

Лекция 6.2. Научение и память.ppt
- Количество слайдов: 57
 Память: Психологические и психофизиологические аспекты; нейрофизиологические механизмы
Память: Психологические и психофизиологические аспекты; нейрофизиологические механизмы
 Виды памяти Существует множество классификаций по самым разным основаниям, которые грубо можно поделить на две группы – «что запоминается» и «как запоминается» . В нейрофизиологии - элементарные механизмы научения и памяти: привыкание, сенситизация, временная связь. Эти формы памяти и научения называют простыми, чтобы отличать от научения, имеющего произвольный, осознанный характер. Генотипическая и фенотипическая. Первая — генотипическая, или филогенетическая, связана с безусловными рефлексами и инстинктами. Вторая — фенотипическая — обеспечивает обработку и хранение информации, приобретаемой в ходе онтогенеза на основе различных механизмов научения.
Виды памяти Существует множество классификаций по самым разным основаниям, которые грубо можно поделить на две группы – «что запоминается» и «как запоминается» . В нейрофизиологии - элементарные механизмы научения и памяти: привыкание, сенситизация, временная связь. Эти формы памяти и научения называют простыми, чтобы отличать от научения, имеющего произвольный, осознанный характер. Генотипическая и фенотипическая. Первая — генотипическая, или филогенетическая, связана с безусловными рефлексами и инстинктами. Вторая — фенотипическая — обеспечивает обработку и хранение информации, приобретаемой в ходе онтогенеза на основе различных механизмов научения.
 Биологическая память – способность живых существ, воспринимая воздействие извне, закреплять, сохранять и в последующем воспроизводить вызываемые этими воздействиями изменения функционального состояния и структуры. Генетическая память. Именно на неё опирается стабилизация органических систем и их структурное самовоспроизведение. Носителем генетической памяти являются нуклеиновые кислоты (ДНК, РНК), которые позволяют обеспечить стабильность хранения информации.
Биологическая память – способность живых существ, воспринимая воздействие извне, закреплять, сохранять и в последующем воспроизводить вызываемые этими воздействиями изменения функционального состояния и структуры. Генетическая память. Именно на неё опирается стабилизация органических систем и их структурное самовоспроизведение. Носителем генетической памяти являются нуклеиновые кислоты (ДНК, РНК), которые позволяют обеспечить стабильность хранения информации.
. Состоит в способности после первой встречи") Иммунологическая память (тесно связана с генетической памятью). Состоит в способности после первой встречи с генетически чужеродными телами и веществами узнавать их при повторной встрече, связывать и включать неспецифические механизмы их уничтожения. Чужеродные вещества – антигены, а иммунные белки, обладающие способность разрушать антигены, - антитела. Первая встреча с антигеном вызывает увеличение числа соответствующих лимфоцитов и их дифференцировку на эффекторные клетки и клетки памяти. Последние остаются в организме на всю жизнь.
Иммунологическая память (тесно связана с генетической памятью). Состоит в способности после первой встречи с генетически чужеродными телами и веществами узнавать их при повторной встрече, связывать и включать неспецифические механизмы их уничтожения. Чужеродные вещества – антигены, а иммунные белки, обладающие способность разрушать антигены, - антитела. Первая встреча с антигеном вызывает увеличение числа соответствующих лимфоцитов и их дифференцировку на эффекторные клетки и клетки памяти. Последние остаются в организме на всю жизнь.
 память В основу её формирования легли такие свойства нервной системы как") Нейрологическая (нервная) память В основу её формирования легли такие свойства нервной системы как длительное хранение информации о событиях внешнего мира и реакциях организма на эти события, а также использование этой информации для построения текущего поведения. Непрестанно возрастающую совокупность следов от пройденного в прошлом, определяющую поведение в наступающем настоящем А. А. Ухтомский и назвал памятью. Таким образом, память заключается в таких изменениях в НС, которые сохраняются в течение некоторого времени и существенно влияют на характер протекания будущих рефлекторных реакций. Комплекс таких структурно-функциональных изменений получил название процесса образования энграммы.
Нейрологическая (нервная) память В основу её формирования легли такие свойства нервной системы как длительное хранение информации о событиях внешнего мира и реакциях организма на эти события, а также использование этой информации для построения текущего поведения. Непрестанно возрастающую совокупность следов от пройденного в прошлом, определяющую поведение в наступающем настоящем А. А. Ухтомский и назвал памятью. Таким образом, память заключается в таких изменениях в НС, которые сохраняются в течение некоторого времени и существенно влияют на характер протекания будущих рефлекторных реакций. Комплекс таких структурно-функциональных изменений получил название процесса образования энграммы.
 Классификация видов памяти по материалу запоминания
Классификация видов памяти по материалу запоминания
 Специфические виды памяти - психологическая характеристика Модально-специфические виды. Мнестические процессы могут быть связаны с деятельностью разных анализаторов, поэтому существуют специфические виды памяти соответственно органам чувств: зрительная, слуховая, тактильная, обонятельная, двигательная. Образная память. Запечатление и воспроизведение картин окружающего мира связаны с синтезом модальноспецифических впечатлений. В этом случае фиксируются сложные образы, объединяющие зрительные, слуховые и другие модально-специфические сигналы. Такую память называют образной. Образная память гибка, спонтанна и обеспечивает длительное хранение следа.
Специфические виды памяти - психологическая характеристика Модально-специфические виды. Мнестические процессы могут быть связаны с деятельностью разных анализаторов, поэтому существуют специфические виды памяти соответственно органам чувств: зрительная, слуховая, тактильная, обонятельная, двигательная. Образная память. Запечатление и воспроизведение картин окружающего мира связаны с синтезом модальноспецифических впечатлений. В этом случае фиксируются сложные образы, объединяющие зрительные, слуховые и другие модально-специфические сигналы. Такую память называют образной. Образная память гибка, спонтанна и обеспечивает длительное хранение следа.
 Эмоциональная память - связана с запоминанием и воспроизведением эмоциональных переживаний, которые могут возникать как при повторном воздействии раздражителей, обусловивших это состояние, так и в отсутствии последних. Эмоционально окрашенное впечатление фиксируется мгновенно и непроизвольно, обеспечивая пополнение подсознательной сферы человеческой психики. Так же непроизвольно информация воспроизводится из эмоциональной памяти. Этот вид памяти во многом сходен с образной, но иногда эмоциональная память оказывается даже более устойчивой, чем образная. Ее морфологической основой предположительно служат распределенные нервные сети, включающие нейрональные группы из разных отделов коры и ближайшей подкорки.
Эмоциональная память - связана с запоминанием и воспроизведением эмоциональных переживаний, которые могут возникать как при повторном воздействии раздражителей, обусловивших это состояние, так и в отсутствии последних. Эмоционально окрашенное впечатление фиксируется мгновенно и непроизвольно, обеспечивая пополнение подсознательной сферы человеческой психики. Так же непроизвольно информация воспроизводится из эмоциональной памяти. Этот вид памяти во многом сходен с образной, но иногда эмоциональная память оказывается даже более устойчивой, чем образная. Ее морфологической основой предположительно служат распределенные нервные сети, включающие нейрональные группы из разных отделов коры и ближайшей подкорки.
