыва межнуклеотидных фосфодиэфирных связей в молекулах нуклеиновых связей кислот. Ферменты подкласса фосфодиэстераз
Пути распада нуклеиновых кислот Полимерные молекулы нуклеиновых кислот расщепляются в тканях преимущественно гидролитическим путем при участии путем специфических ферментов – нуклеаз Ускоряют реакцию разрыва межнуклеотидных фосфодиэфирных связей в молекулах нуклеиновых связей кислот. Ферменты подкласса фосфодиэстераз

 1. По месту действия Эндонуклеазы ü расщепляют внутренние межнуклеотидные связи в молекуле ДНК и РНК ü вызывают деполимеризацию нуклеиновых кислот и образование олигонуклеотидов q Экзонуклеазы ü вызывают последовательное гидролитическое отщепление концевых мононуклеотидов от ДНК, РНК или от олигонуклеотидов ü обеспечивают распад нуклеиновых кислот до свободных нуклеотидов. q
1. По месту действия Эндонуклеазы ü расщепляют внутренние межнуклеотидные связи в молекуле ДНК и РНК ü вызывают деполимеризацию нуклеиновых кислот и образование олигонуклеотидов q Экзонуклеазы ü вызывают последовательное гидролитическое отщепление концевых мононуклеотидов от ДНК, РНК или от олигонуклеотидов ü обеспечивают распад нуклеиновых кислот до свободных нуклеотидов. q
 ускоряют реакции распада межнуклеотидных связей в молекулах РНК;") 2. По специфичности действия Рибонуклеазы (РНК-азы) ускоряют реакции распада межнуклеотидных связей в молекулах РНК; q Дезоксирибонуклеазы (ДНК-азы) выполняют такую же функцию по отношению ДНК; q Неспецифические нуклеазы действуют и на РНК, и на ДНК. q
2. По специфичности действия Рибонуклеазы (РНК-азы) ускоряют реакции распада межнуклеотидных связей в молекулах РНК; q Дезоксирибонуклеазы (ДНК-азы) выполняют такую же функцию по отношению ДНК; q Неспецифические нуклеазы действуют и на РНК, и на ДНК. q
 3. По характеру действия на фосфодиэфирные связи 3’-нуклеазы – расщепляют связи межнуклеозидного фосфата с 5’-атомом углерода остатка рибозы или дезоксирибозы; q 5’-нуклеазы – расщепляют связи межнуклеозидного фосфата с 3’-атомом углерода остатка рибозы или дезоксирибозы. q
3. По характеру действия на фосфодиэфирные связи 3’-нуклеазы – расщепляют связи межнуклеозидного фосфата с 5’-атомом углерода остатка рибозы или дезоксирибозы; q 5’-нуклеазы – расщепляют связи межнуклеозидного фосфата с 3’-атомом углерода остатка рибозы или дезоксирибозы. q

 Дезоксирибонуклеазы I Ускоряют гидролиз фосфодиэфирных связей в одной гидролиз из цепей ДНК между остатком Н 3 РО 4 и 3’-атомом углерода остатка дезоксирибозы с образованием олигодезоксирибонуклеотидов Пример: панкреатическая ДНК-аза оптимум р. Н – 6, 8 -8, 0 активаторы – Mg 2+, Mn 2+ ингибиторы – олигонуклеотиды и анионы, связывающие Mg 2+ и Mn 2+
Дезоксирибонуклеазы I Ускоряют гидролиз фосфодиэфирных связей в одной гидролиз из цепей ДНК между остатком Н 3 РО 4 и 3’-атомом углерода остатка дезоксирибозы с образованием олигодезоксирибонуклеотидов Пример: панкреатическая ДНК-аза оптимум р. Н – 6, 8 -8, 0 активаторы – Mg 2+, Mn 2+ ингибиторы – олигонуклеотиды и анионы, связывающие Mg 2+ и Mn 2+
 Дезоксирибонуклеазы II Вызывают деполимеризацию молекулы ДНК в результате парных разрывов фосфодиэфирных связей обеих цепей с образованием более крупных олигонуклеотидов Пример: ДНК-аза селезенки оптимум р. Н – 4, 5 -5, 5 активаторы – Mg 2+ ингибиторы – SO 42–, РО 43–, As. О 43–, т. РНК
Дезоксирибонуклеазы II Вызывают деполимеризацию молекулы ДНК в результате парных разрывов фосфодиэфирных связей обеих цепей с образованием более крупных олигонуклеотидов Пример: ДНК-аза селезенки оптимум р. Н – 4, 5 -5, 5 активаторы – Mg 2+ ингибиторы – SO 42–, РО 43–, As. О 43–, т. РНК
 Экзодезоксирибонуклеазы Гидролизуют молекулы ДНК с Гидролизуют образованием дезокисрибонуклеозид 5’-фосфатов.
