Особенности бактериального метаболизма.ppt
- Количество слайдов: 61



Особенности бактериального метаболизма

Вопросы 1. 2. 3. 4. 5. 6. 7. 8. Основные понятия Ферменты Пути катаболизма Общая схема катаболизма (на примере катаболизма гексоз) Мембранное фосфорилирование Особенности метаболизма аэробных микроорганизмов Особенности метаболизма анаэробных микроорганизмов Брожение
 Катаболизм Анаболизм Амфиболизм")
1. Основные понятия Метаболизм (вся совокупность химических реакций в клетке) Катаболизм Анаболизм Амфиболизм

АТФ-универсальный переносчик энергии

АТФ Мембранное фосфорилирование синтез клеточных компонентов Субстратное фосфорилирование активный транспорт движение тепло, фосфоресценция АДФ

2. Структура ферментов простые Ферменты сложные Ферментный белок кофермент или простетическая группа Металлоферменты – ферменты, в состав коферментов которых входят ионы металлов (Fe, Cu, Mo, Zn, Mn)
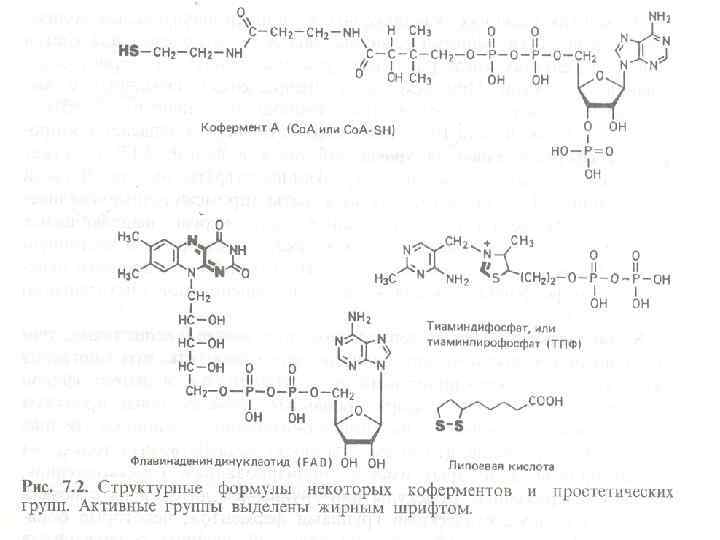

 1")
Пример классификации фермента Бетаин-гомоцистеинметилтрансфераза КФ 2. 1. 1. 5. 2 – класс (трансферазы) 1 – подкласс (переносящие одноуглеродные остатки) 1 – подподкласс (переносят в форме метила) 5 – номер конкретного фермента