 - это память на словесные сигналы и символы, обозначающие") Словесно-логическая память (или семантическая) - это память на словесные сигналы и символы, обозначающие как внешние объекты, так и внутренние действия и переживания. Ее морфологическую основу можно схематически представить как упорядоченную последовательность линейных звеньев. В результате, выпадение даже одного звена (например, вследствие органического поражения нервной ткани) ведет к разрыву всей цепи, нарушению последовательности хранимых событий и к выпадению из памяти некоего объема информации.
Словесно-логическая память (или семантическая) - это память на словесные сигналы и символы, обозначающие как внешние объекты, так и внутренние действия и переживания. Ее морфологическую основу можно схематически представить как упорядоченную последовательность линейных звеньев. В результате, выпадение даже одного звена (например, вследствие органического поражения нервной ткани) ведет к разрыву всей цепи, нарушению последовательности хранимых событий и к выпадению из памяти некоего объема информации.
 Эпизодическая Семантическая Недекларативная (имплицитная) Неассоциативная: Габитуация Сенситизация Ассоциативная: Условные Рефлексы Классический") Память Декларативная (эксплицитная) Эпизодическая Семантическая Недекларативная (имплицитная) Неассоциативная: Габитуация Сенситизация Ассоциативная: Условные Рефлексы Классический Инструментальный
Память Декларативная (эксплицитная) Эпизодическая Семантическая Недекларативная (имплицитная) Неассоциативная: Габитуация Сенситизация Ассоциативная: Условные Рефлексы Классический Инструментальный
 это? » , декларируется, объясняется суть") Декларативная память - на вопрос «что (кто) это? » , декларируется, объясняется суть объекта; иначе – семантическая, вербальная)» и процедурная ( «завязывание галстука» , «вождение авто» – легче показать, чем декларировать, объяснить). Эксплицитная (осознанный процесс) и имплицитная (напримет, латентное обучение) (аналогия - произвольная и непроизвольная память). Эксплицитная память (explicate — ясный, осознанный, англ. ) связана с быстрым осознанным обучением. Ранее этот вид памяти также называли декларативной. С ее помощью человек различает знакомые и незнакомые события. Многочисленные исследования подтвердили, что структуры, обеспечивающие функционирование этого типа памяти, находятся в височных областях мозга.
Декларативная память - на вопрос «что (кто) это? » , декларируется, объясняется суть объекта; иначе – семантическая, вербальная)» и процедурная ( «завязывание галстука» , «вождение авто» – легче показать, чем декларировать, объяснить). Эксплицитная (осознанный процесс) и имплицитная (напримет, латентное обучение) (аналогия - произвольная и непроизвольная память). Эксплицитная память (explicate — ясный, осознанный, англ. ) связана с быстрым осознанным обучением. Ранее этот вид памяти также называли декларативной. С ее помощью человек различает знакомые и незнакомые события. Многочисленные исследования подтвердили, что структуры, обеспечивающие функционирование этого типа памяти, находятся в височных областях мозга.
 В настоящий момент считается, что эксплицитная память на нейрональном уровне представлена пре-, постсинаптическим совпадением активности нейрона. Этот ассоциативный механизм действует в гиппокампе. Имплицитная память связана с медленным обучением, при котором испытуемый иногда сам не знает, чему и как он обучился. В этом типе обучения участвуют сенсорные и моторные области мозга, непосредственно контролирующие сенсорные входы и двигательную активность, которые сочетаются в том или ином задании. На нейрональном уровне имплицитная память обеспечивается премодулирующим ассоциативным механизмом, при котором условный и безусловный раздражители представлены внутри одной клетки конвергенцией двух сигналов (например, кальция и серотонина) на одном и том же ферменте.
В настоящий момент считается, что эксплицитная память на нейрональном уровне представлена пре-, постсинаптическим совпадением активности нейрона. Этот ассоциативный механизм действует в гиппокампе. Имплицитная память связана с медленным обучением, при котором испытуемый иногда сам не знает, чему и как он обучился. В этом типе обучения участвуют сенсорные и моторные области мозга, непосредственно контролирующие сенсорные входы и двигательную активность, которые сочетаются в том или ином задании. На нейрональном уровне имплицитная память обеспечивается премодулирующим ассоциативным механизмом, при котором условный и безусловный раздражители представлены внутри одной клетки конвергенцией двух сигналов (например, кальция и серотонина) на одном и том же ферменте.
 Автобиографическая память - в последнее время по данным локальных поражений мозга, иногда выделяют также автобиографическую память - как промежуточную между эпизодической и семантической. Э. Тульвинг относит автобиографическую память к эпизодической подсистеме в предложенной им дихотомии семантической - эпизодической подсистем памяти.
Автобиографическая память - в последнее время по данным локальных поражений мозга, иногда выделяют также автобиографическую память - как промежуточную между эпизодической и семантической. Э. Тульвинг относит автобиографическую память к эпизодической подсистеме в предложенной им дихотомии семантической - эпизодической подсистем памяти.
 Временная организация памяти Другим основанием для классификации памяти является продолжительность закрепления и сохранения материала. Принято подразделять память на три вида: • иконическую, или сенсорную, память (ИП); • кратковременную, или оперативную, память (КВП); • долговременную, или декларативную, память (ДВП). Иногда последний вид памяти называется пропозиционной, вторичной или семантической. Считается, что каждый из этих видов памяти обеспечивается различными мозговыми процессами и механизмами, связанными с деятельностью функционально и структурно различных мозговых систем.
Временная организация памяти Другим основанием для классификации памяти является продолжительность закрепления и сохранения материала. Принято подразделять память на три вида: • иконическую, или сенсорную, память (ИП); • кратковременную, или оперативную, память (КВП); • долговременную, или декларативную, память (ДВП). Иногда последний вид памяти называется пропозиционной, вторичной или семантической. Считается, что каждый из этих видов памяти обеспечивается различными мозговыми процессами и механизмами, связанными с деятельностью функционально и структурно различных мозговых систем.
 Трехкомпонентная модель памяти — концепция, в которой структура памяти представлена тремя совместно работающими блоками: блоком сенсорных регистров, где информация хранится не более 1 с. почти в полном объеме в форме модально закодированных физических признаков стимуляции, блоком кратковременного хранилища, где объем хранимой в вербально акустическом коде информации небольшой, а длительность хранения (около 30 с) обусловлена проговариванием, перекодированием, выбором способа запоминания, и блоком долговременного хранилища, объем и время хранения которого неограничены, а информация представлена в форме семантических кодов.
Трехкомпонентная модель памяти — концепция, в которой структура памяти представлена тремя совместно работающими блоками: блоком сенсорных регистров, где информация хранится не более 1 с. почти в полном объеме в форме модально закодированных физических признаков стимуляции, блоком кратковременного хранилища, где объем хранимой в вербально акустическом коде информации небольшой, а длительность хранения (около 30 с) обусловлена проговариванием, перекодированием, выбором способа запоминания, и блоком долговременного хранилища, объем и время хранения которого неограничены, а информация представлена в форме семантических кодов.