Экзодезоксирибонуклеазы Гидролизуют молекулы ДНК с Гидролизуют образованием дезокисрибонуклеозид 5’-фосфатов.
 ДНК в строго Расщепляют определенных зонах, имеющих") Рестриктазы ДНК-азы бактериального происхождения. Расщепляют чужеродные (фаговые) ДНК в строго Расщепляют определенных зонах, имеющих палиндромную зонах структуру, на ограниченное число фрагментов Рестриктазы применяются в генетической инженерии, так как из ДНК выщепляются фрагменты, инженерии которые затем встраиваются в бактериальную ДНК, принося бактерии новые биохимические признаки (например, способность синтезировать интерферон, инсулин и другие белки).
Рестриктазы ДНК-азы бактериального происхождения. Расщепляют чужеродные (фаговые) ДНК в строго Расщепляют определенных зонах, имеющих палиндромную зонах структуру, на ограниченное число фрагментов Рестриктазы применяются в генетической инженерии, так как из ДНК выщепляются фрагменты, инженерии которые затем встраиваются в бактериальную ДНК, принося бактерии новые биохимические признаки (например, способность синтезировать интерферон, инсулин и другие белки).
 Рибонуклеазы I Ускоряют реакцию гидролиза РНК по РНК пиримидиновым нуклеотидным остаткам.
Рибонуклеазы I Ускоряют реакцию гидролиза РНК по РНК пиримидиновым нуклеотидным остаткам.
 Гуанилрибонуклеазы Ускоряют гидролиз связей по 5’углеродному атому рибозы остатка рибозы гуаниловой кислоты и межнуклеотидного фосфата в молекуле РНК, образуя РНК гуанозин-3’-фосфат и олигонуклеотиды с концевым гуанозин-3’-фосфатом.
Гуанилрибонуклеазы Ускоряют гидролиз связей по 5’углеродному атому рибозы остатка рибозы гуаниловой кислоты и межнуклеотидного фосфата в молекуле РНК, образуя РНК гуанозин-3’-фосфат и олигонуклеотиды с концевым гуанозин-3’-фосфатом.
 Экзорибонуклеазы Ускоряют реакции отщепления рибонуклеотидов по концевым остаткам рибонуклеотидов РНК. Нуклеиновые кислоты дают при распаде смесь рибо- и дезоксирибонуклеозид-3’ и 5’-фосфатов
Экзорибонуклеазы Ускоряют реакции отщепления рибонуклеотидов по концевым остаткам рибонуклеотидов РНК. Нуклеиновые кислоты дают при распаде смесь рибо- и дезоксирибонуклеозид-3’ и 5’-фосфатов
 Полинуклеотидфосфорилаза Переносит нуклеотидные остатки с 3’-конца РНК на Н 3 РО 4 с образованием нуклеозиддифосфатов.
Полинуклеотидфосфорилаза Переносит нуклеотидные остатки с 3’-конца РНК на Н 3 РО 4 с образованием нуклеозиддифосфатов.