Схема взаимодействия фермента с субстратом

Схема аллостерической регуляции деятельности ферментов
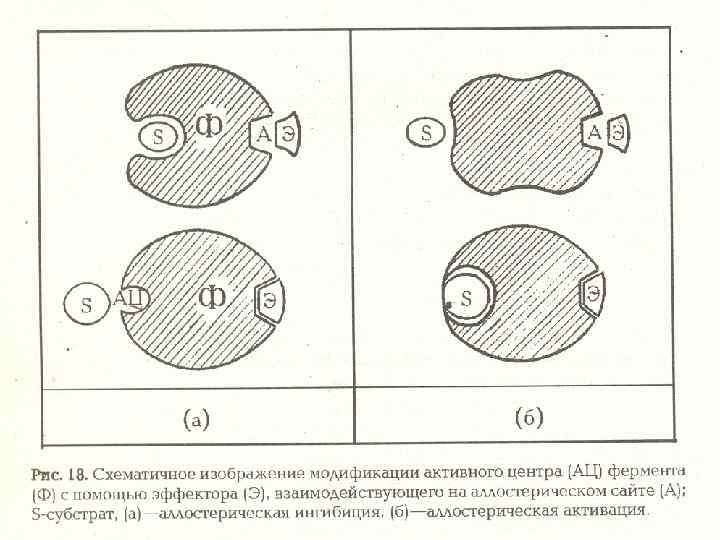
: - аэробное органического субстрата - аэробное")
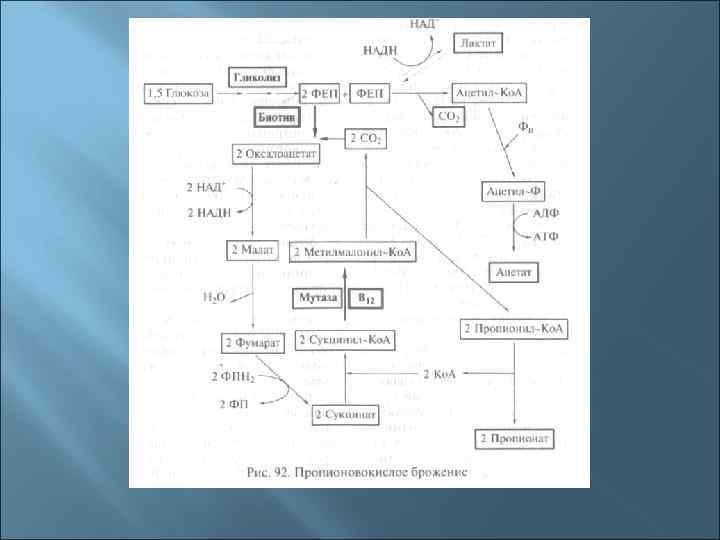
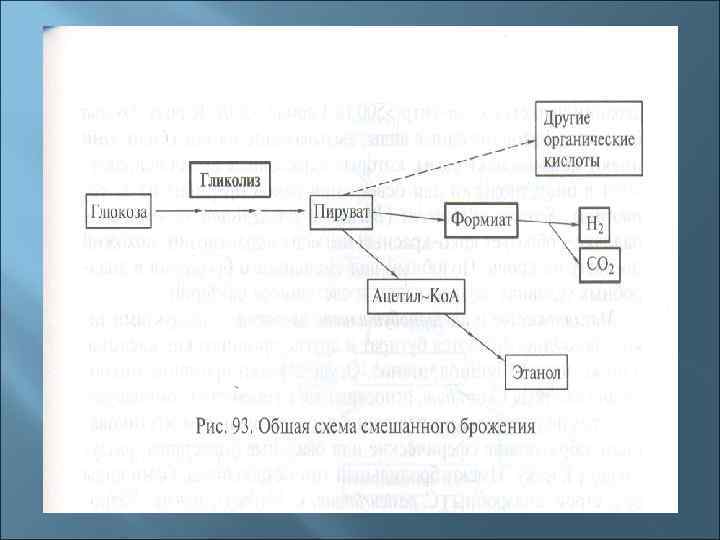
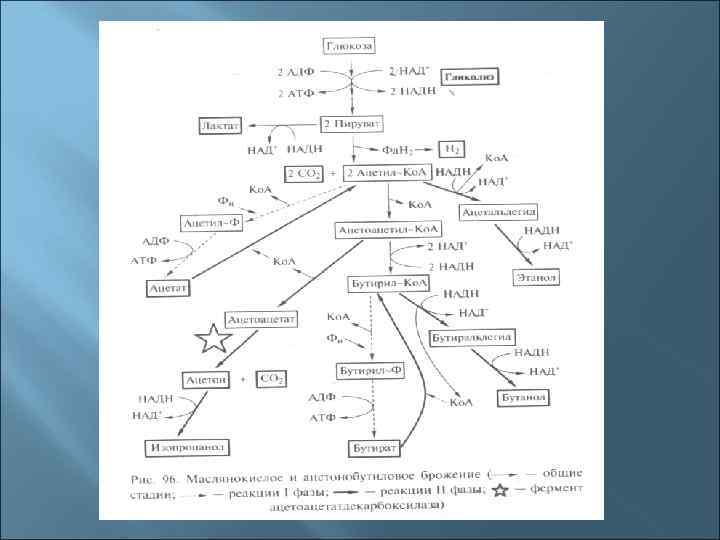
3. Пути катаболизма у прокариот 1. Окисление (дыхание): - аэробное органического субстрата - аэробное неорганического субстрата - анаэробное неорганического субстрата 2. Брожение: - молочнокислое - маслянокислое - спиртовое - пропионовокислое - муравьинокислое
 Глюкоза пируват ацетил-Ко. А окислительные")
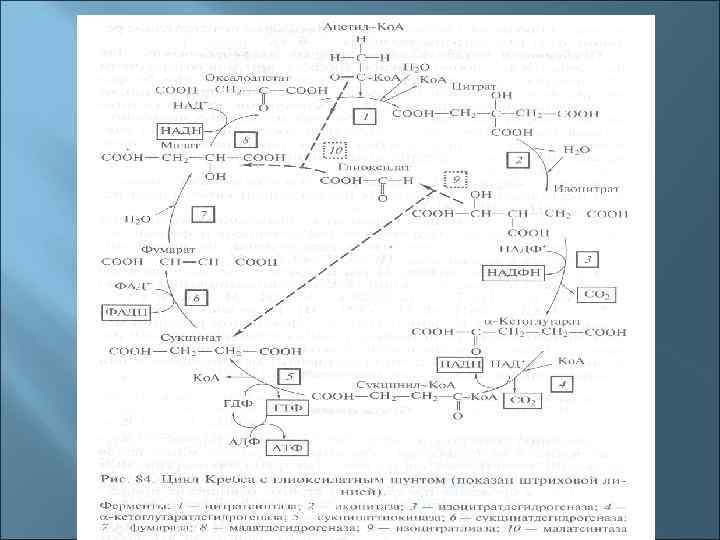
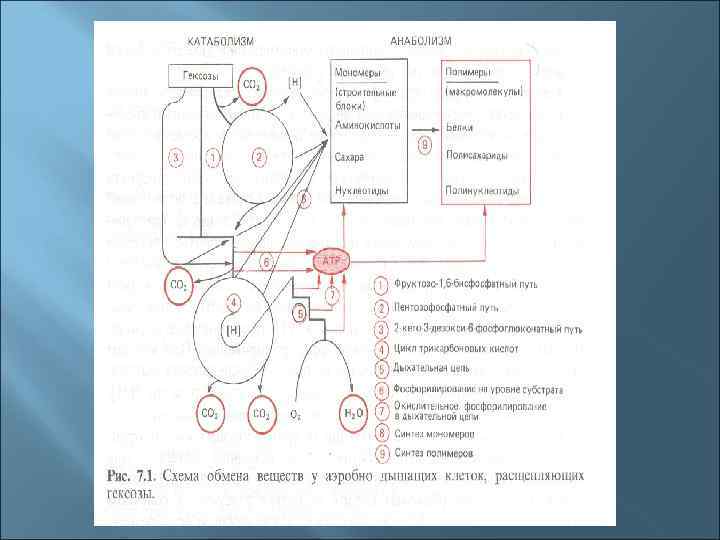
4. Общая схема катаболизма (на примере катаболизма гексоз ) Глюкоза пируват ацетил-Ко. А окислительные циклы (цикл Кребса) брожение
. 2. Пентозофосфатный путь")
Три основные пути окисления глюкозы до пирувата 1. Гликолиз (путь Эмбдена-Мейергофа-Парнаса). 2. Пентозофосфатный путь (путь Варбурга-Диккенса. Хорекера). 3. Кетодезоксифосфоглюконатный (КДФГ-путь, путь Этнера-Дудорова)