 КП забывание СР - сенсорные регистры ДП") Трехкомпонентная система памяти вход внимание СР (селекция) КП забывание СР - сенсорные регистры ДП забывание КП - кратковременная память ДП - долговременная память
Трехкомпонентная система памяти вход внимание СР (селекция) КП забывание СР - сенсорные регистры ДП забывание КП - кратковременная память ДП - долговременная память
 Длительность хранения в сенсорной, или иконической, памяти - 250 -400 мс, однако по некоторым данным этот процесс может продолжаться до 4 -х сек. Объем ИП при наличии соответствующей инструкции от 12 до 20 элементов. Длительность хранения в кратковременной памяти около 12 сек. , при повторении дольше. Объем КВП представлен широко известным числом Миллера 7 ± 2 элемента. Длительность хранения в ДВП неопределенно долгая, объем велик, по некоторым представлениям, неограничен. Временную типологию памяти подтверждают эксперименты с животными, в которых показано, что запоминание ухудшается, если сразу же за научением следует удар электрическим током (т. е. ЭКШ препятствует переносу информации из КП в ДП.
Длительность хранения в сенсорной, или иконической, памяти - 250 -400 мс, однако по некоторым данным этот процесс может продолжаться до 4 -х сек. Объем ИП при наличии соответствующей инструкции от 12 до 20 элементов. Длительность хранения в кратковременной памяти около 12 сек. , при повторении дольше. Объем КВП представлен широко известным числом Миллера 7 ± 2 элемента. Длительность хранения в ДВП неопределенно долгая, объем велик, по некоторым представлениям, неограничен. Временную типологию памяти подтверждают эксперименты с животными, в которых показано, что запоминание ухудшается, если сразу же за научением следует удар электрическим током (т. е. ЭКШ препятствует переносу информации из КП в ДП.
 Аналогично, травма, полученная человеком, не сказывается сразу на воспроизведении событий, но уже через несколько минут человек не может точно вспомнить всех обстоятельств происшествия. О существовании двух различных хранилищ памяти (долговременного и кратковременного) свидетельствуют такие факты. Две группы испытуемых — здоровые и больные амнезией должны были воспроизвести список из 10 слов сразу после заучивания и с задержкой в 30 сек. В момент задержки испытуемые обеих групп должны были решать арифметическую задачу. Значимых различий между двумя группами испытуемых при немедленном воспроизведении обнаружено не было, в то время как при отсроченном воспроизведении у больных амнезией объем запоминания был намного ниже. Этот факт подтверждает, что механизмы КВП и ДВП у человека различны.
Аналогично, травма, полученная человеком, не сказывается сразу на воспроизведении событий, но уже через несколько минут человек не может точно вспомнить всех обстоятельств происшествия. О существовании двух различных хранилищ памяти (долговременного и кратковременного) свидетельствуют такие факты. Две группы испытуемых — здоровые и больные амнезией должны были воспроизвести список из 10 слов сразу после заучивания и с задержкой в 30 сек. В момент задержки испытуемые обеих групп должны были решать арифметическую задачу. Значимых различий между двумя группами испытуемых при немедленном воспроизведении обнаружено не было, в то время как при отсроченном воспроизведении у больных амнезией объем запоминания был намного ниже. Этот факт подтверждает, что механизмы КВП и ДВП у человека различны.
 Во всех вышеперечисленных видах памяти имеют место: Кодирование, в процессе которого выделяется та информация, которая должна храниться; Консолидация – переход информации в долговременную память; Хранение и связывание новой информации с уже хранящейся; Воспроизведение (извлечение) информации из памяти. Забывание информации может быть связано с различными факторами.
Во всех вышеперечисленных видах памяти имеют место: Кодирование, в процессе которого выделяется та информация, которая должна храниться; Консолидация – переход информации в долговременную память; Хранение и связывание новой информации с уже хранящейся; Воспроизведение (извлечение) информации из памяти. Забывание информации может быть связано с различными факторами.
 каждый вид информации") В итоге, из простого анализа типов классификаций памяти следует, что: 1) каждый вид информации может (и часто одновременно) храниться в разных видах П; 2) механизмы П – это, скорее всего, не отдельный локус, но «сеть множества локусов» , распределенная по мозгу «сетевая (узловая) система» . Причем, как следует из «семантических моделей П» (измерение времени принятия решения о сходстве далеких и близких по смыслу понятий), это именно сетевая неэкономно организованная структура, допускающая – в отличие от структуры графа – «странные» неупорядоченные прямые связи «всех со всеми» . В мат. «граф» – это некоторая экономно построенная структура ( «дерево поиска» ), при которой существует единственный путь (поиска) связи между любыми двумя точками ( «узлами графа» – понятиями).
В итоге, из простого анализа типов классификаций памяти следует, что: 1) каждый вид информации может (и часто одновременно) храниться в разных видах П; 2) механизмы П – это, скорее всего, не отдельный локус, но «сеть множества локусов» , распределенная по мозгу «сетевая (узловая) система» . Причем, как следует из «семантических моделей П» (измерение времени принятия решения о сходстве далеких и близких по смыслу понятий), это именно сетевая неэкономно организованная структура, допускающая – в отличие от структуры графа – «странные» неупорядоченные прямые связи «всех со всеми» . В мат. «граф» – это некоторая экономно построенная структура ( «дерево поиска» ), при которой существует единственный путь (поиска) связи между любыми двумя точками ( «узлами графа» – понятиями).
 Локализация механизмов памяти в мозге: данные нормы и патологии. Данные о существовании образной П, полученные при раздражении мозга. В простейших случаях раздражения периферических сенсорных систем (глаз; уши) возникают т. н. “фосфены” предметно не оформленные ощущения – света, звука, вкуса и т. п. ). Фосфены могут возникать спонтанно при разных видах сенсорной депривации и интерпретироваться как “озарения”, “видения” (условия занятия медитацией близки к условиям сенсорной депривации – темнота, концентрация на собственном “Я”). По этой причине фосфены получили название “кино узников”.
Локализация механизмов памяти в мозге: данные нормы и патологии. Данные о существовании образной П, полученные при раздражении мозга. В простейших случаях раздражения периферических сенсорных систем (глаз; уши) возникают т. н. “фосфены” предметно не оформленные ощущения – света, звука, вкуса и т. п. ). Фосфены могут возникать спонтанно при разных видах сенсорной депривации и интерпретироваться как “озарения”, “видения” (условия занятия медитацией близки к условиям сенсорной депривации – темнота, концентрация на собственном “Я”). По этой причине фосфены получили название “кино узников”.
 возникают при раздражении гол. мозга. Например, при") Более сложные картины (образы восприятия, сцены) возникают при раздражении гол. мозга. Например, при раздражении первичной зрительной коры возникают образы звезд, колес, шаров, спиралей. При раздражении первичной слуховой коры – звон, щелчки, гуденье, щебетанье. Так, на 1000 испыт. получены данные о 15 типичных зрительных фигурах (простые геометрические фигуры), возникающих при наружном раздражении (1 м. В, 1 м. А) виска. Это соответствует данным нейрофизиологии о наличии в средней и верхней височных извилинах “детекторов форм”. Далее, интересно, что максимальная выраженность фигур была при частоте стимуляции в пределах 5 -40 Гц, что соответствует диапазону биоритмов мозга.
Более сложные картины (образы восприятия, сцены) возникают при раздражении гол. мозга. Например, при раздражении первичной зрительной коры возникают образы звезд, колес, шаров, спиралей. При раздражении первичной слуховой коры – звон, щелчки, гуденье, щебетанье. Так, на 1000 испыт. получены данные о 15 типичных зрительных фигурах (простые геометрические фигуры), возникающих при наружном раздражении (1 м. В, 1 м. А) виска. Это соответствует данным нейрофизиологии о наличии в средней и верхней височных извилинах “детекторов форм”. Далее, интересно, что максимальная выраженность фигур была при частоте стимуляции в пределах 5 -40 Гц, что соответствует диапазону биоритмов мозга.