 Урацил-ДНК-гликозидаза Ускоряет отщепление остатка урацила поврежденной ДНК Урацил образуется при дезаминировании остатка цитозина: Образовавшийся разрыв служит сигналом для действия репарационной пиримидинспецифичной (уридин-специфичной) эндонуклеазы, расщепляющей фосфодиэфирную связь возле эндонуклеазы соответствующего участка поврждения После этого под действием ДНК-полимеразы и ДНК-лигазы ДНК-полимеразы ДНК-лигазы происходит заполнение бреши и восстановление исходной правильной структуры ДНК
Урацил-ДНК-гликозидаза Ускоряет отщепление остатка урацила поврежденной ДНК Урацил образуется при дезаминировании остатка цитозина: Образовавшийся разрыв служит сигналом для действия репарационной пиримидинспецифичной (уридин-специфичной) эндонуклеазы, расщепляющей фосфодиэфирную связь возле эндонуклеазы соответствующего участка поврждения После этого под действием ДНК-полимеразы и ДНК-лигазы ДНК-полимеразы ДНК-лигазы происходит заполнение бреши и восстановление исходной правильной структуры ДНК
 Гипоксантин-ДНК-гликозидаза Выполняет аналогичную функцию, но по отношению к гипоксантину, образующемуся из аденина: гипоксантину Эти процессы имеют существенное значение в репарации (восстановлении структуры) ДНК. Сходным образом происходит репарация ДНК, содержащей алкилированные азотистые основания и аналоги азотистых оснований.
Гипоксантин-ДНК-гликозидаза Выполняет аналогичную функцию, но по отношению к гипоксантину, образующемуся из аденина: гипоксантину Эти процессы имеют существенное значение в репарации (восстановлении структуры) ДНК. Сходным образом происходит репарация ДНК, содержащей алкилированные азотистые основания и аналоги азотистых оснований.

