07_Молекулярная эволюция.pptx
- Количество слайдов: 34



Особенности молекулярной эволюции 1. Скорость эволюции любого белка, выраженная через число аминокислотных замен на сайт в год, приблизительно постоянна и одинакова в разных филогенетических линиях, если только функция и третичная структура этого белка остаются в основном неизменными.
, дистанция от")
Зачем нужна аутгруппа Молекулярнофилогенетические методы используют информацию о последовательностях внешней группы (контроля), дистанция от которой для всех остальных последовательностей заведомо выше, чем от других. Таким образом дерево «укореняется» , а также внутри дерева убирается «шум»

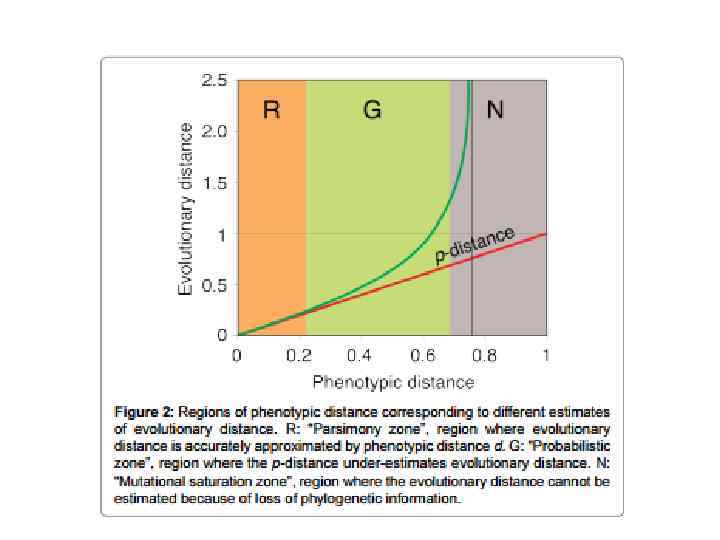
Выбор модели замен Результаты вычисления эволюционных дистанций будут отличаться в зависимости от выбранной модели замен

«Эффект тыквенного пирога» Рецепт тыквенного пирога на филогенетическом дереве креветок.

Различия между деревом генов и деревом видов

Особенности молекулярной эволюции 2. Функционально менее важные молекулы или их части эволюционируют (накапливая эволюционные замены) быстрее, чем более важные 3. Мутационные замены, приводящие к меньшим нарушениям структуры и функции молекулы (консервативные замены), в ходе эволюции происходят чаще тех, которые вызывают существенное нарушение структуры и функции этой молекулы

Yates, Sternberg, 2013. Некоторые белковые домены более устойчивы к несинонимичным заменам (т. е. в данном случае реже проявляется клинический эффект), чем другие

Hecht et al. , 2013. Ландшафт мутабильности адренергического рецептора ADRB 2 человека. Смоделированы эффекты от замены аминокислоты «дикого типа» на все остальные (красный — структура и функция меняется, зеленый — нейтральность)
 в аспарагин в металло-бета-лактамазе (E. coli)")
Мутации в активном центре Мутация аспарагиновой кислоты (120) в аспарагин в металло-бета-лактамазе (E. coli) привела изменению расстояния и перераспределению заряда между ионами цинка, что привело к невозможности расщепления антибиотиков.

Мутации, меняющие доступ к активному центру Замена изолейцина на лейцин в CYP 6 B 1 Papilio polyxenes ограничила доступ субстрата к активному центру, что привело к невозможности метаболизировать фуранокумарин.

Мутации в сайте связывания с коферментом Изменение в месте связывания с НАДФН, расположенному в удалении от активного центра, в алкогольдегидрогеназе Clostridium autoethanogenum привело к изменению в специфичности к субстрату. В некоторых случаях менялась специфичность к коферменту (НАДН вместо НАДФН)

Du et al. , 2014. У родопсинов млекопитающих частота использования различных синонимичных кодонов сильно отличается. Сильнее, чем ожидалось от мутационного процесса или дрейфа. Отбор может действовать на синонимичные замены высоко экспрессируемых генов.

Chaney, Clark, 2015. Механизм этого отбора: Более редкие кодоны дольше транслируются, и из-за этого часть белков сворачивается раньше времени в неправильную (нерабочую) структуру. Можно проверить с помощью двух флуоресцентных меток.