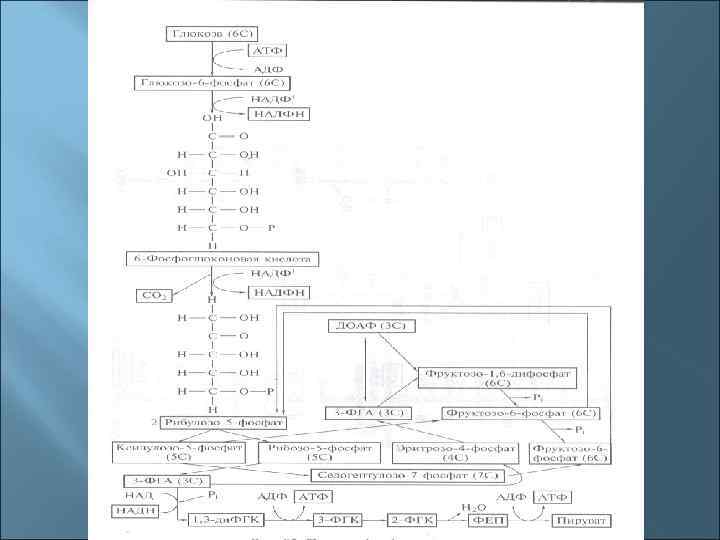
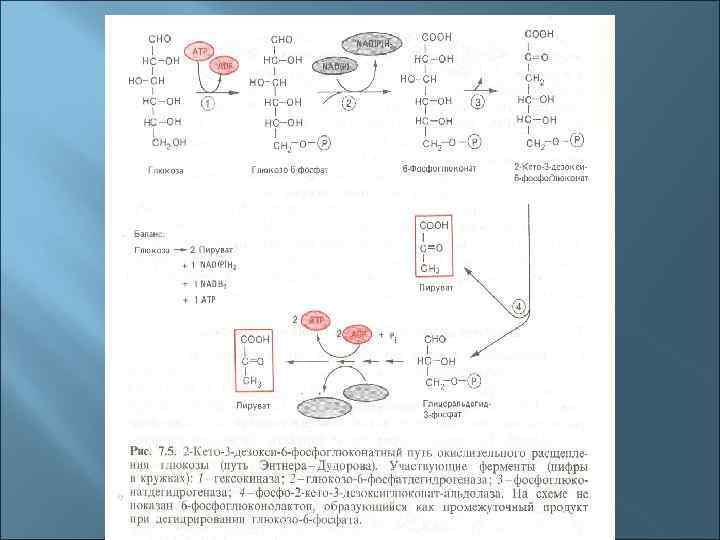