 связь между частотой стимуляции") Более того, для каждого испытуемого имелась устойчивая (несколько месяцев) связь между частотой стимуляции и содержанием фигур, что свидетельствует о том, что обработка информации производится устойчивыми специализированными нейронными сетями со своими резонансными частотами. По данным нейрохирургии этой области мозга – у больного возникают образы и эпизоды из прошлой жизни (звучание оркестра; зрит. сцены). Этот эффект, обнаружен в работах знаменитого нейрохирурга В. Пенфильда (50 -60 г. г. ХХв. ) и назван “flash-back” – «вспышка пережитого» . Важно отметить, что содержание видений человека не имело в прошлом для него никакого значения, это говорит о том, что мы запоминаем намного больше, чем вспоминаем.
Более того, для каждого испытуемого имелась устойчивая (несколько месяцев) связь между частотой стимуляции и содержанием фигур, что свидетельствует о том, что обработка информации производится устойчивыми специализированными нейронными сетями со своими резонансными частотами. По данным нейрохирургии этой области мозга – у больного возникают образы и эпизоды из прошлой жизни (звучание оркестра; зрит. сцены). Этот эффект, обнаружен в работах знаменитого нейрохирурга В. Пенфильда (50 -60 г. г. ХХв. ) и назван “flash-back” – «вспышка пережитого» . Важно отметить, что содержание видений человека не имело в прошлом для него никакого значения, это говорит о том, что мы запоминаем намного больше, чем вспоминаем.
 Близкие к эффекту “flash-back” – феномен “deja vu” (эффект ложной П типа «ранее пережитого» ) и обратный эффект «странности (незнакомости) вроде бы привычной ситуации» . Возможно, эти эффекты – результаты ошибок сопоставления текущих данных рабочей П со следами ДП. Механизм эффекта «дежа вю» связан с появлением в височной коре высокоамплитудных синхронизированных колебаний (как и в опытах с искусств. стимуляцией виска). Этим объясняется повышенная частота эффектов ДЖВ у больных с височной эпилепсией (предшественник припадка).
Близкие к эффекту “flash-back” – феномен “deja vu” (эффект ложной П типа «ранее пережитого» ) и обратный эффект «странности (незнакомости) вроде бы привычной ситуации» . Возможно, эти эффекты – результаты ошибок сопоставления текущих данных рабочей П со следами ДП. Механизм эффекта «дежа вю» связан с появлением в височной коре высокоамплитудных синхронизированных колебаний (как и в опытах с искусств. стимуляцией виска). Этим объясняется повышенная частота эффектов ДЖВ у больных с височной эпилепсией (предшественник припадка).
 Возможно, этот же эффект лежит в основе представлений о переселении душ, или реинкарнации. Действительно, ложное ощущение того, что ты уже был здесь, рождает предположение, что ты возможно был здесь в другой жизни. Аномалии памяти (парамнезии) -это не патология, это редко, но бывает практически у всех. К ним относятся: феномен «уже пережитого» (“flash-back” , “deja vu” ) Конфабуляции - «вспоминаются» события, которых на самом деле не было. Это не ложь - человек верит, что это с ним было. Криптомнезии - зашифрованное воспоминание, пример – ложный плагиат, т. е. то, что было в действительности выдается за нечто новое.
Возможно, этот же эффект лежит в основе представлений о переселении душ, или реинкарнации. Действительно, ложное ощущение того, что ты уже был здесь, рождает предположение, что ты возможно был здесь в другой жизни. Аномалии памяти (парамнезии) -это не патология, это редко, но бывает практически у всех. К ним относятся: феномен «уже пережитого» (“flash-back” , “deja vu” ) Конфабуляции - «вспоминаются» события, которых на самом деле не было. Это не ложь - человек верит, что это с ним было. Криптомнезии - зашифрованное воспоминание, пример – ложный плагиат, т. е. то, что было в действительности выдается за нечто новое.
 Данные “за” и “против” разделения П на Д и Кр. П. В пользу существования этапа Кр. П свидетельствуют данные о динамике “последовательных зрительных послеобразов” (“положит. ” послеобраз Геринга в интервале 30 -80 мс; ). Они являются, свидетельством механизма продления “образа восприятия” с целью формирования следа П. Роль гиппокампа (в переводе информации в долговр. п. – болезнь Альцгеймера; связь с ОР, нейрогенез в гиппокампе как функция упражнения). “Детская амнезия” – не помним ничего до 3 -4 лет (момента созревания гиппокампа). Префронтальная кора и рабочая память. Опыты с обезьянами по отсроченным реакциям. Дофамин: инъекции старым об.
Данные “за” и “против” разделения П на Д и Кр. П. В пользу существования этапа Кр. П свидетельствуют данные о динамике “последовательных зрительных послеобразов” (“положит. ” послеобраз Геринга в интервале 30 -80 мс; ). Они являются, свидетельством механизма продления “образа восприятия” с целью формирования следа П. Роль гиппокампа (в переводе информации в долговр. п. – болезнь Альцгеймера; связь с ОР, нейрогенез в гиппокампе как функция упражнения). “Детская амнезия” – не помним ничего до 3 -4 лет (момента созревания гиппокампа). Префронтальная кора и рабочая память. Опыты с обезьянами по отсроченным реакциям. Дофамин: инъекции старым об.
 Как свидетельствуют современные данные, хранение следов в памяти – это процесс, развернутый во времени. По отношению к ДП – степень нарушения (амнезии) зависит не только от локуса поражения, но и от времени нанесения повреждающего воздействия на этот локус. Роль активности субъекта в запом. и воспроизвед. : перевод из ОП в ДП – не просто (пассивная) функция времени нахождения следа памяти в ОП (от уровня обработки инф. , связи с личным опытом – мнемоника). При воспроизв. субъект часто кое-что домысливает, интерпретирует. В этом смысле, по аналогии с выражением “мысль изреченная есть ложь“ можно сказать, что “извлеченное из П не тождественно памяти” (“след П хранится, а П создается”).
Как свидетельствуют современные данные, хранение следов в памяти – это процесс, развернутый во времени. По отношению к ДП – степень нарушения (амнезии) зависит не только от локуса поражения, но и от времени нанесения повреждающего воздействия на этот локус. Роль активности субъекта в запом. и воспроизвед. : перевод из ОП в ДП – не просто (пассивная) функция времени нахождения следа памяти в ОП (от уровня обработки инф. , связи с личным опытом – мнемоника). При воспроизв. субъект часто кое-что домысливает, интерпретирует. В этом смысле, по аналогии с выражением “мысль изреченная есть ложь“ можно сказать, что “извлеченное из П не тождественно памяти” (“след П хранится, а П создается”).