 + Н 2 О → нуклеозид + Н") Нуклеозидфосфаты подвергаются гидролизу: 1. нуклеотид (нуклеозидфосфат) + Н 2 О → нуклеозид + Н 3 РО 4 фермент 5’(или 3’)-нуклеотидаза 2. перенос остатка рибозы от нуклеотида на Н 3 РО 4: нуклеозид + Н 3 РО 4 → Р-1 -Ф + АО фермент рибозилтрансфераза или гидролиз: нуклеозид + Н 2 О → АО + рибоза фермент нуклеозидаза
Нуклеозидфосфаты подвергаются гидролизу: 1. нуклеотид (нуклеозидфосфат) + Н 2 О → нуклеозид + Н 3 РО 4 фермент 5’(или 3’)-нуклеотидаза 2. перенос остатка рибозы от нуклеотида на Н 3 РО 4: нуклеозид + Н 3 РО 4 → Р-1 -Ф + АО фермент рибозилтрансфераза или гидролиз: нуклеозид + Н 2 О → АО + рибоза фермент нуклеозидаза

 Конечным продуктом пуринового продуктом обмена является мочевая кислота 18– 20% распадается до СО 2 и NН 3 и 18– 20% выделяется через кишечник, остальное кишечник выводится с мочой ( в сутки до 400– 600 мг). Мочевая кислота антиоксидантом является
Конечным продуктом пуринового продуктом обмена является мочевая кислота 18– 20% распадается до СО 2 и NН 3 и 18– 20% выделяется через кишечник, остальное кишечник выводится с мочой ( в сутки до 400– 600 мг). Мочевая кислота антиоксидантом является
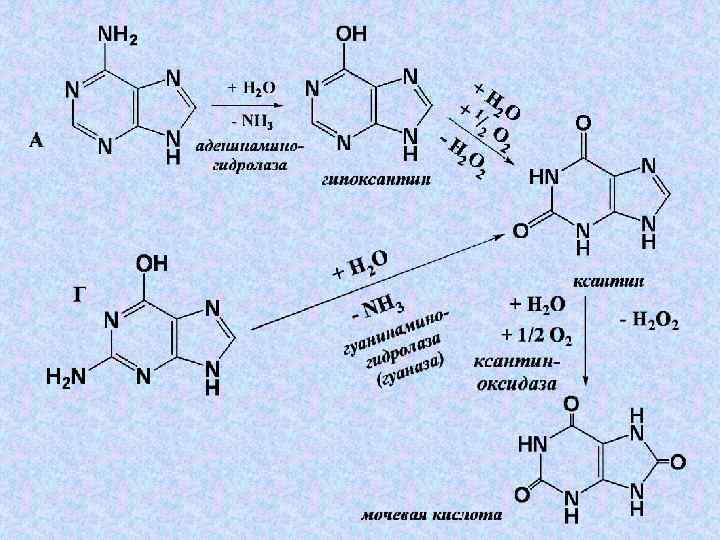
 и человека большинства растений может осуществляться дальнейшее") У ряда животных (но не у человека) и человека большинства растений может осуществляться дальнейшее окисление:
У ряда животных (но не у человека) и человека большинства растений может осуществляться дальнейшее окисление:

 Происходит в основном в печени Пиримидиновые основания подвергаются восстановлению: восстановлению
Происходит в основном в печени Пиримидиновые основания подвергаются восстановлению: восстановлению


 1. Образование карбамилфосфата из NН 3 и СО 2 при участии АТФ
1. Образование карбамилфосфата из NН 3 и СО 2 при участии АТФ
: фермент аспартаткарбамилтрансфераза") 2. Перенос остатка карбаминовой кислоты на аминогруппу аспарагиновой кислоты (первая специфическая реакция): фермент аспартаткарбамилтрансфераза