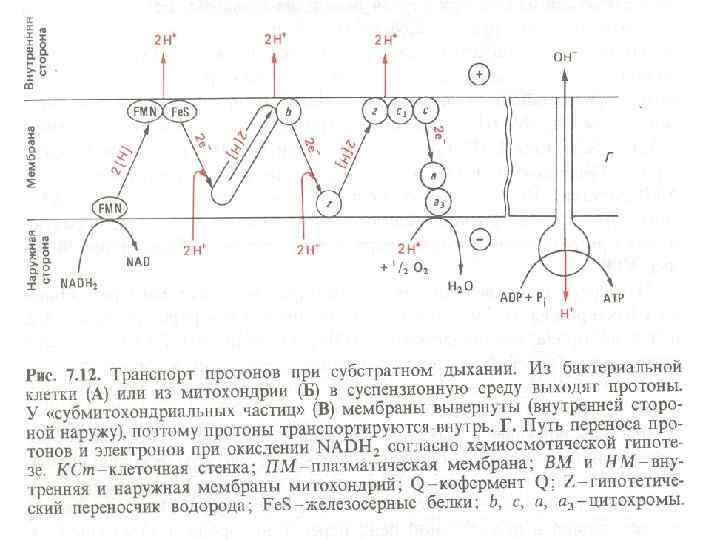
5. Мембранное фосфорилирование Это окислительное и фотосинтетическое фосфорилирование. Происходит в толще мембран в результате работы электронтранспортной (дыхательной) цепи (ЭТЦ). Отданные субстратами восстановительные эквиваленты (протоны и электроны) транспортируются через мембрану таким образом, что между внутренней и наружной сторонами мембраны создается электрохимический градиент с положительным потенциалом снаружи и отрицательным внутри. Электрохимический градиент служит движущей силой для процесса регенерации АТФ (с помощью фермента АТФ -синтазы, также расположенного в мембране).

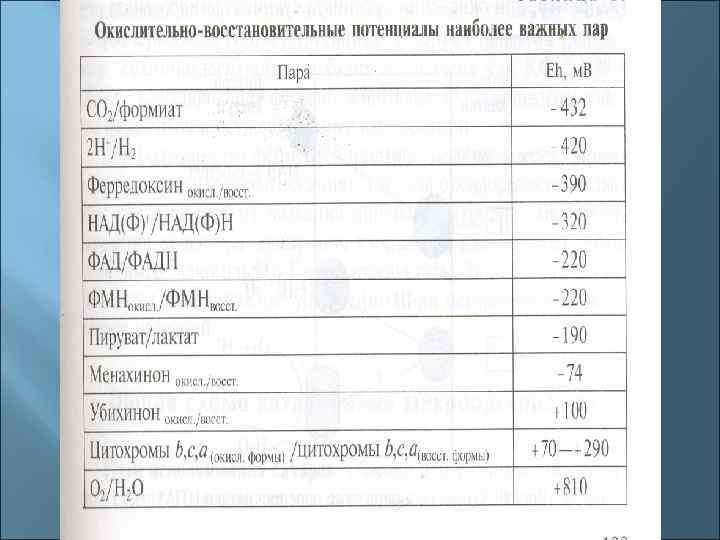
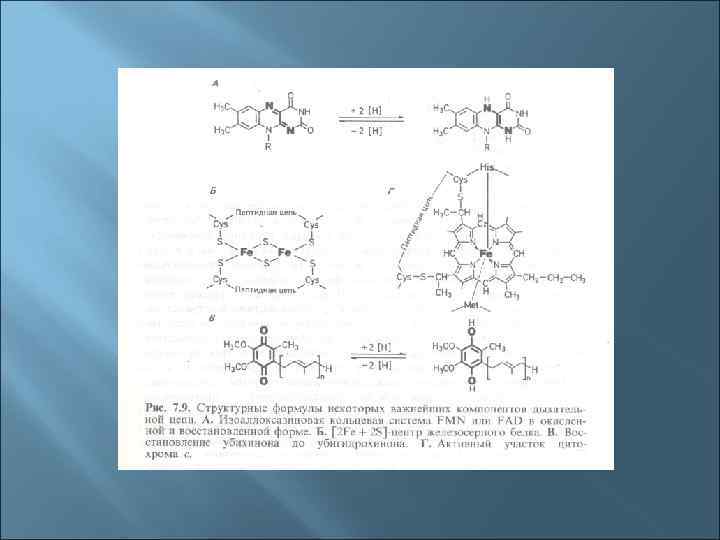
1. В состав ЭТЦ входит 3 группы ферментов: Дегидрогеназы с коферментом НАД, ФМН. Переносят протоны и электроны. Дегидрогеназы с коферментом НАД находятся в цитозоле, дегидрогеназы с коферментами-ФП находятся на внутренней стороне ЦПМ. 1. Убихиноны (активный центр - кетогруппы) находятся в толще ЦПМ, переносят протоны и электроны. 1. Цитохромы (металлоферменты, активный центр - гем) расположены в толще и на внутренней поверхности ЦПМ, переносят только электроны. Основные цитохромы бактерий - в, с, с1, а, а 3.

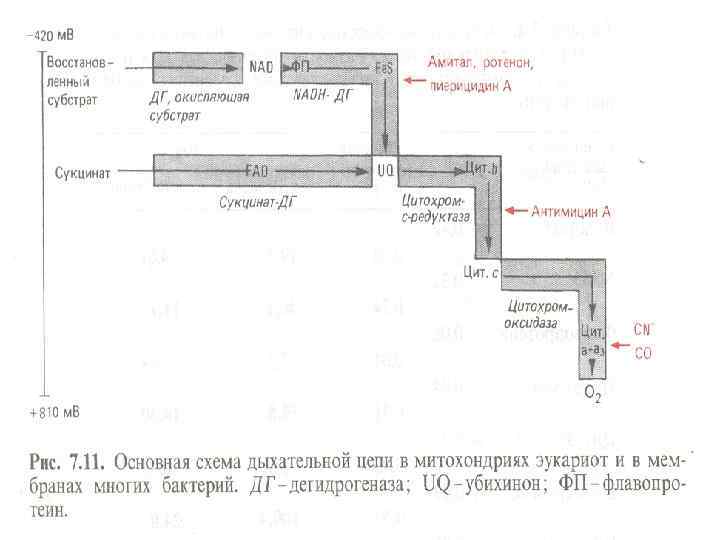
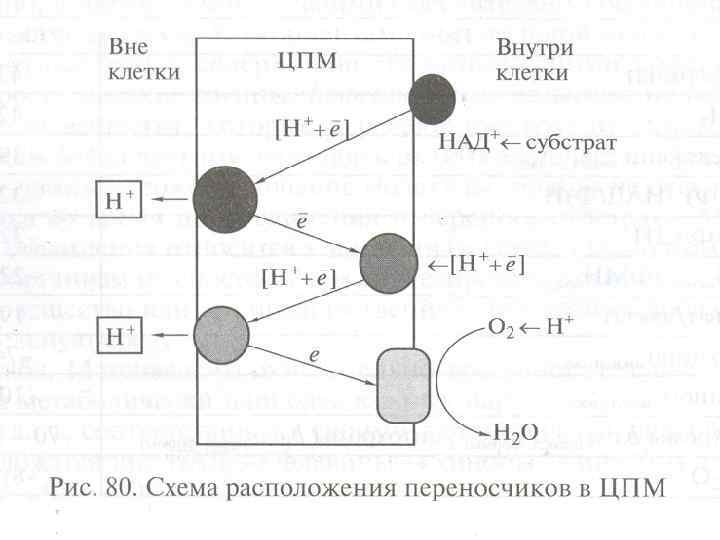

1. 6. Особенности метаболизма аэробных микроорганизмов Аэробное окисление органического субстрата - Использование многоуглеродных субстратов У большинства м/о дыхание не отличается от такового в митохондриях. Но, в зависимости от условий, м/о способны использовать альтернативные пути, синтезируя различные цитохромоксидазы. У некоторых м/о с укороченной ЭТЦ образуется больше цитохромов. При полном окислении субстрата единственным окисленным продуктом является СО 2. В качестве субстрата м/о используют сахара, белок, целлюлозу, липиды, ароматические соединения, предельные и непредельные углеводороды

НАДН Высокая аэрация ФП Цит. b 569 Убихинон, менахинон Цит. b 569 Цит. bd O 2 Цит. o Низкая аэрация Схема разветвленной ЭТЦ у E. coli
. Окисляют сахара и спирты до более простых органических веществ:")
- Неполное окисление (Acetobacter, Pseudomonas). Окисляют сахара и спирты до более простых органических веществ: Этанол ацетат Глицерин диоксиацетон Сорбит сорбоза Причины: дефекты в ферментных системах или замедление их работы из-за неоптимальности условий культивирования
 СН 4 СН 3 ОН НСООН СО 2 Диоксиацето-")
- Использование одноуглеродных субстратов (метилотрофия) СН 4 СН 3 ОН НСООН СО 2 Диоксиацето- Рибулозомонофос- Сериновый Цикл Кальвина новый цикл фатный цикл путь С 3 -соединения Биомасса

Метилотрофы относятся к филуму В 12: рода Methylomonas, Methylobacter, Methylococcus, Methylocystia и др. Окисляют, как правило, только метан (реже и хуже метанол). Обладают хорошо развитой системой мезосом (ламеллярных). Ключевой фермент окисления метана – метанмонооксигеназа (ММО), чаще мембрансвязанная. Метилотрофы плохо культивируются (часто растут со спутниками олиготрофными Hyphomicrobium) Рассматриваются как перспективный источник пищевого белка.

2. Аэробное окисление неорганического субстрата Процесс хемолитоавтотрофии открыт в 1887 г. С. Н. Виноградским

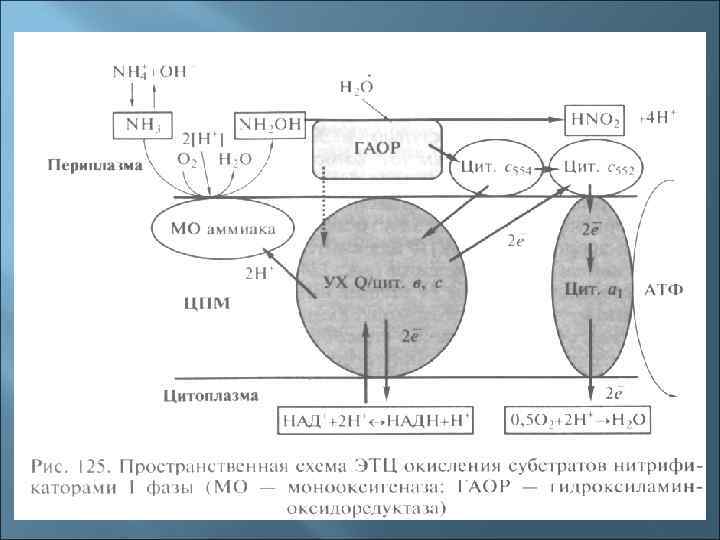
- Нитрификаторы 1 и 2 фазы NH 3 + ½ O 2 NН 2 OН N Н 2 O Н + Н 2 О НNO 2 + 4 Н+ + 4 e(Nitrosomonas, Nitrosococcus) NO 2 - + Н 2 O NO 3 - + 2 Н+ + 2 е(Nitrobacter, Nitrococcus) У нитрификаторов обнаружен механизм обратного переноса электронов с затратой АТФ для синтеза восстановительных эквивалентов (НАДН), на что тратится большое количество субстрата, поэтому нитрификаторы медленно растут.

- Тионовые бактерии и серобактерии S 2 - + 2 O 2 SO 42 - S 0 + H 2 O + ½O 2 SO 42 - + 2 H+ S 2 O 32 - + H 2 O + 2 O 2 2 SO 42 - + 2 H+ (Thiobacillus, Sulfolobus, Beggiatoa, Thiotrix и др. )

-Металлоокисляющие м/о Наиболее изучены железоокисляющие бактерии (С. Н. Виноградский, М. бейеринк, впервые выделил Х. Г. Эренберг в 1836 г. ) Fe 2+ + 4 H+ + O 2 4 Fe 3+ + 2 H 2 O (Acidithiobacillus ferrooxidans, Galionella ferruginea). Mn 2+ + ½O 2 + H 2 O Mn. O 2 + 2 H+ (Sulfolobus acidocaldarus). ЭТЦ короткая, электроны от Fe 2+-оксидоредуктазы восстанавливают цитохром аа 3 (1 пункт сопряжения, мало энергии). Могут быть окислены металлы с переменной валентностью Cu 2+ U 4+ As 3+ Sb 3+ Mo 4+ Zn 2+ Co 2+ Pb 2+ и др.

- Водородные бактерии 6 Н 2 + 2 O 2 + CO 2 (CH 2 O) + 5 H 2 O (рода Pseudomonas, Alcaligenes, Paracoccus, Nocardia, Mycobacterium). Н 2 НАДН ФП УХ Цит. в, с, а О 2
")
-Карбоксидобактерии CO + 2 ½ O 2 + H 2 O (CH 2 O) + 6 CO 2 (Pseudomonas carboxidovorans, P. carboxidoflava, Acinetobacter sp. , Alcaligenes carboxydus) Карбоксидобактерии похожи на водородные, часто эти активности пересекаются. У карбоксидотрофов обнаружен фермент СОдегидрогеназа (обычно НАД+-зависимая), содержащая Мо и Fe. S-кластер. Иногда гены, кодирующие этот фермент, находятся в плазмидах.

7. Особенности метаболизма анаэробных микроорганизмов Анаэробное дыхание – энергодающий процесс, в котором конечным акцептором электронов служит окисленное органическое или неорганическое вещество (отличное от кислорода). Сопряжено с функционированием ЭТЦ. Виды анаэробного дыхания подразделяются по используемому конечному акцептору электронов.


Нитратное дыхание NO 3 - NO 2 - NO N 2 У денитрификаторов донор электронов – органическое вещество, ЭТЦ сформирована полностью (3 пункта сопряжения), конечным ферментом является мембрансвязанные нитрит- и нитратредуктазы (их синтез индуцируется только в анаэробных условиях и в присутствии нитрата). Денитрификаторы – факультативные анаэробы.

Сульфатное дыхание Конечным акцептором электронов служит сульфат. Донорами электронов могут служить формиат, ацетат, водород, бутират, лактат, высшие жирные кислоты, этанол и др. Сульфатредукторы – облигатные анаэробы. Спорообразующие – рр. Desulfotomaculum, Desulfonispora, Desulforosinus. Неспоровые – рр. Desulfobacter, Desulfococcus, Desulfobulbus, Desulfonema. Археи – Archaeoglobus fulgidus.

Самый экзотичный вид анаэробного дыхания – «железное дыхание» . Акцептор электронов Fe 3+, донор ацетат. Представители этой группы м/о: сем. Geobacteriaceae, рр. Bacillus, Clostridium, Escherichia, Pseudomonas, Serratia. Считается, что «железное дыхание» самое древнее. К фумаратному дыханию способны все м/о, имеющие ЭТЦ с сукцинатдегидрогеназой (энтеробактерии, вибрионы, пропионовые бактерии). ЭТЦ короткая (1 пункт сопряжения).


При карбонатном дыхании конечным акцептором электронов служит СО 2 или СО. Донором электронов – водород, ацетат, некоторые спирты, одноуглеродные субстраты (формиат, метиламин, ди- и триметиламины). К этой группе м/о относятся метаногены (самая большая группа архей из филума Euryarchaeota). Классы Methanobacteria, Methanococci, Methanopyri Обычно являются последним звеном в анаэробных пресноводных местообитаниях. Это древнейшие организмы в истории Земли.

Схемы реакций, катализируемых конечным ферментом дыхательной цепи метаногенов 4 Н 2 + СО 2 СН 4 + 2 Н 2 О 4 СООН СН 4 + СО 2 + 2 Н 2 О СН 3 ОН + Н 2 СН 4 + Н 2 О 4 СН 3 NH + 2 H 2 O 3 CH 4 + CO 2 + 4 NH 3 2(CH 3)2 NH + 2 H 2 O 3 CH 4 + CO 2 + 2 NH 3

Метаногенные археи занимают важное место в природных экосистемах. Метаногенез активно идет на рисовых полях, источники метана – рубец жвачных животных, кишечник лошадей и других млекопитающих (примерно 70% людей имеют метаногены в кишечной биоте). Еще один источник образования метана – искусственные сооружения для очистки стоков и получения биогаза (анаэробные реакторы и метантенки). В таких установках обычно работают микробные сообщества, где метаногены – конечное звено пищевой цепи разложения сложных органических соединений.

8. Брожение По определению Л. Пастера, брожение – это жизнь без кислорода. В более узком смысле – это бескислородные превращения пирувата, полученного в реакциях одного из путей преобразования сахаров (гликолиза, пентозофосфатного, КДФГ-пути).
 1, 3 -Бисфосфоглицерат 3 -Фосфоглицерат (фермент фосфоглицераткиназа) 2)")
Наиболее важные реакции субстратного фосфорилирования 1) 1, 3 -Бисфосфоглицерат 3 -Фосфоглицерат (фермент фосфоглицераткиназа) 2) Фосфоенолпируват Пируват (фермент пируваткиназа) 3) Ацетилфосфат Ацетат Бутирилфосфат Бутират (фермент ацетаткиназа)

СО О Р СО НС ОН СН 2 О НС Р АДФ 1, 3 -Бисфосфоглицерат АТФ ОН ОН СН 2 О Р 3 -Фосфоглицерат
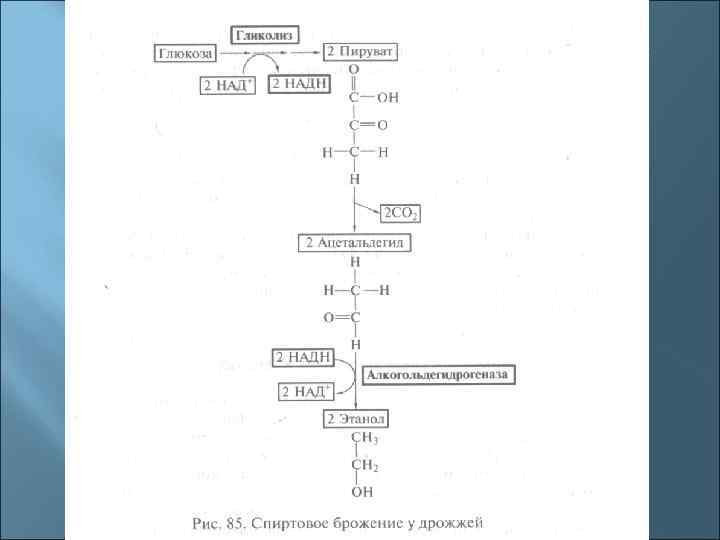

Осуществляют спиртовое брожение р. Saccharomyces р. Schizosaccharomyces р. Sarcina Zymomonas mobilis (агава – пульке - текила) Leuconostoc mesenteroides Некоторые энтеробактерии и клостридии
")
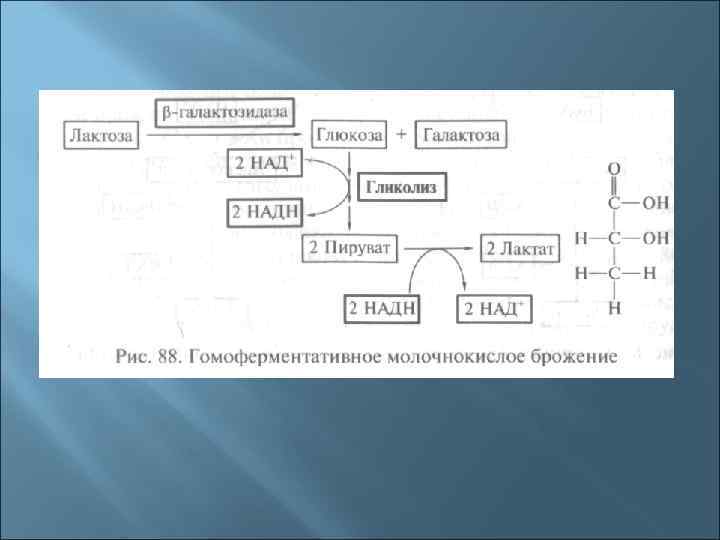
Осуществляют гомоферментативное молочнокислое брожение р. Streptococcus р. Pediococcus р. Lactobacillus (L. lactis)
 р. Bifidobacterium")
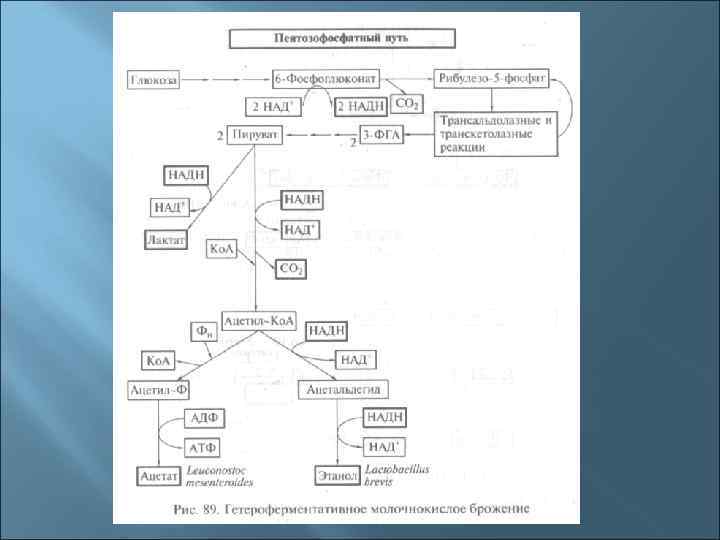
Осуществляют гетероферментативное молочнокислое брожение р. Leuconostoc р. Lactobacillus (L. brevis, L. acidophilis) р. Bifidobacterium Лактолин, низин, бревин, диплококкцин
Особенности бактериального метаболизма.ppt