 Нарушения памяти при локальных поражениях мозга также свидетельствует о том, что в механизмах П задействованы многие области мозга и что, механизм П – это “распределенная система мозга”. мнезия любая памяти масштабу - потеря своему по и степени значительно превосходящая обычное забывание Органические: §ретроградная §антеро-ретроградная Функциональные: §защитная §эпилептическая §постгипнотическая §на сновидения §посталкогольная §инфантильная
Нарушения памяти при локальных поражениях мозга также свидетельствует о том, что в механизмах П задействованы многие области мозга и что, механизм П – это “распределенная система мозга”. мнезия любая памяти масштабу - потеря своему по и степени значительно превосходящая обычное забывание Органические: §ретроградная §антеро-ретроградная Функциональные: §защитная §эпилептическая §постгипнотическая §на сновидения §посталкогольная §инфантильная
 – человек с уникальной памятью. Ким прочитывает страницу за 8") Ким Пик (Kim Peek) – человек с уникальной памятью. Ким прочитывает страницу за 8 - 10 секунд, сразу запоминая ее наизусть. В его памяти хранится 9 тыс. книг, охватывающих огромные области знания от Шекспира до композиторов и карт всех крупных городов США. В мозге Кима обнаруживается множество аномалий, в том числе отсутствие мозолистого тела. У него также деформирован мозжечок, из-за чего многие простые движения в быту даются ему с трудом. В отличие от подавляющего большинства людей с феноменальной памятью, страдающих аутизмом и умственной отсталостью, Ким смог отчасти избавиться от своего аутизма, а недавно неплохо освоил игру на фортепиано. Послужил прототипом главного героя фильма «Человека дождя» , хотя автобиографические детали в фильме не соответствуют реальности. Сходный случай описан в книге Лурия А. P. Маленькая книжка о большой памяти, 1968.
Ким Пик (Kim Peek) – человек с уникальной памятью. Ким прочитывает страницу за 8 - 10 секунд, сразу запоминая ее наизусть. В его памяти хранится 9 тыс. книг, охватывающих огромные области знания от Шекспира до композиторов и карт всех крупных городов США. В мозге Кима обнаруживается множество аномалий, в том числе отсутствие мозолистого тела. У него также деформирован мозжечок, из-за чего многие простые движения в быту даются ему с трудом. В отличие от подавляющего большинства людей с феноменальной памятью, страдающих аутизмом и умственной отсталостью, Ким смог отчасти избавиться от своего аутизма, а недавно неплохо освоил игру на фортепиано. Послужил прототипом главного героя фильма «Человека дождя» , хотя автобиографические детали в фильме не соответствуют реальности. Сходный случай описан в книге Лурия А. P. Маленькая книжка о большой памяти, 1968.
 – (от лат. “gradior” – идти) -") Антероградная амнезия (от греч. “mneme” – память) – (от лат. “gradior” – идти) - после травмы нарушено формирование ДП на будущие события (процедурная П сохранна). Пациент не может вспомнить новый (только что предъявленный) материал. Кр. П сохранна в течение десятков секунд. Словно, нарушен перевод информации из Кр. П в ДП. Примеры: Симптом в составе синдрома Корсакова (конец XIXв. ; хронический алкоголизм: страдают мамиллярные тела и таламус), при болезни Альцгеймера (прозрачная перегородка, гиппокамп). АА свидетельствует о раздельном существовании двух видов П – Кр. П (рабочей) и ДП (на прошлые события она сохранна).
Антероградная амнезия (от греч. “mneme” – память) – (от лат. “gradior” – идти) - после травмы нарушено формирование ДП на будущие события (процедурная П сохранна). Пациент не может вспомнить новый (только что предъявленный) материал. Кр. П сохранна в течение десятков секунд. Словно, нарушен перевод информации из Кр. П в ДП. Примеры: Симптом в составе синдрома Корсакова (конец XIXв. ; хронический алкоголизм: страдают мамиллярные тела и таламус), при болезни Альцгеймера (прозрачная перегородка, гиппокамп). АА свидетельствует о раздельном существовании двух видов П – Кр. П (рабочей) и ДП (на прошлые события она сохранна).
 Ретроградная амнезия - пациент не способен вспомнить материал, предшествующий травме. Например, после сеансов электрошоковой терапии больных депрессий и маний. Длительность заболевания – от нескольких дней до нескольких лет. Забываются разные периоды – от десятков секунд до самых “далеких”. 1) нарушается не только содержание следов П, но и их хронология, 2) возникает не сразу, а через несколько секунд/минут после травмы, 3) частично восстанавливается. Вывод, сформированные в ДП следы не стираются и в основе РА лежит временное (обратимое) нарушение механизма воспроизведения следов из П. Механизм доступа к следам ДП связывают с востановлением функций неспецифической системы активации мозга.
Ретроградная амнезия - пациент не способен вспомнить материал, предшествующий травме. Например, после сеансов электрошоковой терапии больных депрессий и маний. Длительность заболевания – от нескольких дней до нескольких лет. Забываются разные периоды – от десятков секунд до самых “далеких”. 1) нарушается не только содержание следов П, но и их хронология, 2) возникает не сразу, а через несколько секунд/минут после травмы, 3) частично восстанавливается. Вывод, сформированные в ДП следы не стираются и в основе РА лежит временное (обратимое) нарушение механизма воспроизведения следов из П. Механизм доступа к следам ДП связывают с востановлением функций неспецифической системы активации мозга.
 связь между уровнем активации мозга и эффективностью запоминания и") Существует (показано в норме) связь между уровнем активации мозга и эффективностью запоминания и вспоминания, описанная уже известным вам законом Йеркса-Додсона. Это общебиологический и общепсихологический закон: оптимум ФС необходим не только для П, но и для реализации стрессоустойчивости, концентрации внимания, принятия и реализации адекватных решений, управления собственным поведением и поведением других людей.
Существует (показано в норме) связь между уровнем активации мозга и эффективностью запоминания и вспоминания, описанная уже известным вам законом Йеркса-Додсона. Это общебиологический и общепсихологический закон: оптимум ФС необходим не только для П, но и для реализации стрессоустойчивости, концентрации внимания, принятия и реализации адекватных решений, управления собственным поведением и поведением других людей.
 Данные наблюдений в клинике локальных поражений головного мозга – в пользу выделения “эпизодической П” и “П на общую информацию”. Данные клиники свидетельствуют также о том, что в П вовлечены очень многие структуры мозга: поражения префронтальной коры - приводят к конфабуляциям (смешениям), нарушениям оперативной П; миндалины – к нарушениям П на эмоциональные события (особенно, связанные со страхом), обучения с одной попытки; затылочные и височные области коры – нарушению П на зрительные события (например, прозопагнозия- неузнавание хорошо знакомых лиц ).
Данные наблюдений в клинике локальных поражений головного мозга – в пользу выделения “эпизодической П” и “П на общую информацию”. Данные клиники свидетельствуют также о том, что в П вовлечены очень многие структуры мозга: поражения префронтальной коры - приводят к конфабуляциям (смешениям), нарушениям оперативной П; миндалины – к нарушениям П на эмоциональные события (особенно, связанные со страхом), обучения с одной попытки; затылочные и височные области коры – нарушению П на зрительные события (например, прозопагнозия- неузнавание хорошо знакомых лиц ).
 При поражении гиппокампа и других областей медиальной части височной доли (а также медиодорсального таламуса) нарушается декларативная память. Возникает как антероградная амнезия (нарушение запоминанию нового материала), так и некоторая ретроградная амнезия (вплоть до нескольких лет, предшествующих операции или травме). Память на более отдаленные события сохраняется в норме. Интеллект в норме или даже выше нормы. Процедурная память у этих больных не нарушена и не отличается от таковой у здоровых людей.
При поражении гиппокампа и других областей медиальной части височной доли (а также медиодорсального таламуса) нарушается декларативная память. Возникает как антероградная амнезия (нарушение запоминанию нового материала), так и некоторая ретроградная амнезия (вплоть до нескольких лет, предшествующих операции или травме). Память на более отдаленные события сохраняется в норме. Интеллект в норме или даже выше нормы. Процедурная память у этих больных не нарушена и не отличается от таковой у здоровых людей.
 Brain areas that, when damaged, tend to give rise to declarative memory disorders. By inference, declarative memory is based on the physiological activity of these structures. (A) Studies of amnesic patients have shown that the formation of declarative memories depends on the integrity of the hippocampus and its subcortical connections to the mammillary bodies and dorsal thalamus. (B) Diagram showing the location of the hippocampus in a cutaway view in the horizontal plane. (C) The hippocampus as it would appear in a histological section in the coronal plane, at approximately the level indicated by the line in (B).
Brain areas that, when damaged, tend to give rise to declarative memory disorders. By inference, declarative memory is based on the physiological activity of these structures. (A) Studies of amnesic patients have shown that the formation of declarative memories depends on the integrity of the hippocampus and its subcortical connections to the mammillary bodies and dorsal thalamus. (B) Diagram showing the location of the hippocampus in a cutaway view in the horizontal plane. (C) The hippocampus as it would appear in a histological section in the coronal plane, at approximately the level indicated by the line in (B).
") Изучение больных с амнезией, а также открытие длительной потенциации (удобной экспериментальной модели памяти) в гиппокампе привело к ошибочной точке зрения, что гиппокамп и является местом хранения памяти. На самом деле, видимо, гиппокамп необходим для кодирования и консолидации декларативной памяти, а также, возможно, ее воспроизведения, однако сама по себе долговременная память хранится преимущественно не в гиппокампе.
Изучение больных с амнезией, а также открытие длительной потенциации (удобной экспериментальной модели памяти) в гиппокампе привело к ошибочной точке зрения, что гиппокамп и является местом хранения памяти. На самом деле, видимо, гиппокамп необходим для кодирования и консолидации декларативной памяти, а также, возможно, ее воспроизведения, однако сама по себе долговременная память хранится преимущественно не в гиппокампе.
 «Считается, что долговременная память связана с ассоциативной корой. В адресации памятных следов в определенные участки коры - важную роль играют медиальные отделы височной области полушарий, включающие энторинальную кору и гиппокамп … Вышеназванные образования имеют обширные связи как между собой, так и с проекционными (теми, куда приходят сигналы от органов чувств) и ассоциативными отделами коры. При запоминании они направляют сигнал в ассоциативную кору для длительного удержания в памяти, а при необходимости вспомнить - указывают адрес, где хранится связанная с поступившим сигналом информация. Приведем простой пример. Долговременная память соответствует книгохранилищу в библиотеке, а гиппокампальный комплекс можно сравнить с каталогом, который показывает, где хранится нужная книга. » А. М. Иваницкий
«Считается, что долговременная память связана с ассоциативной корой. В адресации памятных следов в определенные участки коры - важную роль играют медиальные отделы височной области полушарий, включающие энторинальную кору и гиппокамп … Вышеназванные образования имеют обширные связи как между собой, так и с проекционными (теми, куда приходят сигналы от органов чувств) и ассоциативными отделами коры. При запоминании они направляют сигнал в ассоциативную кору для длительного удержания в памяти, а при необходимости вспомнить - указывают адрес, где хранится связанная с поступившим сигналом информация. Приведем простой пример. Долговременная память соответствует книгохранилищу в библиотеке, а гиппокампальный комплекс можно сравнить с каталогом, который показывает, где хранится нужная книга. » А. М. Иваницкий
 Connections between the hippocampus and possible declarative memory storage sites. The rhesus monkey brain is shown because these connections are much better documented in non-human primates than in humans. Projections from numerous cortical areas converge on the hippocampus and the related structures known to be involved in human memory; most of these sites also send projections to the same cortical areas. Medial and lateral views are shown, the latter rotated 180° for clarity.
Connections between the hippocampus and possible declarative memory storage sites. The rhesus monkey brain is shown because these connections are much better documented in non-human primates than in humans. Projections from numerous cortical areas converge on the hippocampus and the related structures known to be involved in human memory; most of these sites also send projections to the same cortical areas. Medial and lateral views are shown, the latter rotated 180° for clarity.
 Декларативная память хранится во всех специализированных ассоциативных областях коры больших полушарий. Процедурная память также широко распространена по всей коре больших полушарий, и связана преимущественно с сенсорными и моторными областями коры, а также с мозжечком. Согласно данным томографических исследований на людях, при воспоминании изображений или звуков активируются те же самые области коры, что и при их восприятии соответствующих стимулов. (см. след. рис. )
Декларативная память хранится во всех специализированных ассоциативных областях коры больших полушарий. Процедурная память также широко распространена по всей коре больших полушарий, и связана преимущественно с сенсорными и моторными областями коры, а также с мозжечком. Согласно данным томографических исследований на людях, при воспоминании изображений или звуков активируются те же самые области коры, что и при их восприятии соответствующих стимулов. (см. след. рис. )
 Subjects were") Reactivation of visual cortex during vivid remembering of visual view images. (A) Subjects were instructed to view either images of objects (houses, faces, and chairs) (left) or imagine the objects in the absence of the stimulus (right). (B) (Left) Bilateral regions of ventral temporal cortex are specifically activated during perception of houses (yellow), faces (red), and chairs (blue). (Right) When subjects recall these objects, the same regions preferentially activated during the perception of each object class are reactivated.
Reactivation of visual cortex during vivid remembering of visual view images. (A) Subjects were instructed to view either images of objects (houses, faces, and chairs) (left) or imagine the objects in the absence of the stimulus (right). (B) (Left) Bilateral regions of ventral temporal cortex are specifically activated during perception of houses (yellow), faces (red), and chairs (blue). (Right) When subjects recall these objects, the same regions preferentially activated during the perception of each object class are reactivated.
 Механизмы кратковременной и долговременной памяти
Механизмы кратковременной и долговременной памяти
: «Если аксон клетки А расположен настолько близко") A B Правило Хебба (1949 г. ): «Если аксон клетки А расположен настолько близко к клетке В, что может возбуждать ее, и если он многократно и непрерывно принимает участие в ее активации, то в одной или обеих клетках возникают какой-то процесс роста или метаболические изменения, и в результате эффективность клетки А как одного из активаторов клетки В возрастает» . Примечание: подразумевается, что клетка B может быть активирована каким-то другим способом, помимо синапса с клетки A – т. е. либо имеются и другие входы на клетку B, причем сильные, либо клетка B спонтанно активна.
A B Правило Хебба (1949 г. ): «Если аксон клетки А расположен настолько близко к клетке В, что может возбуждать ее, и если он многократно и непрерывно принимает участие в ее активации, то в одной или обеих клетках возникают какой-то процесс роста или метаболические изменения, и в результате эффективность клетки А как одного из активаторов клетки В возрастает» . Примечание: подразумевается, что клетка B может быть активирована каким-то другим способом, помимо синапса с клетки A – т. е. либо имеются и другие входы на клетку B, причем сильные, либо клетка B спонтанно активна.
 сформулировал теоретически возможный механизм хранения") Нейронные механизмы фиксации следов памяти Д. Хебб (Hebb, 1949) сформулировал теоретически возможный механизм хранения информации. С его точки зрения, основой обучения может быть совпадение во времени активности пресинаптической и постсинаптической клеток. В дальнейшем такого типа связь между клетками стала называться синапсом Хебба.
Нейронные механизмы фиксации следов памяти Д. Хебб (Hebb, 1949) сформулировал теоретически возможный механизм хранения информации. С его точки зрения, основой обучения может быть совпадение во времени активности пресинаптической и постсинаптической клеток. В дальнейшем такого типа связь между клетками стала называться синапсом Хебба.
 Больше постсинаптических рецепторов, сильнее реакция на нейромедиатор Дендритный шипик Выделение большего количества нейромедиатора Спраутинг: образование новой дополнительной терминали Утолщение и укорочение шейки шипика, что снижает его электрическое сопротивление Четыре возможных способа повышения эффективности синапса
Больше постсинаптических рецепторов, сильнее реакция на нейромедиатор Дендритный шипик Выделение большего количества нейромедиатора Спраутинг: образование новой дополнительной терминали Утолщение и укорочение шейки шипика, что снижает его электрическое сопротивление Четыре возможных способа повышения эффективности синапса
 Сейчас считается, что эксплицитная память на нейрональном уровне представлена препостсинаптическим совпадением активности нейрона. В 1986 г. Б. Вигстрем и X. Густафсон показали, что препостсинаптический ассоциативный механизм действует в гиппокампе. В этом случае при прохождении электрического импульса через терминаль пресинаптического аксона происходит выход медиатора - глутамата в синаптическую щель.
Сейчас считается, что эксплицитная память на нейрональном уровне представлена препостсинаптическим совпадением активности нейрона. В 1986 г. Б. Вигстрем и X. Густафсон показали, что препостсинаптический ассоциативный механизм действует в гиппокампе. В этом случае при прохождении электрического импульса через терминаль пресинаптического аксона происходит выход медиатора - глутамата в синаптическую щель.
 Долговременная потенциация сопровождается деполяризацией постсинаптической мембраны в результате активности HENMDA-рецепторных каналов. Деполяризация снимает блокаду NMDA-каналов магнием, что позволяет кальцию поступать в клетку через эти каналы. Кальций активирует зависимые от него киназы, и это приводит к индукции долговременной потенциации.
Долговременная потенциация сопровождается деполяризацией постсинаптической мембраны в результате активности HENMDA-рецепторных каналов. Деполяризация снимает блокаду NMDA-каналов магнием, что позволяет кальцию поступать в клетку через эти каналы. Кальций активирует зависимые от него киназы, и это приводит к индукции долговременной потенциации.
рецепторы на постсинаптической мембране, что приводит к запуску последовательности") Глутамат активирует NMDA ( N-метил-D-аспартат)рецепторы на постсинаптической мембране, что приводит к запуску последовательности реакций, результатом которых является выработка оксида азота — недавно открытого медиатора нового типа. Молекулы оксида азота невелики и легко проходят сквозь мембрану постсинаптической клетки. Большее количество его инактивируется ферментами, а часть оксида азота, которая возвращается к пресинаптической терминали, возбуждает рецепторы кальциевых каналов в ней, способствуя дополнительному выбросу глутамата из везикул.
Глутамат активирует NMDA ( N-метил-D-аспартат)рецепторы на постсинаптической мембране, что приводит к запуску последовательности реакций, результатом которых является выработка оксида азота — недавно открытого медиатора нового типа. Молекулы оксида азота невелики и легко проходят сквозь мембрану постсинаптической клетки. Большее количество его инактивируется ферментами, а часть оксида азота, которая возвращается к пресинаптической терминали, возбуждает рецепторы кальциевых каналов в ней, способствуя дополнительному выбросу глутамата из везикул.
 Измение формы и размеров пре- и постсинаптических структур – морфофизиологический субстрат ДП. Основной механизм - посттетаническая потенциация. Этот механизм особенно хорошо выражен в таких структурах памяти как гиппокамп и миндалина. Пресинаптический механизм (рост числа синаптических бутонов – нейрогенез). Модуляция кальциевых и калиевых каналов с помощью серотонин-эргических МН. Постсинаптический механизм (рост числа ионных каналов).
Измение формы и размеров пре- и постсинаптических структур – морфофизиологический субстрат ДП. Основной механизм - посттетаническая потенциация. Этот механизм особенно хорошо выражен в таких структурах памяти как гиппокамп и миндалина. Пресинаптический механизм (рост числа синаптических бутонов – нейрогенез). Модуляция кальциевых и калиевых каналов с помощью серотонин-эргических МН. Постсинаптический механизм (рост числа ионных каналов).
 Особое значение в процессах памяти и обучения придают NMDA-рецепторам, с которыми связывается глютаминовая к-та (N-метил-D-аспартат). В мозге млекопитающих эти рецепторы локализованы, главным образом, в коре, базальных ганглиях и сенсорноассоциативных системах. Наивысшая плотность – в гиппокампе. При связывании глютамата с NMDA-рецепторами в клетку входят ионы натрия и кальция. Последний вступает в контакт с кальций-кальмодулин-зависимой киназой II, активация которой и запускает непосредственно цикл внутриклеточных биохимических реакций, повышающих чувствительность нейрона к поступающим пресинаптическим сигналам. Так, м. б. объяснен, феномен посттетанической потенциации (LTP: long-term potentiation).
Особое значение в процессах памяти и обучения придают NMDA-рецепторам, с которыми связывается глютаминовая к-та (N-метил-D-аспартат). В мозге млекопитающих эти рецепторы локализованы, главным образом, в коре, базальных ганглиях и сенсорноассоциативных системах. Наивысшая плотность – в гиппокампе. При связывании глютамата с NMDA-рецепторами в клетку входят ионы натрия и кальция. Последний вступает в контакт с кальций-кальмодулин-зависимой киназой II, активация которой и запускает непосредственно цикл внутриклеточных биохимических реакций, повышающих чувствительность нейрона к поступающим пресинаптическим сигналам. Так, м. б. объяснен, феномен посттетанической потенциации (LTP: long-term potentiation).
 Есть данные и за участие в этом процессе изменения эффективности синапса также пресинаптических структур (увеличение выброса медиатора). В настоящее время имеются две гипотезы относительно изменения синаптических контактов под влиянием обучения: функциональная - обучение не меняет количество синапсов, но увеличивает отношение активных синапсов к пассивным; структурная - обучение стимулирует образование новых контактов между нейронами, которые создаются заново.
Есть данные и за участие в этом процессе изменения эффективности синапса также пресинаптических структур (увеличение выброса медиатора). В настоящее время имеются две гипотезы относительно изменения синаптических контактов под влиянием обучения: функциональная - обучение не меняет количество синапсов, но увеличивает отношение активных синапсов к пассивным; структурная - обучение стимулирует образование новых контактов между нейронами, которые создаются заново.
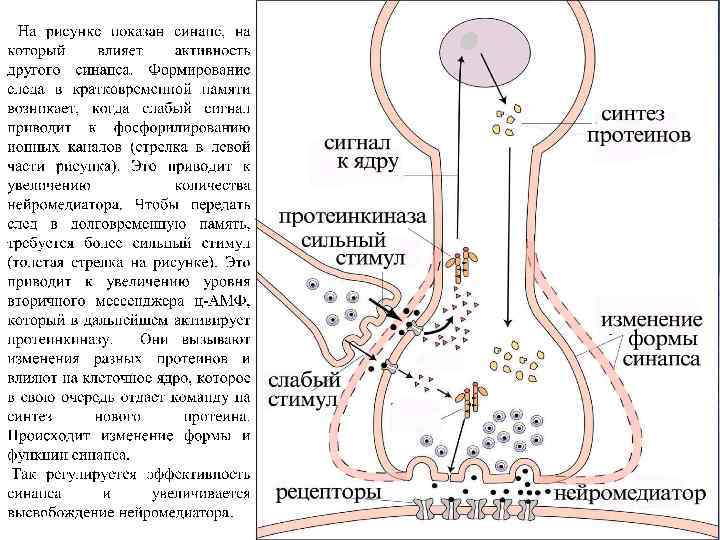
 Каскад молекулярных биохимических реакций, лежащих в основе формирования следа памяти. Опыты на цыплятах: поиск механизмов формирования условной реакции избегания цветных бусинок с определенным запахом с использованием меченой по углероду 2 -дезоксирибозы (выявление локусов мозга, связанных с обучением) и меченой по фосфору АТФ (источника энергии процессах фосфорилирования (мембранных) белков в нейронах из “локусов памяти”, предварительно определенныых с помощью 2 дезоксиглюкозы) из “локусов памяти”. Выявили последовательность (каскад) биохимических реакций, сопровождающих формирование следов П на условный раздражитель(“бусинка определенного цвета и запаха”) в соответствующих нейронах.
Каскад молекулярных биохимических реакций, лежащих в основе формирования следа памяти. Опыты на цыплятах: поиск механизмов формирования условной реакции избегания цветных бусинок с определенным запахом с использованием меченой по углероду 2 -дезоксирибозы (выявление локусов мозга, связанных с обучением) и меченой по фосфору АТФ (источника энергии процессах фосфорилирования (мембранных) белков в нейронах из “локусов памяти”, предварительно определенныых с помощью 2 дезоксиглюкозы) из “локусов памяти”. Выявили последовательность (каскад) биохимических реакций, сопровождающих формирование следов П на условный раздражитель(“бусинка определенного цвета и запаха”) в соответствующих нейронах.
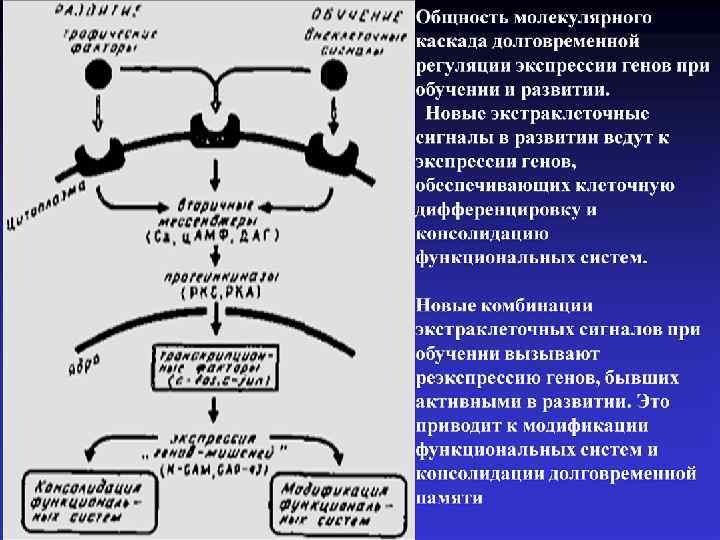
 через") Схема двухфазной регуляции экспрессии генов в клетке. Внеклеточные стимулы (гормоны, факторы роста, нейромедиаторы) через посредство вторичных мессенджеров вызывают активацию транскрипции "непосредственных ранних генов", в частности генов из семейств fos и jun. Синтезирующиеся при этом белки Fos и Jun быстро транспортируются в ядро, где образовывают гетеро- и гомодимерные комплексы. Эти комплексы обладают специфической ДНК-связывающей активностью и способны изменять транскрипцию многих других генов - "поздних" генов
Схема двухфазной регуляции экспрессии генов в клетке. Внеклеточные стимулы (гормоны, факторы роста, нейромедиаторы) через посредство вторичных мессенджеров вызывают активацию транскрипции "непосредственных ранних генов", в частности генов из семейств fos и jun. Синтезирующиеся при этом белки Fos и Jun быстро транспортируются в ядро, где образовывают гетеро- и гомодимерные комплексы. Эти комплексы обладают специфической ДНК-связывающей активностью и способны изменять транскрипцию многих других генов - "поздних" генов
 Сигнал – выделение глутамата в синапсах – активация вторичных мессенджеров (ц. АМФ и Са 2+) – активация определенных протеинкиназ – фосфорилирование определенных (сигнальных, регуляторных) белков – активация определенных участков ДНК генома ядра клетки – синтез рецепторных белков и гликопротеинов (белок+ остатки сахаров, напр. , глюкозы) для увеличения площади пре- и постсинаптической (“шипики” дендритов) мембраны. Через 12 -24 ч. после обучения в соответствующих “локусах П” гиперстриатума цыплят число шипиков увеличилось на 60%, увеличилось также число синапсов и постсинаптических утолщения (результат формирования ДП, обучения).
Сигнал – выделение глутамата в синапсах – активация вторичных мессенджеров (ц. АМФ и Са 2+) – активация определенных протеинкиназ – фосфорилирование определенных (сигнальных, регуляторных) белков – активация определенных участков ДНК генома ядра клетки – синтез рецепторных белков и гликопротеинов (белок+ остатки сахаров, напр. , глюкозы) для увеличения площади пре- и постсинаптической (“шипики” дендритов) мембраны. Через 12 -24 ч. после обучения в соответствующих “локусах П” гиперстриатума цыплят число шипиков увеличилось на 60%, увеличилось также число синапсов и постсинаптических утолщения (результат формирования ДП, обучения).
 Сенсорный нейрон
Сенсорный нейрон
 1. Сильная стимуляция деполяризует клеточную мембрану. 2. Деполяризация заставляет клетку разрядиться потенциалом действия. 3. Потенциал-чувствительные кальциевые каналы открываются. 4. Ионы кальция активируют ферменты, которые активируют CREB. 5. CREB активирует гены, ответственные за белки, усиливающие синаптическую связь. 6. Белки распространяются по всей клетке, оказывая влияние только на те синапсы, сила которых временно увеличена.
1. Сильная стимуляция деполяризует клеточную мембрану. 2. Деполяризация заставляет клетку разрядиться потенциалом действия. 3. Потенциал-чувствительные кальциевые каналы открываются. 4. Ионы кальция активируют ферменты, которые активируют CREB. 5. CREB активирует гены, ответственные за белки, усиливающие синаптическую связь. 6. Белки распространяются по всей клетке, оказывая влияние только на те синапсы, сила которых временно увеличена.