Лекция 3_ Ядро ооцита.ppt
- Количество слайдов: 22



Особенности структуры и функции ядра ооцита
 и пыльце лилии (F –")
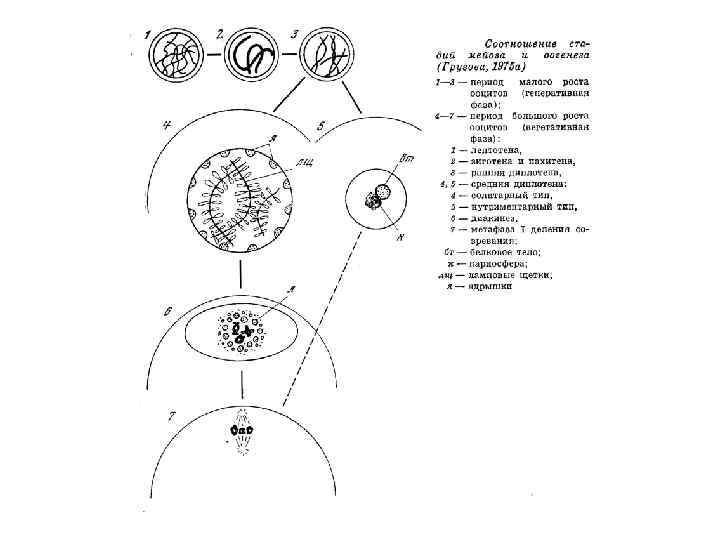
Стадии профазы мейоза в ооцитах человека (А – Е) и пыльце лилии (F – G) A. Лептотена B. Зиготена С. Пахитена D. Диплотена Е. Диакинез F. Ранняя диплотена G. Поздняя диплотена
 и")
Схема синаптонемного комплекса между гомологичными хромосомами (показано по одной хроматиде в каждой хромосоме) и образующихся путем самосборки полисинаптонемных комплексов в ядре и цитоплазме ооцита.

Последовательность событий при синапсисе и разъединении хромосом в профазе I мейоза. Полностью сформированный синаптонемальный комплекс существует на протяжении всей стадии пахитены. Схема типичного синаптонемного комплекса. Представлен небольшой участок длинного, похожего на лестницу образования с латеральными и центральным элементами и рекомбинационным узелком.

Электронно-микроскопическое изображение синаптонемных комплексов и ассоциированных рекомбинационных узелков в оогенезе Drosophila (по A. T. C. Carpenter, 1994 и Schmekel et al. 1993)

Хромосомы типа ламповых щеток в ядре ооцита тритона

Схема строения хромосомы типа ламповой щетки: транскрипционные единицы представляют собой растянутые петельные домены

Электронно-микроскопическое изображение растянутой петли хромосомы типа ламповых щеток у тритона Pleurodeles waltli. Транскрипционная единица на боковой петле. РНК транскрибируется с большой скоростью, о чем говорят многочисленные фибриллы вдоль осевой фибриллы ДНП. Масштабная линейка 5 мкм.
. А. Две гигантские петли хромосомы с")
Хромосомы типа ламповых щеток Pleurodelles waltlii (фазовый контраст). А. Две гигантские петли хромосомы с матриксом, свидетельствующем о наличии одной транскрипционной единицы в каждой. B. Пара гранулярных петель с полярным матриксом. C. Глобулярные петли у двух гомологичных хромосом. Оси петель не видны. D. Пара гомологичных хромосом с плотными петлями (d) с матриксом из мелких гранул (D) или глобул (Е).

Структура рибонуклеопротеинового матрикса в хромосомах типа ламповых щеток в оогенезе Pleurodeles waltlii. 1. Два бивалента из двух гомологичных хромосом каждый в диплотене профазы 1 -го деления созревания. 2. Организация РНП матрикса нормальной петли. Каждый РНП транскрипт состоит из частиц РНП одинакового размера (50 nm, включая 20 nm частицы золота). Транскрипты спирально расположенны вокруг оси петли. p – РНП частица, gl – глобулярная петля, gr – гранулярная петля, n – нормальная петля.

Компактизация РНП матрикса хромосом типа ламповых щеток Pleurodeles waltlii с образованием гранулярных и глобулярных петель. Глобулярные петли образуются при слиянии гранул, глобулы имеют 3 – 4 мкм в диаметре, Размер РНП частиц в гранулах постоянный (50 nm) и не зависит от длины транскрипта.
, гранулярной (b) и плотной (с) петлях. РНП частицы")
Организация РНП матрикса в нормальной (a), гранулярной (b) и плотной (с) петлях. РНП частицы и гранулы матрикса располагаются спирально вокруг оси ДНК.

Хромосомы типа ламповых щеток Pleurodeles waltlii. Схема прогрессивной компактизации РНП-матрикса при сопутствующем изгибании оси (ДНК). А. РНК-транскрипты. В. Нормальная петля с РНП-матриксом. С. Гранулярная петля. Взаимодействие РНП частиц соседних транскриптов приводит к формированию гранул. D. Глобулярная петля. Агрегаты соседних РНПтранскриптов увеличиваются в размерах. Изгибание оси петли (ах) усиливается. Е. Плотная петля. Максимальная компактизация матрикса.

Выявление транскриптов повторяющихся последовательностей ДНК на петлях ламповых щеток 4 х видов амфибий. In situ гибридизация петель Xenopus laevis (a), Notophthalmus viridescens (b); Euproctus (c) и Pleurodeles walti (d) радиоактивными пробами к. РНК тритона P. walti. к. РНК транскрибирована с повторяющихся последовательностей в присутствии Н 3 уридина и амплифицирована. Гибридизация одной пробы с хромосомами разных видов животных свидетельствует об универсальности транскрибируемых повторов на ХТЛЩ.

Локализация генов гистонов на хромосомах типа ламповых щеток в ооците саламандры Notophtalmus viridescens. Гены выявлены с помощью авторадиографии и in situ гибридизации с меченой по тритию (Н 3) к. РНК гистоновых генов.

Множественные свободные ядрышки и кариосфера в ядре ооцита тритона на стадии поздней диплотены На стадии поздней диплотены продолжается активный синтез р. РНК в экстрахромосомных ядрышках, а хромосомы стягиваются в центре зародышевого пузырька (ядро) и отделяются от остального ядерного пространства капсулой. Уровень синтеза РНК на хромосомах в это время низкий. Капсула состоит из эластичного волокнистого материала (сеть псевдомембран и мембранных структур).
")
Организация рибосомальной ДНК у разных видов организмов (D. L. Lafontaine, D. Tollervey, 2001)

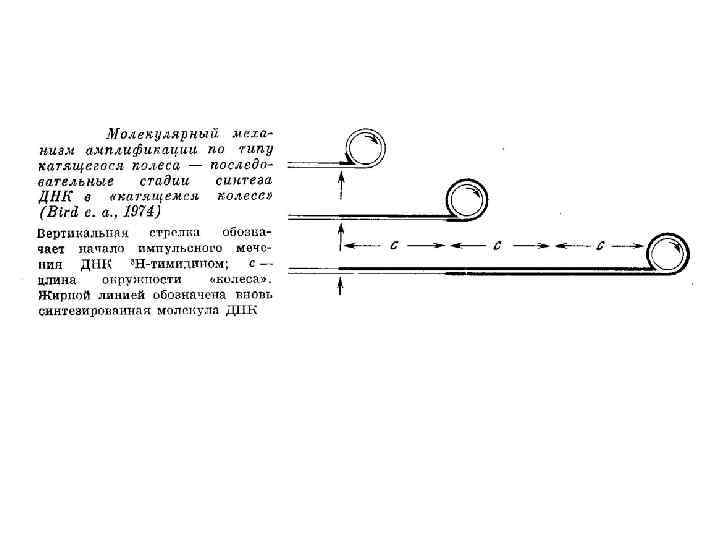
Электронно-микроскопический анализ амплифицированной рибосомальной ДНК Xenopus laevis Кольцевые молекулы r. DNA из градиента плотности Cs. Cl (слева) и репликативная единица- кольцевая молекула с хвостом (справа)

Локализация генов 5 S и 18 S - 28 S рибосомальных РНК в ядрах соматических и половых клеток домашнего сверчка Acheta domesticus. Радиоавтографы соматических интерфазных клеток (а, b), сперматоцитов (c, d) и ооцитов (e, f). Гибридизация с H 3 меченой к. РНК генов рибосомной ДНК (r. DNA) и с H 3 -меченой к. РНК генов 5 S р. ДНК

Автор презентации С. М. Ефремова
Лекция 3_ Ядро ооцита.ppt