2. Перенос остатка карбаминовой кислоты на аминогруппу аспарагиновой кислоты (первая специфическая реакция): фермент аспартаткарбамилтрансфераза
 3. Образование дигидрооротовой кислоты фермент дигидрооротаза
3. Образование дигидрооротовой кислоты фермент дигидрооротаза
 4. Окисление дигидрооротовой кислоты фермент дигидрооротатдегидрогеназа
4. Окисление дигидрооротовой кислоты фермент дигидрооротатдегидрогеназа
 5. Образование оротидин-5’-фосфата фермент оротатфосфорибозилтрансфераза
5. Образование оротидин-5’-фосфата фермент оротатфосфорибозилтрансфераза
 Все эти реакции происходят в цитозоле клетки, а остальные цитозоле реакции – в митохондриях
Все эти реакции происходят в цитозоле клетки, а остальные цитозоле реакции – в митохондриях
 6. Декарбоксилирование оротидин-5’-фосфата О-5’-Ф → У-5’-Ф + СО 2 фермент О-5’-Ф-декарбоксилаза
6. Декарбоксилирование оротидин-5’-фосфата О-5’-Ф → У-5’-Ф + СО 2 фермент О-5’-Ф-декарбоксилаза
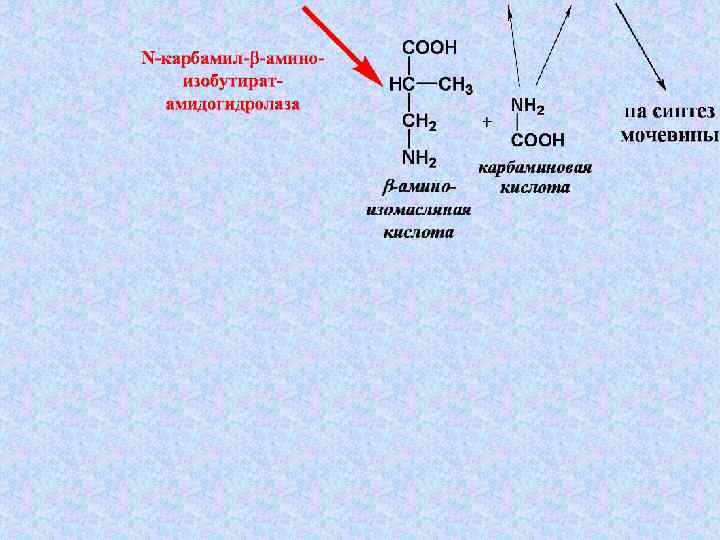
 7. Превращения У-5’-Ф в другие пиримидиновые нуклеотиды Путем реакций восстановления, аминирования и восстановления аминирования метилирования моно-, ди- и трифосфорных эфиров нуклеозидов. метилирования Трифосфорные эфиры образуются при взаимодействии нуклеозидмонофосфатов с АТФ: УМФ + АТФ → УДФ + АДФ УДФ + АТФ → УТФ + АДФ УТФ аминируется до ЦТФ : УТФ + АТФ + Глн ЦТФ + Глу + АДФ + Н 3 РО 4 фермент ЦТФ-синтаза Восстановление по остатку рибозы приводит к образованию дезоксирибонуклеотидов (фермент рибонуклеотидредуктаза). рибонуклеотидредуктаза Тимидиловые нуклеотиды образуются путем метилирования д. УМФ (фермент тимидилатсинтаза). тимидилатсинтаза