Duan et al. , 2003 Даже при синонимичной замене одного нуклеотида м. РНК может сильно изменить свою конформацию, что приведёт к уменьшению или увеличению её стабильности

Особенности молекулярной эволюции 4. Появлению нового в функциональном отношении гена всегда должна предшествовать дупликация гена.

Callie et al. , 2003 D 1 и D 2 допаминовые рецепторы менее родственны другу, чем некоторым серотониновым и адреналиновым. Внутри семейства D 1 произошла субфункционализация: гены дифференциально экспрессируются в различных тканях и органах
 и потери")
Niimura, Nei, 2003 Внутри обонятельных рецепторов позвоночных животных происходили множественные дупликации (амплификации) и потери генов. По сравнению с мышью, у человека значительно меньше обонятельных рецепторов.

Эволюция генных семейств Ортологи — гомологичные гены, образовавшиеся в результате видообразования Паралоги — гомологичные гены, образовавшиеся в результате дупликации

Эволюция генных семейств

Эволюция глобинов позвоночных

Филогенетическое дерево белков теплового шока hsp 70 демонстрирует согласованную эволюцию

Эволюция по модели «рождение и смерть» Возникновение новых генов путём дупликации уравновешивается псевдогенизацией.

Другие способы возникновения новых генов • Горизонтальный перенос • Перетасовка экзонов • Химеризация генов

Особенности молекулярной эволюции 5. Селективная элиминация вредных мутаций и случайная фиксация селективно нейтральных или очень слабо вредных мутаций происходят в эволюции гораздо чаще, чем положительный дарвиновский отбор полезных мутаций

Детекция отбора 1. Поиск «выметания отбором» — снижения генетической изменчивости при отборе полезной мутации из-за сцепления 2. На основании соотношения числа синонимичных и несинонимичных замен

Тест Макдональда-Крейтмана Сравнение доли синонимичных и несинонимичных точечных мутаций среди полиморфных и дивергировавших локусов. Между видами Внутри вида Синонимичные Ds Ps Несинонимичные Dn Pn Если Ds / Dn = Ps / Pn, то все замены нейтральны Если Ds / Dn > Ps / Pn, то часть замен происходила в результате отбора.

Tadjima’s D Параметр может быть оценен разными способами: 1. Как число попарных различий между образцами, где dij — число различий между двумя последовательностями, n — число последовательностей 1. Как отношение где S — наблюдаемое число отличающихся (сегрегирующих) остатков.
 D=0 => Наблюдаемая дисперсия равна ожидаемой. Нет следов отбора. 2) D<0")
Tadjima’s D 1) D=0 => Наблюдаемая дисперсия равна ожидаемой. Нет следов отбора. 2) D<0 => Редкие аллели присутствуют в небольшом количестве. Недавнее «выметание отбором» , или бутылочное горлышко, или сцепление с геном, по которому отбор, 3) D>0 => Множество аллелей в разных концентрациях. Балансирующий отбор или недавнее внезапное слияние популяций.

Индекс фиксации Fst Метод обнаружения потока генов, а не детекции отбора. — среднее число попарных различий между двумя особями из разных субпопуляций — среднее число попарных различий между двумя особями из одной субпопуляции Варьирует от 0 (свободное скрещивание между образцами) до 1(нет общей генетической изменчивости)

О чём говорят нам данные значения Tajima’s D и Fst? Liu et al. , 2015
")
Z-тест • Для пары последовательностей, сначала подсчитывается число синонимичных замен на синонимичный сайт (ds) и число несинонимичных замен на синонимичный сайт (dn), а также их дисперсия σ2(ds) и σ2(dn). • Далее нулевая гипотеза H 0: dn= ds проверяется с помощью Z-теста:

Z-тест Уровень значимости, при котором нулевая гипотеза отвергается, будет зависеть от альтернативной гипотезы HA: a) d. N ≠ d. S (тест на нейтральность), b) d. N > d. S (положительный отбор), c) d. N < d. S (очищающий отбор). Для альтернативных гипотез (b) и (c) используется односторонний тест, для (a) используется двусторонний тест.

Методы, ищущие отбор по каждому кодону • • Счетные методы Модели со случайными эффектами Модели с фиксированными эффектами Модели со смешанными эффектами
07_Молекулярная эволюция.pptx