7. Превращения У-5’-Ф в другие пиримидиновые нуклеотиды Путем реакций восстановления, аминирования и восстановления аминирования метилирования моно-, ди- и трифосфорных эфиров нуклеозидов. метилирования Трифосфорные эфиры образуются при взаимодействии нуклеозидмонофосфатов с АТФ: УМФ + АТФ → УДФ + АДФ УДФ + АТФ → УТФ + АДФ УТФ аминируется до ЦТФ : УТФ + АТФ + Глн ЦТФ + Глу + АДФ + Н 3 РО 4 фермент ЦТФ-синтаза Восстановление по остатку рибозы приводит к образованию дезоксирибонуклеотидов (фермент рибонуклеотидредуктаза). рибонуклеотидредуктаза Тимидиловые нуклеотиды образуются путем метилирования д. УМФ (фермент тимидилатсинтаза). тимидилатсинтаза
 Синтез пиримидиновых нуклеотидов регулируется конечными продуктами
Синтез пиримидиновых нуклеотидов регулируется конечными продуктами

 1. Образование 5 -фосфорибозил-1 -пирофосфата Р-5 -Ф + АТФ → 5 -ФР-1 -ПФ + АМФ фермент пирофосфорилаза
1. Образование 5 -фосфорибозил-1 -пирофосфата Р-5 -Ф + АТФ → 5 -ФР-1 -ПФ + АМФ фермент пирофосфорилаза
 2. Образование 5 -фосфорибозиламина фермент ФРПФ-глутаминамидотрансфераза
2. Образование 5 -фосфорибозиламина фермент ФРПФ-глутаминамидотрансфераза
 3. Присоединение глицина фермент фосфорибозил-глицинамидсинтетаза
3. Присоединение глицина фермент фосфорибозил-глицинамидсинтетаза
 4. Преобразования 5 -фосфорибозилглицинамида
4. Преобразования 5 -фосфорибозилглицинамида
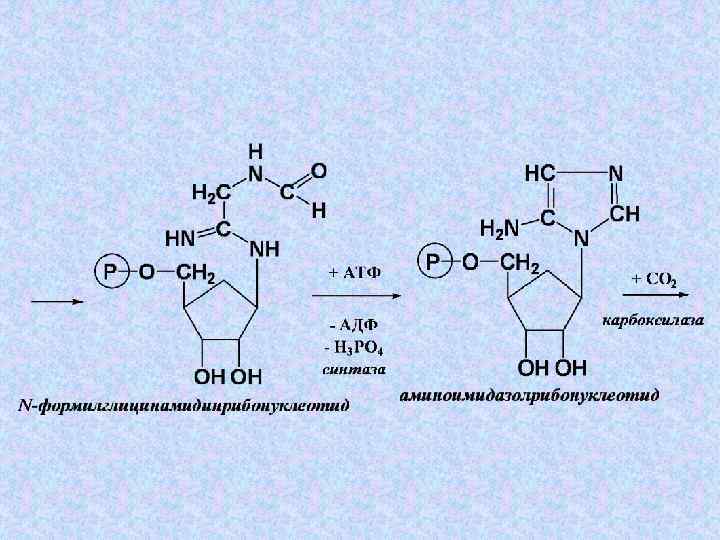
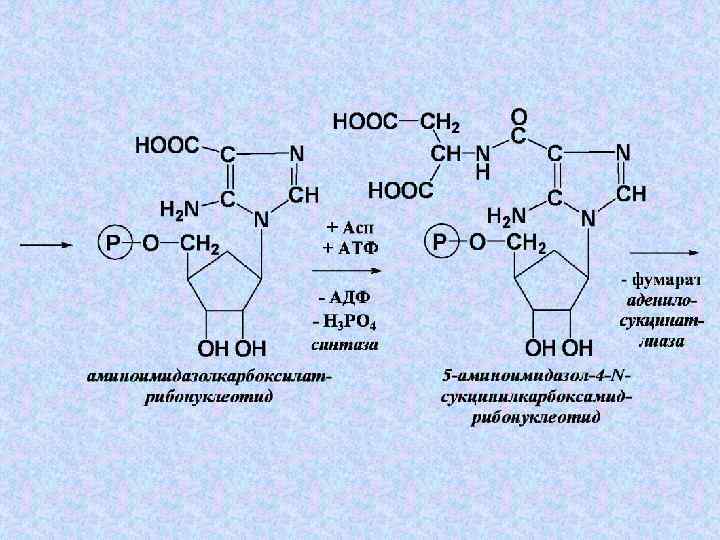
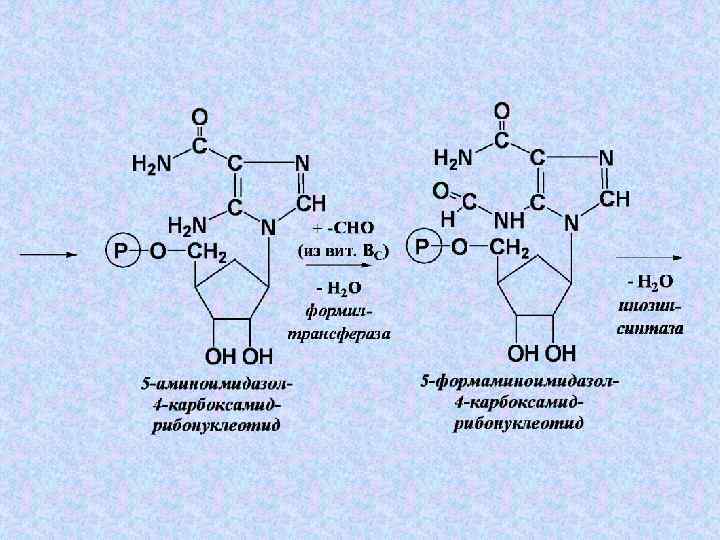
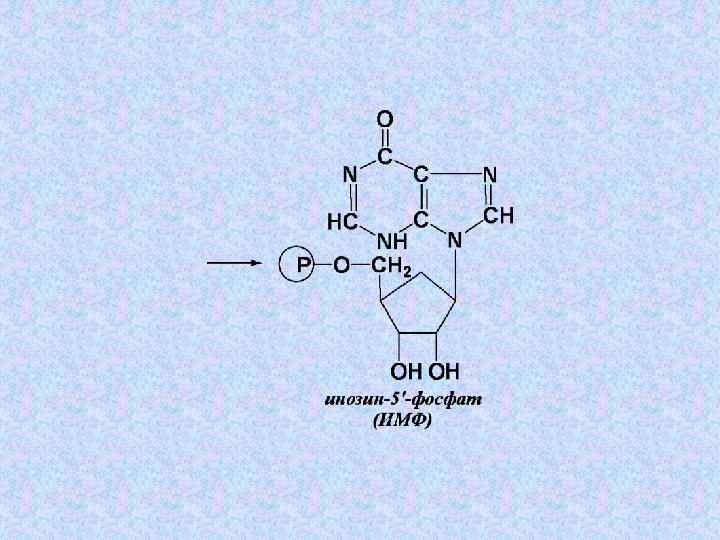
 + Глн + 3 АТФ") Суммарное уравнение этапа 4: 5 -фосфорибозилглицинамид + 2–СНО(Вит ВС) + Глн + 3 АТФ + СО 2 + Асп → → 2 Н 2 О + Глу + 3 АДФ + 3 Н 3 РО 4 + Фумарата + ИМФ
Суммарное уравнение этапа 4: 5 -фосфорибозилглицинамид + 2–СНО(Вит ВС) + Глн + 3 АТФ + СО 2 + Асп → → 2 Н 2 О + Глу + 3 АДФ + 3 Н 3 РО 4 + Фумарата + ИМФ
 Пуриновый цикл строится из очень простых веществ:
Пуриновый цикл строится из очень простых веществ: