d3ce7c91aa6e58bcb122a69298548792.ppt
- Количество слайдов: 137



ОСНОВЫ БИОЛОГИИ РАЗВИТИЯ ЛЕКЦИИ 10 часов Практические занятия 4 часа ауд. 216, 220



• Биология развития - наиболее объемная из всех биологических дисциплин. Это также одна из самых своеобразных наук, это наука о становлении, а не о существовании.

• В настоящее время биология развития представляет одну из наиболее захватывающих и быстро прогрессирующих областей биологической науки, приближающейся к пониманию молекулярных механизмов развития живых организмов. • Ей принадлежит объединяющая роль в отношении разных биологических наук

Связь с другими науками Биология развития физиология Биология клетки анатомия генетика иммунология Эволюционные и биоэкологически е исследования

Задача курса Задачей курса биологии индивидуального развития является - изучение закономерностей, процессов и механизмов онтогенеза. Изучение эмбрионального развития организмов стало необходимым для понимания любой другой области биологии.

Отличия биологии развития от других биологических дисциплин Основное отличие биологии индивидуального развития от физиологии, биохимии, цитологии и других биологических дисциплин в том, что они изучают статические, относительно неизмененные структуры, либо обратимые циклические процессы. Основное же свойство эмбриональных процессов – их необратимость, образование сложных и совершенных природных систем. Причем столь сложная организация возникает исключительно за счет внутренних факторов, почти не требуя внешних источников информации или управления.

Нарастание связей эмбриологии с цитологией, генетикой, молекулярной биологией привело к возникновению в ХХ веке новой комплексной науки – биологии развития. Важным этапом ее формирования стало развитие прикладных направлений исследований. В 1973 г Л. Шеттлз извлек предовуляторную яйцеклетку из яичника бесплодной женщины и оплодотворил ее сперматозоидами мужа. Было положено начало техники получения и пересадки зародыша из пробирки (экстракорпоральное оплодотворение).

В 1978 г в Великобритании в результате успешной пересадки зародыша человека на стадии 8 бластомеров после 2, 5 суточного культивирования родился первый в мире пробирочный ребенок весом 2700 г –Луиза Браун. Пересадка зародышей, зачатых в пробирке в матку –составляет основу лечения бесплодных браков.

Эмбриологические знания и методы имеют непосредственное приложение в: рыбоводстве птицеводстве шелкопрядстве животноводстве (искусственное осеменение рыб, крупного рогатого скота, инкубирование яйцеклеток ценных пород скота в матке малоценных пород). .

Эмбриологические исследования -неотъемлемая часть любых природоохранных мероприятий. Наиболее важные для человечества приложения эмбриологии относятся к области медицины. Уже сегодня аномалии внутриутробного развития человека считаются международной проблемой, так как проявляются в более или менее опасных формах у 1% новорожденных. Эмбриологические подходы важны также для понимания тканевых заболеваний взрослого организма. Именно эмбриологические знания привели в свое время Мечникова к открытию фагоцитоза, теперь помогают ученым в разработке проблем рака, исследованию иммунных реакций организма и многих смежных проблем Очень большим может быть ее вклад в развитие трансплантационной хирургии.

. Когда мы до конца поймем закономерности развития сложных многоклеточных структур - можно будет ставить вопрос об искусственном культивировании тканей и даже органов человека. Успешное развитие работ по генной инженерии невозможно без участия эмбриологов. Широко используются эксперименты по клонированию (созданию генетических копий), созданию трансгенных животных и растений. (Овечка Долли) Открытие в 1999 г эмбриональных стволовых клеток явилось третьим по значимости событием в биологии 20 го века после открытия двойной спирали ДНК и расшифровки генома человека.

• Об актуальности и достижениях этой комплексной науки -биологии развития - свидетельствует вручение Нобелевских премий за многолетний творческий труд плеяде ученых: • Исследования последних десятилетий были направлены на выяснение генетических основ развития (Нобелевская премия 1995), механизмов регуляции клеточного цикла (Нобелевская премия 2001) и програмируемой гибели клеток (апоптоза) – (Нобелевская премия 2002).

Краткая история эмбриологии XX-XXI вв. : молекулярные механизмы онтогенеза Генетический контроль раннего эмбрионального развития (Нобелевская премия, 1995). Открытие ключевых регуляторов клеточного цикла (Нобелевская премия, 2001). Э. Льюис (19182004) К. Нюслейн. Э. Вейсхаус Фолхард (1942) (1947) Л. Хартвелл (1939) Т. Хант (1943) П. Нёрз (1947) С. Бреннер (1939) Р. Хорвитц (1943) Дж. Салстон Открытия, касающиеся генетической регуляции развития органов и программируемой клеточной гибели (Нобелевская премия, 2002).

Краткая история эмбриологии XX-XXI вв. : прикладные исследования эмбриогенеза Репродуктивные технологии Экстракорпоральное оплодотворение (Р. Эдвардс, Нобелевская премия, 2010 и П. Степто). Р. Эдвардс (1925) Технология клонирования, эмбриональные стволовые клетки (Нобелевская премия, 2012). Исследования (60 -е гг. ХХ века) Дж. Гердона по пересадке ядер положили начало клонированию (появление в 1996 г. знаменитой овцы Долли). В 2006 г. Ш. Яманака экспериментально доказал, что для перехода соматической клетки в плюрипотентное состояние необходимо наличие всего четырёх транскрипционных факторов. Дж. Гердон (1927) Ш. Яманака (1962)

СТРОЕНИЕ И РАЗВИТИЕ МУЖСКИХ ПОЛОВЫХ КЛЕТОК лекция 1

Индивидуальное развитие животных имеет ациклический поступательный характер. Обычно онтогенез подразделяется на ряд стадий, каждая из которых имеет свои функциональные и морфологические особенности и создаёт основу для дальнейшего развития. Последовательная смена стадий развития во времени отражает закономерное изменение программ развития: активацию одних и выключение других генетических систем управления развитием.

• Ранние стадии развития имеют универсальный характер. • Огромное морфологическое разнообразие этих стадий у разных животных достигается на основе комбинаций ограниченного, в сущности, числа клеточных механизмов. • Особенности раннего онтогенеза во многом определяются свойствами женских гамет.

• Для понимания процессов, происходящих в раннем развитии, необходимо иметь представление о гаметогенезе, в ходе которого происходит формирование половых клеток, обязательных для обеспечения индивидуального развития животных.

• Гаметогенез - это процесс формирования половых клеток, способных при определенных условиях обеспечить воспроизводство новых особей. • Клетки, потомки которых дают начало исключительно гаметам, называются примордиальными (перевод с латинского - начало, возрождение) или первичными половыми клетками (ППК). .
,")
ППК достигают зачатка гонад, которые они заселяют активным путем, обладая амебоидной подвижностью (млекопитающие), либо пассивно, перемещаясь с током крови (рептилии, птицы). В период миграции они активно митотически делятся, благодаря чему численность популяции ППК возрастает

ППК могут нормально дифференцироваться в половые клетки только в том случае, если попадут в половые валики и вступят в контакт с соматическими клетками гонад, получая необходимую информацию для последующей дифференцировки в оогонии или сперматогонии. Если же на своем пути ППК оказываются в окружении других зачатков и не достигают гонад, то либо гибнут, либо становятся источником тератом.

• Зачатки мужских и женских гонад состоят из корковой и мозговой области. В случае развития семенников или яичников возникают различия в положении половых клеток: у самцов они располагаются в мозговой, у самок в корковой зоне. .

Первичные половые клетки Закладка половых клеток происходит на ранних этапах эмбриогенеза экстрагонадно. С момента отделения от линии соматических клеток и до вселения в гонаду это первичные половые клетки (ППК). Мужские и женские ППК неотличимы. Различия в их строении становятся заметными лишь при дифференцировке половой железы. ППК заселившие зачатки гонад –гонии: мужских –сперматогонии, женских - оогонии. Этап размножения. Вступившие на путь делений созревания клетки называют цитами (соответственно сперматоциты, ооциты)
 •")
Отличия половых клеток от соматических: • Гаплоидный набор хромосом (соматические 46, половые 23) • тотипотентность (способность формировать любые (все) органы и ткани организма); • изменённое ядерно-плазменное отношение: пониженное у яйцеклеток (объём цитоплазмы увеличен вследствие накопления питательных веществ) и высокое у сперматозоидов (из-за малого количества цитоплазмы); • различный уровень метаболизма: яйцеклетка находится в состоянии депрессии, а у сперматозоидов нормальный метаболизм и вовсе исключается; • высокая степень специализации (жгутик, оболочки яйцеклеток); • отсутствие способности к неиндуцированному митозу.

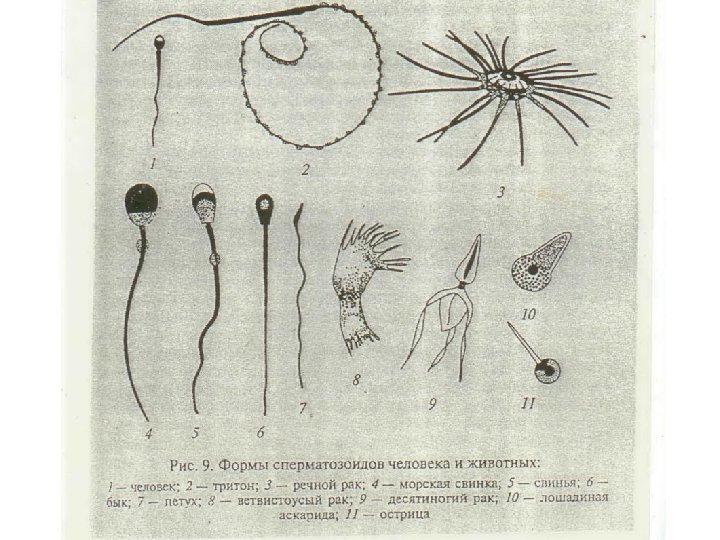
. . Особенности строения и развитие мужских половых клеток Первые описания подвижных живчиков в семени животных принадлежат И. Гаму и А. Левенгуку. Термин введен в научную литературу К. Бэром(1817 г). Окончательное решение вопроса о природе мужских половых клеток принадлежат Келликеру(1817 -1905 гг). У подавляющего боль-ва видов сперматозоиды жгутиковые, для них характерно наличие головки с ядром, шейки, вставочного отдела и жгутика. Однако известны некоторые группы животных (круглые черви, членистоногие) у которых спермии не имеют жгутиков. Спермии у них округлой или звездчатой формы, могут перемещаться с помощью псевдоподий или остаются неподвижными

Строение спермия: А • гаплоидное ядро – содержит генетический материал; • двигательная система – перемещение спермия; • акросома – используется при оплодотворении. 1 Я 2 Ц М Ан Ферменты акросомы: гиалуронидаза (растворяет 3 блестящую оболочку яйцеклетки) и трипсин (нарушает целостность фолликулярной оболочки). Части спермия: • головка, 1: акросома (А), ядро (Я) 4 • шейка, 2: центриоли (Ц) • промежуточный отдел, 3: митохондрии (М) 1 – акросома; 2 – головной колпачок; 3 – ядро; 4 – цитоплазма; 5 – проксимальная центриоль; 6 – осевые нити; 7 – дистальная центриоль • хвост: главный(4), и концевой(5) отделы: аксонема (Ан) и дуплеты микротрубочек. 5
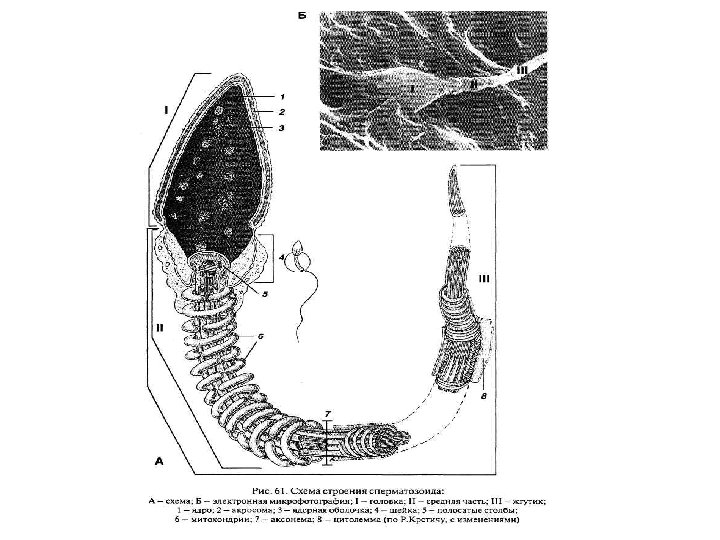

Строение жгутика спермия: • аксонема – двигательная основа. Начинается от дистальной центриоли. 2 центральные и 9 пар периферических фибрилл, связанных спицами; • митохондриальная спираль – 12– 15 витков митохондрий; Белки аксонемы: поперечный срез • тубулин – образует микротрубочки; • динеин – АТФ-аза. сборка Б – строение аксонемы (2+9) В– расположение тубулина (13+11) тубулин разрушение Отсутствие динеина (синдром Картедженера): стерильность мужских особей, подверженность респираторным инфекциям, 50 % вероятность situs inversus totalis (триада). Mov_2

В концевом отделе хвоста элементы цитосклета исчезают, дуплеты аксонемы распадаются на 18 одиночных микротрубочек. Скорость движения 2– 5 мм/мин. Реотаксис – движение против тока секрета. Направление движения: вперед вверх или вниз, вращение вокруг собственной оси. Размеры сперматозоидов разные: 200 мкм у воробья, 100 мкм у морской свинки, у крокодила 20 мкм, у человека 60 мкм.

Развитие и созревание сперматозоидов Для всех животных характерно: 1. Мужские половые клетки никогда не развиваются в одиночку и растут в виде клона синцитиально связанных клеток, где все клетки оказывают друг на друга влияние. 2. У бол-ва животных и человека в процессе сперматогенеза принимают участие вспомогательные соматические клетки (клетки Сертоли) 3. Сперматогенез у мужчин начинается только после полового созревания и затем непрерывно продолжается в эпителиальной выстилке извитых семенных канальцев, находящихся в мужских половых железах - семенниках При крипторхизме (семенники не опускаются из брюшной полости в мошонку) мужские особи, достигшие половой зрелости, стерильны. Однако у ряда млекопитающих (киты, слоны) семенники всю жизнь находятся в брюшной полости, что не сказывается на репродуктивной способности.
")
Строение семенников: Семенник – парный дольчатый орган, который разделен на дольки (250300 в каждом) за счет ответвлений, отходящих от белковой оболочки семенников. В каждой дольке 3– 4 извитых канальца (50 см в длину и 200 мкм в диаметре) в которых происходит сперматогенез. Сперматогенез человека - 70 дн. На 1 г веса яичка образуется 107 млн спермиев в сутки. Длительность жизни спермия половых путях женщины 1– 3 дня. схема строения Путь семенной жидкости: извитые семенные канальцы – прямые семенные канальцы – сеть семенника – 10– 20 выносящих канальцев – выносящий каналец придатка – семявыносящий проток – мочеиспускательный канал.

Строение извитого семенного канальца интерстициальное пространство слой клеток Сертоли соединительно-тканная основа; половые клетки (сперматогенный эпителий).

Поперечный срез через сменник
: • интерстициальное пространство – вырабатывающие тестостерон клетки Лейдига, кровеносные")
Строение извитого семенного канальца (1): • интерстициальное пространство – вырабатывающие тестостерон клетки Лейдига, кровеносные и лимфатические сосуды, нервные волокна. соединительно-тканная основа;

• слой клеток Сертоли – • опора, питание, упорядочение. Непролиферируют, формируют гемато-тестикулярный барьер, продуцируют антимюллеровский гормон (обеспечивает развитие по мужскому фенотипу, гоноциты в сперматогонии, создание необходимой концентрации андрогенов, до полового созревания ингибирует мейоз • • •
: Сперматогенный эпителий: • сперматогонии • сперматоциты I порядка •")
Строение извитого семенного канальца (1): Сперматогенный эпителий: • сперматогонии • сперматоциты I порядка • сперматоциты II порядка • сперматиды • сперматозоиды

При сперматогониальных делениях цитогенез не доходит до конца и формируется синцитий. Клетки сообщаются посредством цитоплазматических (фузом). мостиков Их наличие обеспечивает cинхронность развития половых клеток, равномерное распределение продуктов экспрессии генов между ними.
: схема 40 строения Объектив х 100 Представлена")
Строение извитого семенного канальца Объектив х (2): схема 40 строения Объектив х 100 Представлена схема строения извитого семенного канальца человека и микропрепарат извитого семенного канальца крысы (разные увеличения).
.")
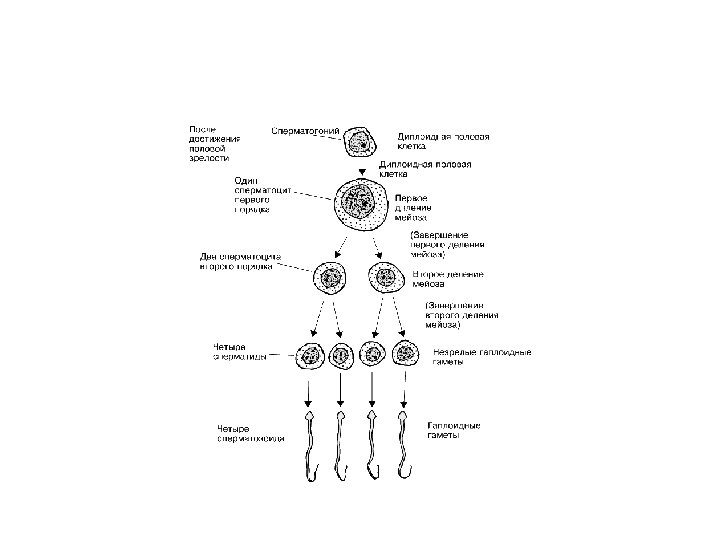
Стадии сперматогенеза: • Стадия размножения. Деление ППК и образование сперматогониев А 1 (стволовая клетка). Самопроизводство А 1 и образование сперматогониев А 2, затем А 3, А 4 и промежуточных сперматогониев, митоз которых приводит к образованию сперматогониев В. • Стадия роста. Профаза мейоза в сперматоците I порядка. Готовятся к делениям созревания • Стадия созревания. Два последовательных деления мейоза. В результате первого деления из сперматоцита I порядка образуются 2 сперматоцита II порядка. Из них в результате второго деления образуются 4 сперматиды. • Стадия формирования. Сперматиды преобразуются в сперматозоиды (спермиогенез). Длительность сперматогенеза человека – 74 дня. Скорость образования: 100 млн/час
")
Генетическая схема сперматогенеза: • Стадия размножения. Сперматогониальные митотические деления. Их число невелико (1– 14) и генетически детерминировано. • Стадия роста. Сперматоциты растут, в ядрах идет редупликация ДНК, образуются тетрады хромосом. • Стадия созревания. В сперматоцитах I порядка диплоидное число хромосом, в сперматидах – гаплоидное. При оплодотворении происходит соединение ядерного материала мужской и женской половых клеток. Зигота несёт диплоидный набор хромосом. • Стадия формирования. В ходе спермиогенеза не происходит количественного изменения числа хромосом или ДНК. А – стадия размножения; Б – стадия роста; В – стадия созревания; Г – стадия формирования

Стадия формирования • Ядро уплотняется, хроматин конденсируется и становится генетически инертным. • Перемещение органелл клетки: аппарат Гольджи уплотняется, прижимается к ядру и формирует акросому. Центриоли смещаются на противоположный от неё полюс, располагаясь одна ближе к ядру, другая дальше. • Из дистальной к ядру центриоли начинает расти жгутик. Вокруг основания жгутика в виде спиралей располагаются митохондрии. Проксимальная центриоль участвует в формировании веретена первого деления дробления. • Цитоплазма отторгается, зрелый сперматозоид практически ее лишен. Разрушаются фузомы.
. Декапептид. • Гипофиз: лютеинизирующий (лютропин, ЛГ) и")
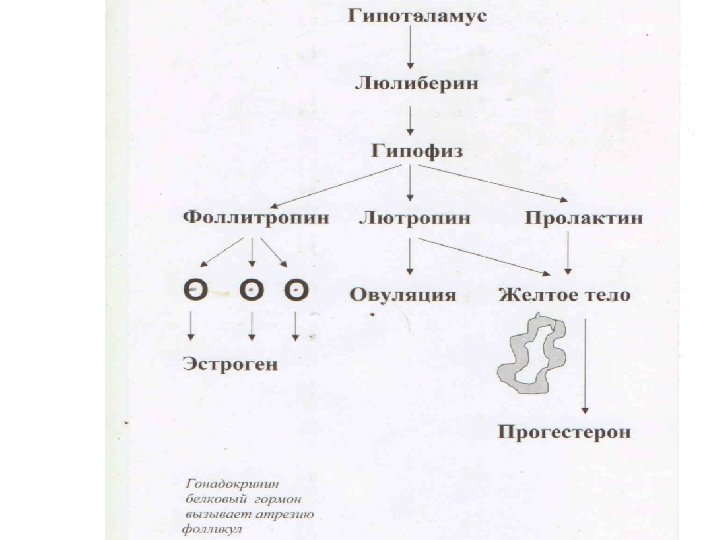
Нейрогуморальная регуляция сперматогенеза • Гипоталамус: гонадотропин (гонадотропинрилизинг-гормон). Декапептид. • Гипофиз: лютеинизирующий (лютропин, ЛГ) и фолликулостимулирующий (фоллитропин, ФСГ) гормоны. Оба – гликопротеины. • клетки Лейдига: андрогены (тестостерон, 4 -10 мг/день), эстрогены (17 -эстрадиол). Стероидные гормоны. • клетки Сертоли: 5 -дигидротестостерон, 17 эстрадиол (оба стероиды) , андрогеновый рецептор, андроген-связывающий белок, ингибины, активины, антимюллеровский гормон, окситоцин (все – пептиды). Семенная жидкость содержит фруктозу, простагландины, фибриноген (питание гамет и коагуляция спермы), даёт щелочную реакцию, обеспечивающую подвижность спермиев. Образуется в семенных пузырьках, простате, бульбо-уретальных железах под действием тестостерона. Как и все цитодифференцировки сперматогенез осуществляется под генетическим контролем.
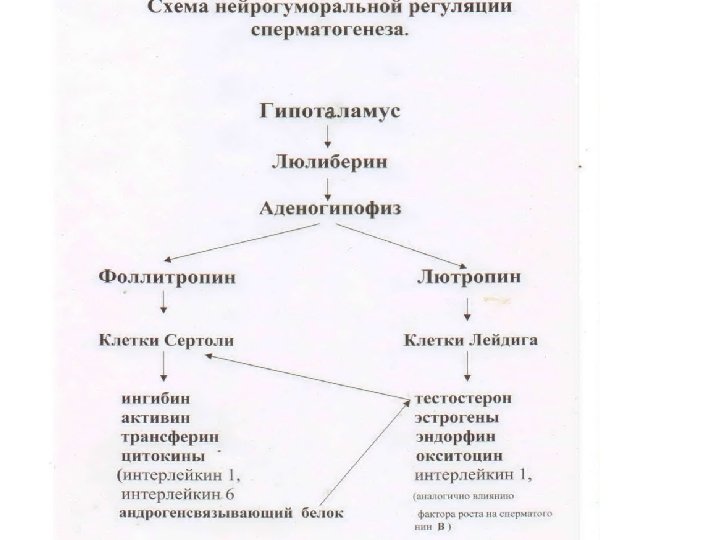

• Гормоны ингибин и активин являются физиологическими антагонистами. Ингибин, по принципу отрицательной обратной связи, подавляет секрецию фоллитропина в аденогипофизе и, таким образом, тормозит сперматогенез. Действие активина противоположно и менее выражено. • Разные изоформы интерлейкинов , которые также синтезируются в сустеноцитах под влиянием фоллитропина, оказывают на сперматогенез разнохарактерное действие. Будучи по своей природе цитокинами, интерлейкины влияют только на фазу размножения сперматогоний: . интерлейкин 1 стимулирует ее, интерлейкин 6 тормозит.

• Важным фактором регуляции фазы размножения сперматогенеза служит белок трансферрин. Это естественный митоген, стимулирующий пролиферацию сперматогоний и, вероятно, мейоз и сперматогенез, поскольку специфические рецепторы к трансферрину , помимо сперматогоний, обнаружены у сперматоцитов 1 порядка и сперматид. • Андрогенсвязывающий белок поддерживает высокую концентрацию мужского полового гормона тестостерона, который выделяют под влиянием лютропина клетки Лейдига. Связывает тестостерон и переводит его в полость канальца, создавая высокую концентрацию тестостерона, необходимую для фазы формирования.
. Под воздействием ЛГ")
После наступления половой зрелости гипофиз мужчины начинает выделять лютеинизирующий гормон (ЛГ). Под воздействием ЛГ клетки Лейдига выделяют большие количества мужского полового гормона тестостерона. Последний, в свою очередь, инициирует сперматогенез, возможно благодаря действию на клетки Сертоли. Тестостерон оказывает прямой эффект на сперматиды, стимулируя их к трансформации в сперматозоиды. Важнейший эффект тестостерона заключается также в репрессии генов апоптоза в половых клетках, что крайне важно для формирования полноценного количества зрелых спермиев. Недавно открыто влияние тестостерона на миоидные (гладкомышечные) клетки извитых семенных канальцев. • Кроме тестостерона, клетки Лейдига выделяют небольшое количество эстрогенов. Помимо половых гормонов они вырабатывают эндорфин, небольшие количества окситоцина, стимулирующего сокращения перитубулярных миоидных клеток извитых семенных канальцев, а также интерлейкин 1, действующий как фактор роста на сперматогонии типа В. •

Спасибо за внимание

СТРОЕНИЕ И РАЗВИТИЕ ЖЕНСКИХ ПОЛОВЫХ КЛЕТОК лекция 2

• Наиболее очевидная отличительная черта яйцеклетки – это ее большие размеры. Типичная яйцеклетка имеет сферическую или овальную форму, а диаметр ее составляет у человека и морского ежа около 100 мкм, у лягушек и рыб от 1 до 2 мм, а у птиц и рептилий измеряется сантиметрами (величина типичной соматической клетки всего лишь около 20 мкм). Столь же внушительными могут быть размеры ядра яйцеклетки.

Отличительные особенности яйцеклетки • Все материалы, необходимые для начальных стадий роста и развития зародыша должны быть запасены в зрелом яйце. • Если спермий избавляется от большей части своей цитоплазмы, то развивающееся яйцо не только сохраняет содержащиеся в нем запасные материалы, но и активно увеличивает их количество. Либо синтезирует само, либо поглощает белки желтка, который служит источником питания развивающегося зародыша.

• Объем яйца как правило превосходит объем спермия во много раз ( у морского ежа в 10. 000 р). Ядерные гаплоидные компоненты примерно у них равны, а вот по содержанию цитоплазматических компонентов различия велики. Цитоплазма яйца - огромная кладовая запасов белка, рибосом, т. РНК. Вскоре после оплодотворения в яйце резко увеличивается синтез белка, который осуществляется при участии рибосом и т. РНК, присутствующих в яйце.

• У большинства организмов информация для осуществления белкового синтеза происходящего в раннем развитии заключена уже в ооците (25 -50 тысяч различных типов и. РНК), однако до оплодотворения они не активны. • Морфогенетические факторы, это молекулы, которые направляют дифференцировку клеток для образования различных тканей и органов, Они рассеяны по всему яйцу и распределяются между разными клетками при дроблении. Внутри этого огромного объема цитоплазмы яйца находится большое ядро.

Особенности строения яйцеклеток: В зрелом яйце сосредоточены ВСЕ материалы, необходимые для начальных стадий роста и развития зародыша. • Объем яйца превосходит объем спермия во много раз (морской ёж – 10 000 : 1). Различия по содержанию цитоплазматических компонентов. • Цитоплазма содержит: белки, рибосомы, т- и и-РНК, морфогенетические факторы (рассеяны по всему яйцу, распределяются между разными клетками при дроблении). Накоплены в период вителлогенеза. • Кортикальный слой цитоплазмы: высокая концентрация глобулярного актина, кортикальные гранулы многочисленные (до 15 000) гомологи акросомного пузырька спермия. Содержат протеолитические ферменты, мукополисахариды, гиалин.

Оболочки яйцеклетки: Располагаются поверх плазмалеммы. • первичная: производная плазмалеммы яйца. Иногда называется желточной. Присуща всем животным (кроме губок и стрекающих). Образована гликопротеинами. У человека называется прозрачной оболочкой (zona pellucida), образуя ее внутреннюю часть (внешняя часть продуцируется фолликулярными клетками). Блестящая оболочка яйцеклетки клеток. У насекомых называются хорионом. . В хорионе млекопитающих имеется микропиле (для проникновения спермия в яйцо). • вторичная: продукт деятельности фолликулярных • третичная: яйцо окружается ими во время прохождения по яйцеводу. Развиты у хрящевых рыб, амфибий, позвоночных (пресмыкающиеся, птицы, низшие млекопитающие). Образуются из секретов желез яйцевода. Функция: защита зародыша, функции запаса воды, питательных веществ. • К третичным относится также – студенистая оболочка (прикрепление к предметам, привлечение спермиев). Строение яйца курицы


Схема оогенеза: • размножение: оогонии • рост: ооцит I порядка • созревание: ооцит II порядка, полярные тельца, яйцеклетка. Яйцевые клетки образуются в яичнике. ППК мигрируют из места возникновения в развивающиеся гонады. А – стадия размножения, Б – стадия роста, В – стадия созревания

Стадии оогенеза: • стадия размножения: в гонадах ППК пролиферируют, делятся митозом и называются оогониями. У большинства низших позвоночных они сохраняют способность к делению на протяжении всего репродуктив- ного периода(наружное оплодот- ворение). • У млекопитающих размножение оогониев протекает только в эмбриональном периоде и к концу внутриутробного развития прекращается (у человека максимальное количество оогониев (6– 7 млн) наблюдается у 5 -ти месячного плода, у новорожденной девочки их около 1 млн, а к 7 -ми годам сокращается до 300 тысяч). А – стадия размножения, Б – стадия роста, В – стадия созревания

Стадии оогенеза: • стадия роста: женская половая клетка, прекратившая размножение, называется ооцит I порядка. Связан с поступлением в яйцеклетку питательных веществ извне и с синтезом их в самой яйцеклетке. Масса яй-ки увеличивается в 40 раз у мл-х различают 2 периода: – малый (цитоплазматический) рост или превителлогенез: пропорциональное увеличение массы ядра и цитоплазмы; ядерно-цитоплазматическое отношение не изменяется) – большой (трофоплазматический) рост или вителлогенез: интенсифицируется рост цитоплазматических компонентов, откладывается желток. А – стадия размножения, Б – стадия роста, В – стадия созревания
: пропорциональное увеличение")
Периоды роста яйцеклетки 3 • превителлогенез (малый или цитоплазматический 4 рост): пропорциональное увеличение массы ядра и цитоплазмы при подготовке ооцита I порядка к мейозу. На стадии диакинеза мейоз замедляется, вплоть до 2 половозрелости, 1 -й блок мейоза. • В ядре синтезируются все виды РНК. Синтез р. РНК связан с процессом амплификации генов(временное 5 увеличение числа генов, кодирующих данные виды РНК). Отделившиеся копии обособляются в виде ядрышек, 6 1 после созревания ооцита они выходят в цитоплазму и лизируются. • Синтез низкомолекулярных р. РНК и т. РНК идёт без Ооциты лягушки (Маслова, Сидоров, 2008): амплификации, т. к. кодирующие их гены многократно 1 – клетки на ранней повторены. стадии вителлогенеза; 2 – клетка на поздней • Усиливающаяся синтетическая активность стадии вителлогенеза; нуклеиновых кислот приводит к образованию хромосом 3 – ядро с ядрышками; типа ламповых щеток, что связано с наличием 4 – фолликулярные клетки; деспирализованных участков ДНК, на которых идет 5 –цитоплазма с синтез и. РНК В зрелом яйце насчитывается до 25 -50 гранулами желтка; тыс. типов и. РНК. Боль-во их находится в неактивном 6 – кортикальный слой состоянии в цитоплазме и активируется после оплодотворения.
: • • • интенсифицируется рост цитоплазматических компонентов, образуется и")
вителлогенез (большой или трофоплазматический рост): • • • интенсифицируется рост цитоплазматических компонентов, образуется и откладывается желток, жиры, гликоген. Количество желтка детерминировано генетически и не зависит от условий питания. Желток откладывается в вегетативном, а ядро ооцита оттесняется в анимальное полушарие: эндогенный желток: синтезируется в эндоплазматическом ретикулуме из цистерн аппарата Гольджи, накапливается в митохондриях, перерождающихся в желточные гранулы; Ооциты лягушки (Маслова, Сидоров, 2008): 1 – клетки на ранней стадии вителлогенеза; 2 – клетка на поздней стадии вителлогенеза; 3 – ядро с ядрышками; 4 – фолликулярные клетки; 5 –цитоплазма с гранулами желтка; 6 – кортикальный слой 3 4 2 5 6 1

экзогенный желток: синтез экзогенного желтка вителлогенина осуществляется клетками печени под контролем эстрогена, который выделяют в кровоток, под влиянием гормонов гипоталамуса и гипофиза, фолликулярные клетки. Вителлогенин позвоночных из печени матери транспортируется к фолликулу по кровеносным сосудам и поглощается ооцитом путем пиноцитоза, при формировании гранул желтка распадается на фосфовитин и липовителлин.

Стадия созревания: Деления созревания резко неравномерны. В результате первого деления половина хромосомного набора выталкивается в редукционное (полярное или направительное) тельце (ни само, ни его потомки никакого участия в дальнейшем развитии не принимают). Яйцеклетка после выделения первого редукционного тельца называется ооцитом II порядка. При втором делении созревания выделяется второе редукционное тельце таких же размеров, как и первое. После его выделения ооцит II порядка превращается в истинную яйцеклетку) Биологический смысл неравномерности делений- невыгодно дробить накопленный запас питательных веществ яй-ки

• При подготовке к 1 делению созревания ооцит долго находится в фазе диакинеза, когда и происходит его рост и вителлогенез. • Выход из фазы диакинеза и начало собственно делений созревания приурочены к достижению самкой половозрелости. • Наличие блока (на стадии диакинеза профазы 1 мейоза) обусловлено накоплением необходимых компонентов для последующего развития и связано с отсутствием функционально активного фактора стимулирующего созревание (MPF), отвечающего за возобновление мейоза.

• Это белковый комплекс, состоит из 2 -х субъединиц: малой –протеинкиназный компонент Сdk 1( для позвоночных) и большой - регуляторный компонент циклин В. • В виде активного комплекса циклинзависимая киназа и циклин осуществляет фосфорилирование определенных белков, активируя или ингибируя их активность в соответствующую фазу клеточного цикла. • Активность комплекса начинает расти в S фазе цикла, достигает максимума в начале митоза, а затем, в анафазе резко падает из-за деградации циклина, который ведет к инактивации Сdkкиназы.

Механизм снятия 1 блока мейоза • Лучше всего изучен у амфибий. При достижении половой зрелости гонадотропины вызывают выделение фолликулярными клетками прогестерона, связываясь с рецепторными белками плазмалеммы, он инактивирует аденилатциклазу (падение уровня ц. АМФ в цитозоле и снижение активности Акиназы, отвечающей за фосфорилирование белков), что приводит к дефосфорилированию MPF и переходу его в активное состояние (нарастает его количество т. к. способен к автокаталитическому саморазмножению).

• Под действием MPF ооцит 1 порядка приступает к делениям созревания (запускает переход из профазы I деления мейоза в метафазу II). • Зрелые ооциты задерживаются на стадии метафазы II, когда уровень MPF высок. • Для завершения клеточного деления необходимо разрушить циклин В (разрушить MPF). Он разрушается в результате протеолиза комплексом убиквитинлигаза (anaphas promoting complex(APC/С), который присоединяет к циклину небольшой белок убиквитин.
 • Однако в созревающих ооцитах, параллельно с нарастанием активности MPF, при")
Стадия созревания (механизм) • Однако в созревающих ооцитах, параллельно с нарастанием активности MPF, при подготовке к 1 делению созревания, в цитоплазме под, действием прогестерона, накапливается цитостатический фактор (СSF), продукт с-mos гена, способный ингибировать APC/С. • В результате циклин В не разрушается, уровень MPF в цитоплазме остается высоким, наблюдается второй блок мейоза, хромосомы остаются в составе метафазной пластинки веретена второго деления.
 Разрушение MPF происходит при оплодотворении и связано с деградацией CSF,")
Стадия созревания (механизм) Разрушение MPF происходит при оплодотворении и связано с деградацией CSF, которую осуществляет Сазависимая протеинкиназа кальпен П, активирующаяся ионами Са 2+, поступающими из эндоплазматического ретикулума в результате активации яйцеклетки при оплодотворении. В итоге восстанавливается активность APC/С, циклин В разрушается, cdk инактивируется и клетка завершает деления созревания, затем процесс синтеза циклина, образование активного MPF и его разрушение в анафазе митоза повторяется в многочисленных делениях дробления зародыша. .
.")
Блок мейоза У большинства животных течение мейоза останавливается на некотором этапе созревания (блок мейоза). Для дальнейшего развития требуется оплодотворение. Типы блока мейоза (на этом этапе происходит овуляция): • на стадии диакинеза (губки, моллюски, отдельные представители плоских, круглых, кольчатых червей, млекопитающие: собака, лиса, лошадь); • на стадии метафазы 1 -го деления созревания (губки, немертины, кольчатые черви, насекомые); • на стадии метафазы 2 -го деления созревания (хордовые; у летучих мышей блок мейоза наступает в анафазе 2 -го деления созревания). Цитостатический фактор (CSF, продукт сmos гена) блокирует развития мейоза, предотвращая деградацию циклина и сохраняя активность MPF. Комплекс APC/C (циклосома) опосредует деградацию циклина-В в протеосомах (убиквитинизация белка).

Отличия оогенеза от сперматогенеза • . Период роста при оогенезе длиннее, чем при сперматогенезе и состоит из двух периодов – превителлогенеза и вителлогенеза. • 2. В ходе оогенеза стадия формирования отсутствует • 3. Из ооцита 1 порядка в противоположность сперматоциту 1 порядка получается не 4, а лишь одна полноценная половая клетка 4 Образование яйцеклетки идет периодически, сперматогенез идет постоянно. • • 1 • 5. Стадия размножения при оогенезе заканчивается в пренатальном периоде или сразу же после рождения, при сперматогенезе идет постоянно. • 6. Деления созревания оогенеза неравномерны и приводят к выделению полярных телец не участвующих в дальнейшем развитии. • 7. Сперматогенез более подвержен влиянию факторов внешней среды, чем оогенез, что обусловлено расположением половых органов • Зрелый семенник непрерывно вырабатывает огромное количество сперматозоидов, а половозрелый яичник женщины один раз в лунный месяц выделяет зрелую яйцеклетку, созревающую из ооцитов, заложенных на ранних стадиях эмбриогенеза. Чем старше возраст женщины, тем больше вероятность повреждения ее генетического материала мутагенными факторами. У мужчин сперматогонии постоянно интенсивно обновляются митозом, что приводит к обновлению генетического материала, а в некоторых случаях и к его репарации, поэтому возраст мужчин существенно не влияет на частоту наследственных болезней потомства.

Поляризация яйцеклетки Полюс яйцеклетки, на котором выделяются редукционные тельца, называется анимальным, а противоположный ему – вегетативным. Анимально-вегетативная ось ориентирует последующие морфогенетические процессы. Морфологические проявления поляризации приурочены к вителлогенезу. Желток откладывается преимущественно в вегетативном полушарии. Ядро ооцита оттесняется в анимальное полушарие (где больше свободной цитоплазмы). Поляризация становится устойчивой и необратимой в период выделения второго редукционного тельца. Расположение ионных каналов и насосов определяет полярность. На анимальном полюсе больше ионных каналов, на вегетативном –насосов Материальные носители полярности локализованы в плазмат. мембране Ооцит II порядка после овуляции (по Алмазову, Сутулову, 1978): 1 – первое редукционное тельце; 2 – веретено второго деления созревания; 3 – блестящая зона; 4 – фолликулярные клетки

Строение яичника млекопитающего Яичник – парный орган, покрытый с поверхности брюшиной. Собственно яичник покрыт специальным типом эпителия – герминативным, или зародышевым. Имеет корковое и мозговое вещество. Развитие фолликул в корковом слое. Общая продолжительность развития примордиального фолликула до стадии преовуляторного фолликула у человека составляет около 120 суток. • примордиальные фолликулы: по периферии. Это ооцит I порядка (диплотена профазы I), покрытый слоем плоских фолликулярных клеток (из коркового слоя яичника), внедряющиеся в соединительнотканную строму яичника. Порциями вступают при половом созревании в стадию роста и развития в течение всего репродуктивного п-да • первичные фолликулы: растущий ооцит с образующейся на этой стадии блестящей оболочкой и 1 -2 слоями кубических фолликулярных клеток. Начинается формирование вторичной оболочки окружающей фолликул( теки). Пока растет сам ооцит 1 пор. фолликул остается компактным :
 сохраняется вплоть до имплантации. Проницаема")
Функции оболочек • Прозрачная оболочка неклеточное образование (гликопротеины, гликозаминогликаны) сохраняется вплоть до имплантации. Проницаема для микромолекул и барьер для макромолекулярных соединений, обеспечивает защиту как самой яйцеклетки, так и зародыша, особенно важно это когда организм матери сенсибилизирован к отцовским аллоантигенам. На ней присутствуют рецепторы для присоединения сперматозоидов. • Главная функция фолликулярного эпителия – избирательный транспорт к ооциту необходимых питательных веществ, синтезируемых в печени или других органах материнского организма. Фолликулярные клетки выделяют белки, идущие на построение вторичной оболочки яйцеклетки(тека). Фолликулярные клетки играют роль не только опорных, трофических элементов, но вместе с соединительнотканными клетками теки являются железой внутренней секреции, вырабатывающей эстроген. Кроме того , на них расположены рецепторы к гонадотропным гормонам гипофиза, под воздействием которых осуществляется развитие фолликулов и оогенез. •

Строение яичника млекопитающего • вторичные фолликулы: в результате размножения фолликулярных клеток первичные фолликулы становятся многослойными. Фолликулярные клетки выделяют жидкость и постепенно резорбируются с образованием полостей. Внешняя соединительно-тканная оболочка фолликула (тека) дифференцируется на два слоя (внутренний и наружный). В наружную оболочку врастают многочисленные капилляры Ооцит во вторичном фолликуле находится на стадии диплотены профазы I мейоза. Полости затем сливаются в одну.

Строение яичника млекопитающего • третичный фолликул: или Граафов пузырёк. • Снаружи покрыт базальной мембраной и текальной оболочкой, содержащей сосуды во внутреннем слое и мышечные клетки в наружном. Фолликулярные клетки стенки фолликула (зернистая оболочка) секретируют жидкость, содержащую эстроген. Ооцит находится на яйценосном бугорке, окружённый фолликулярными клетками лучистого венца (corona radiata). В ооците реиннициируется мейоз – возникает ооцит II порядка и первое редукционное тельце. Приступает к 2 -му делению созревания и блок на стадии метафазы II

Строение яичника млекопитающего • овуляция: у большинства млекопитающих происходит на стадии метафазы II. Завершение второго деления созревания (образование яйцеклетки) и выделение второго редукционного тельца происходит лишь в процессе оплодотворения. Выпячивает поверхность яичника (стигма). Разрыв стенки Граафова пузырька приводит к освобождению ооцита в брюшную полость (овуляция). • атретические тела: большинство ооцитов в фолликулах в разные периоды своего роста претерпевают обратное развитие (атрезию). Для таких фолликулов характерны прогрессирующее уплотнение яйцеклетки, утолщение прозрачной зоны, регрессия фолликулярного эпителия, лизис органелл, кортикальных гранул, сморщивание ядра. Блестящая зона становится складчатой, утолщается и гиалинизируется. Атрезия фолликул ранней стадии – дегенерационная(деструкция и гибель ооцита). Поздней – атретические тела продуцируют андрогены и небольшое количество эстрогенов. Атрезию следует рассматривать как механизм, направленный на предотвращение образования избыточного количества яйцеклеток.
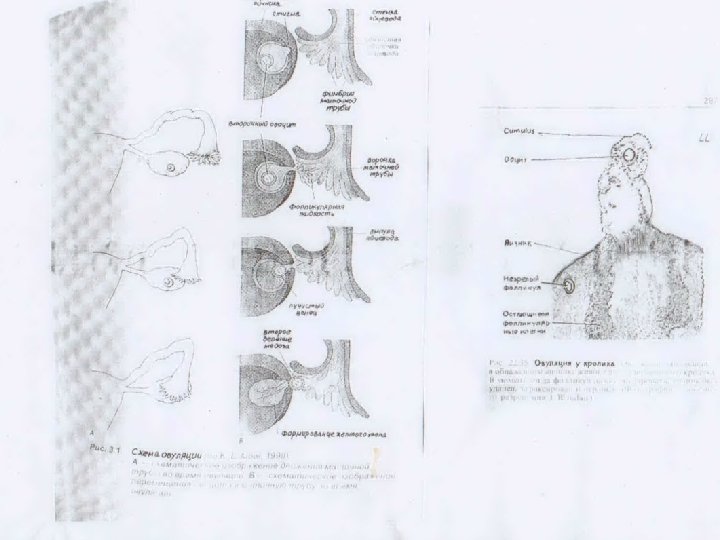
 Под влиянием")
Жёлтое тело яичника: (граафов пузырек после овуляции яйцеклетки преобразуется в желтое тело) Под влиянием лютеинизирующего гормона, фолликулярные клетки стенки лопнувшего Граафова пузырька претерпевают изменения, приводящие к формированию желтого тела – гипертрофируются и накапливают лютеин (лютеиновые клетки). Стенка фолликула частично спадается, фолликулярный эпителий и внутренняя тека собираются в складки, исчезают фрагменты базальной мембраны (лепестки цветка).

Различают два вида желтых тел овариальное жёлтое тело: существует в течение второго периода менструального цикла (с 14– 16 по 26– 28 день). Размер 1, 5– 2 см. В различные периоды овариального цикла оно обладает гормональной активностью – выделяет прогестерон (гормональная активность снижается к концу цикла). Претерпевает инволюцию. • жёлтое тело беременности: формируется при наступлении беременности. Размер – более 5 см, существует дольше, более активно в функциональном плане. Начиная с 3– 4 месяцев беременности функцию образования прогестерона берет на себя плацента. В последующем подвергается обратному развитию. Низкий уровень прогестерона может привести к выкидышу.

• В ходе инволюции жёлтого тела его железистые клетки атрофируются, а соединительная ткань центрального рубца разрастается. В результате на его месте формируется белое тело. Оно сохраняется в яичнике на протяжении нескольких лет, а затем практически полностью рассасывается
")
Развитие фолликулов в яичнике млекопитающего (схема)
")
Развитие фолликул в яичнике млекопитающего(схема)
 рост")
Регуляция полового цикла млекопитающих Рост фолликулов начинается еще в яичниках зародыша. Первичный (малый) рост не зависит от гормонов гипофиза и приводит к возникновению фолликулов с небольшой полостью. Для дальнейшего роста (большого) необходимо стимулирующее влияние гормонов аденогипофиза. Гонадолиберин, секретируемый нейронами гипоталамуса, запускает выработку клетками аденогипофиза следующих гормонов, попадающих затем в кровеносное русло: • фолликулостимулирующего гормона (ФСГ): повышение уровня ФСГ стимулирует рост фолликула и способствует развитию первичных фолликул во вторичные, а затем одного из вторичных фолликулов в Граафов пузырёк; • лютеинизирующего гормона (ЛГ): усиливает в яичниках секрецию прогестинов. Созревающие фолликулы синтезируют эстрогены (17 эстрадиол). Выброс ЛГ инициирует овуляцию, а оставшиеся после неё в составе яичника фолликулярные клетки начинают продуцировать прогестерон уже в качестве клеток жёлтого тела. Схема регуляции овариальноменструального цикла человека (по Вутке, 1996): ГЛ – гонадолиберин, ЛГ – лютеинизирующий гормон, ФСГ – фолликуло-стимулирующий гормон
 фаза: растущие фолликулы секретируют")
Фазы овуляторного цикла человека • фолликулиновая (эстрагенная или пролиферативная) фаза: растущие фолликулы секретируют всё больше эстрогенов, вызывая пролиферацию эндометрия матки и увеличивая чувствительность гипофиза к гонадолиберину, что повышает уровень ФСГ и ЛГ, выброс которых инициирует овуляцию (14 день цикла). • лютеиновая (гестагенная или секреторная) фаза: желтое тело (corpus luteum), фаза расцвета которого усиливается и удлиняется благодаря влиянию пролактина гипофиза, секретирует прогестерон. Эндометрий матки превращается в секреторный, подготавливая её для имплантации зародыша, ингибируется рост новых фолликулов. В сочетании с эстрогеном, уровень которого всё ещё высок, Овариально-менструальный цикл человека (по Гилберт, 1994): прогестерон подавляет активность гипофиза Шкала времени указывает на дни менструального и гипоталамуса, возвращая исходный цикла. М – менструация, ЛФ – лютеиновая фаза, ФФ – фолликулиновая фаза. уровень ФСГ и ЛГ.

Способы питания яйцеклеток • • • Лишь немногие яйцеклетки развиваются исключительно за счет эндогенного желтка. В зависимости от способа доставки к клетке веществ, необходимых для синтеза желтка различают: диффузный (фагоцитарный): наиболее примитивный способ. Встречается у губок и пресноводной гидры. Настоящих желточных гранул не образуется. Растущий ооцит питается, перемещаясь по межклеточному пространству (фагоцитированный материал откладывается в фаголизосомах). солитарный (одиночный): растущий ооцит получает все необходимые для синтеза желтка вещества из окружающей среды в низкомолекулярной форме. Присущ колониальным гидроидным полипам, морским звездам, ланцетнику. При этом желток и все типы РНК синтезируются самими ооцитами. нутриментарный: ооцит окружен трофоцитами, связанными с ним цитоплазматическими мостиками (различные группы червей и членистоногие). Функция трофоцитов – синтез р. РНК, поступающей в виде комплекса с рибосомными белками в яйцеклетку. Основная часть желточных белков синтезируется в соматических клетках и поступает в ооцит посредством пиноцитоза. фолликулярный: наиболее распространенный и совершенный способ. Связан с образованием слоев фолликулярного эпителия, окружающего ооцит.

Регуляция полового цикла млекопитающих Половозрелый яичник только раз в 28 дней выделяет зрелую яйцеклетку, созревающую из ооцитов, заложенных на ранних стадиях эмбриогенеза. Рост фолликулов начинается еще в яичниках зародыша. Малый рост не зависит от гормонов гипофиза, а для большого роста необходим фолликулостимулирующий гормон (ФСГ) и лютеинизирующий гормон (ЛГ). Их продукция контролируется гонадолиберином гипоталамуса.

С п а с и б о внимание за

ОПЛОДОТВОРЕНИЕ лекция 3

• Интерес к проблеме оплодотворения выходит далеко за рамки собственно эмбриологии. Слияние гамет – плодотворно используемая модель для изучения тонких молекулярных и клеточных механизмов специфического взаимодействия клеточных мембран; для изучения молекулярных основ активации метаболизма и пролиферации соматических клеток. Общебиологический интерес представляет и то, что оплодотворение являет собой яркий уникальный пример полного обращения клеточной дифференциации.

• . • Высокоспециализированные половые клетки не способны к самовоспроизведению. Однако после слияния они превращаются в тотипотентную клетку, которая служит источником формирования всех клеточных типов, присущих данному организму. • Сущность этого процесса была определена Оскаром Гертвигом при изучении морских ежей в конце 1870 годов и представляет собой слияние ядер половых клеток.

• После своего выхода из гонады как яйцеклетка, так и спермий обречены на гибель в считанные часы, если они не отыщут друга и не сольются в процессе оплодотворения. Оплодотворение спасает их от гибели, яйцеклетка активируется и приступает к осуществлению программы развития, а ядра двух гамет сливаются, формируя геном нового организма.

• Большая часть того, что нам известно об оплодотворении –это результат исследований на морских беспозвоночных, в особенности на морских ежах. В связи с этим наше обсуждение оплодотворения будет относиться, главным образом, к морским ежам и, в сравнительном аспекте, применительно к млекопитающим. Несмотря на огромную эволюционную дистанцию, которая отделяет млекопитающих от морских ежей, клеточные и молекулярные механизмы, лежащие в основе процесса оплодотворения, весьма сходны. Готовность яйцеклеток к оплодотворению определяется выделением направительных телец. У одних животных( морской еж) сперматозоиды проникают в яйцеклетку после выделения направительных телец, у лошадиной аскариды до их выделения, у млекопитающих и человека в момент выделения.

Особенности процесса оплодотворения: Оплодотворение – слияние сперматозоида с яйцеклеткой, завершающееся объединением их ядер в единое ядро оплодотворенного яйца (зиготы). Функции: половая (комбинирование генов двух родителей) – передача генов от родителей потомкам; репродуктивная (создание нового организма) – включает инициацию в цитоплазме яйца тех реакций, которые позволяют продолжать развитие. Роль спермия: активация яйца – побуждение яйцеклетки к началу развития; внесение в яйцеклетку генетического материала мужской особи Классификация: по месту проникновения сперматозоида в яйцеклетку: по количеству сперматозоидов, участвующих в оплодотворении: наружное – во внешней среде; моноспермное – один спермий внутреннее – в половых путях полиспермное – два и более самки.

У беспозвоночных животных, рыб, хвостатых амфибий и птиц возможна полиспермия, когда в яйцеклетку проникает несколько сперматозоидов, однако сливается с ядром яйцеклетки ядро одного спермия. Любому оплодотворению предшествует процесс эякуляции – излияние жидкости в женские половые пути или в водную среду. Продолжительность жизни спермия и яйцеклетки как при наружном, так и внутреннем оплодотворении невелика. Яйцеклетки многих беспозвоночных, а также рыб и амфибий должны быть оплодотворены сразу же после овуляции.

К моменту встречи с яйцеклеткой спермий должен сохранить не только активное движение, но и свою оплодотворяющую способность. Она зависит от концентрации спермы, р. Н среды, температуры. В щелочной среде спермии более активны, но быстро растрачивая энергию, раньше погибают. В подкисленной среде их активность меньше, а продолжительность жизни больше. Концентрация сперматозоидов в 1 см 3 эякулята достигает 100 000, при снижении ее наполовину – достаточно высокий риск бесплодия, при концентрации 20 000 и ниже мужчины бесплодны.

Оплодотворение - это не одномоментный процесс, а многоступенчатый, в котором выделяют следующие стадии: дистантные взаимодействия – осуществляются на некотором расстоянии, до соприкосновения гамет друг с другом; контактные взаимодействия – начинаются с момента контакта сперматозоида с наружной оболочкой яйцеклетки; проникновение спермия в яйцеклетку – в основе процесса лежит слияние плазматических мембран спермия и яйца; слияние генетического материала – (пронуклеусов) приводит к формированию диплоидного ядра зиготы.

Дистантные взаимодействия Представление о гамонах веществах обеспечивающих активацию или блокирование отдельных этапов процесса оплодотворения • • Первая стадия, дистантное взаимодействие по названию не предполагает контакт между гаметами Уже в ходе ранних исследований процесса оплодотворения возникло представление о гамонах – веществах, обеспечивающих активацию или блокирование отдельных его этапов. Яйцеклетка способна продуцировать гиногамоны, а сперматозоид андрогамоны. Полагали, что гиногамон 1 - это низкомолекулярное вещество небелковой природы, которое активирует движение сперматозоидов, повышая тем самым вероятность их встречи с яйцом. Андрогомон 1 подавляет подвижность спермиев Гиногамон II, часто именуемый фертилизином –гликопротеин, расположеный в периферической области яйца, вызывает склеивание его со сперматозоидами при взаимодействии с комплементарным ему андрогамоном II (антифертилизин), встроенным в поверхностную оболочку спермия. Андрогамон II способен разжижать студенистое вещество и растворять оболочку яйца, поэтому его отождествляют с гиалуронидазой. Гипотезы оплодотворения, основанные на идее о взаимодействии фертилизина с комплементарным ему антифертилизином, сыграли свою положительную роль, поскольку позволили обнаружить существование целого семейства специфических молекул на поверхности взаимодействующих гамет.

Дистантные взаимодействия гамет: - направлены на повышение вероятности встречи сперматозоидов и яйцеклетки -характерны для водных организмов, с наружным типом оплодотворения. - встречу спермиев и яиц при их низкой концентрации в среде; - предотвращение оплодотворения яиц спермиями другого вида. - видоспецифичные привлечения спермиев и их активацию (кишечнополостные, моллюски, иглокожие, первичнохордовые и т. п. ) Хемотаксис – движение по градиенту концентрации какого-либо вещества. У морских ежей (пептиды студенистой оболочки яиц): • сперакт (10 а. к. ) Взаимодействуют с рецепторами плазматической мембраны спермия (кислые гликопротеины), активируют метаболизм, увеличивают подвижность муж. гамет • резакт (14 а. к. ) Mov 1

Контактное взаимодействие гамет Первый этап – акросомная реакция. У морского ежа акросомную реакцию инициируют сульфатированные полисахариды студенистой оболочки. Восприятие этих сигналов обеспечивается соответствующими рецепторами мембраны спермия, что приводит к открытию неселективных катионных каналов и поступлению внешнего кальция в головку спермия. Концентрация кальция в цитозоле клетки увеличивается. Как известно повышение концентрация Са 2+ служит предпосылкой различного рода секреторных процессов.

Акросомная реакция У сперматозоида повышение уровня Са 2+ сверх определенного порога служит сигналом к началу экзоцитоза содержимого акросомы. Мембрана акросомы сливается с мембраной спермия, содержимое пузырька секретируется во внешнюю среду и различные гидролитические ферменты начинают разрушать студенистую и желточную оболочку яйца. Увеличение концентрации ионов кальция вызывает активацию Na+/H+ -обменника, происходит выведение ионов водорода в ответ на повышенный приток ионов натрия, что приводит к внутриклеточному сдвигу р. Н в щелочную сторону.

Сдвиг р. Н способствует полиме ризации глобулярного актина и образованию акросомного выроста, активации динеиновой АТФ-азы в шейке спермия, что увеличивает его подвижность. Повышение концентрации осмотически активных ионов (Са 2+, Na+, СI-) вызывает приток воды и резкое увеличение гидростатического давления, что обеспечивает удлинение акросомального выроста.

Контактные взаимодействия гамет: акросомная реакция • Инициируют сульфатированные полисахариды студенистой оболочки • поступление Ca 2+ в головку спермия • слияние мембран головки спермия и акросомы • экзоцитоз акросомного пузырька (выброс протеолитических ферментов) • активация Na+/H+-обменника (защелачивание цитоплазмы) • полимеризация глобулярного актина (образование акросомного выроста) • активация динеиновой АТФ-азы (увеличение подвижности спермия) • повышение осмотического давления (способствует удлинению акросомального выроста) Акросомная реакция спермия морского ежа 1 – акросомная мембрана; 2 – плазматическая мембрана спермия; 3 – глобулярный актин; 4 – акросомные ферменты; 5 – актиновые микрофиламенты; 6 – акросомный вырост

Контактное взаимодействие гамет Второй этап контактного взаимодействия гамет связан с узнаванием спермия и яйца. Сразу же после того, как спермий морского ежа проник в студенистую оболочку яйца, его акросомный вырост вступает в контакт с наружной поверхностью желточной оболочки. Основной этап узнавания в этот момент. Белок акросомы при посредстве которого происходит это узнавание – Биндин. Рецептором биндина у морских ежей служит комплекс из четырех гликопротеиновых субъединиц, находящийся на желточной оболочке.

Контактные взаимодействия гамет: Узнавание спермия и яйцеклетки Морские ежи: Видоспецифичное узнавание при помощи белка (биндина), расположенного на поверхности акросомного выроста. На желточной оболочке яйца находится гликопротеиновый комплекс (рецептор), способный образовывать связи с биндином. Локализация биндина на акросомном выросте (по Гилберту, 1993) ДАБ – диаминобензидин.

Гликопротеиновый комплекс состоит из четырех субъединиц Каждая субъединица имеет короткую цитоплазматическую область, трансмембранную область и внеклеточный, взаимодействующий с биндином домен, часть которого имеет структурное сходство с белком теплового шока Нsp 70. Во внеклеточном домене выявлена зона с высоким содержанием цистеина. Как полагают, именно здесь образуются дисульфидные мостики, соединяющие субъединицы в гомотетрамер – функционально активный рецептор биндина, который видоспецифически связывает биндин. У каждого вида морских ежей вырабатывается свой особый тип биндина и молекулы его связываются с вителлиновым слоем. У морского ежа насчитывается до 6000 сайтов таких соединений.

• Таким образом узнавание гамет морского ежа происходит на двух уровнях: уровень активации акросомной реакции при контакте со студенистой оболочкой и уровень прикрепления спермия к желточной оболочке

Особенности оплодотворения у млекопитающих У млекопитающих оплодотворение внутреннее и процесс этот приспособлен к внутренней среде организма. Акросомной реакции предшествует заякоривание спермия на оболочке яйца. Акросомная нить у них не образуется. Половые пути самки принимают активное участие в процессе оплодотворения. Спермии млекопитающих сразу после эякуляции неспособны к акросомной реакции, так как некоторое время должны находиться в половых путях самки. Условия, требующиеся для капацитации (приобретение спермием оплодотворяющей способности) варьируют в зависимости от вида.
: Капацитация – приобретение спермием оплодотворяющей способности под воздействием секрета половых путей самки")
Капацитация (1): Капацитация – приобретение спермием оплодотворяющей способности под воздействием секрета половых путей самки Суть процесса капацитации: - изменение структуры клеточной мембраны: - снижение соотношения холестерин : фосфолипиды в мембране спермия (молекулы альбумина половых путей самки отнимают холестерин у спермия) – дестабилизация мембраны сперматозоида – возможность слияния мембран спермия и акросомы - удаление с поверхности спермия особых факторов, (coating factors), оставаясь на поверхности спермия, эти факторы препятствуют оплодотворению
: Поверхность спермия содержит гликозилтрансферазу (узнаёт концевые остатки Nацетилглюкозамина на прозрачной оболочке яйцеклетки)")
Капацитация (2): Поверхность спермия содержит гликозилтрансферазу (узнаёт концевые остатки Nацетилглюкозамина на прозрачной оболочке яйцеклетки) В спермиях, не прошедших капацитации, активные центры фермента блокированы остатками Nацетилглюкозамина (NАг) и галактозы (Гал). Схема узнавания спермием и яйцом друга у млекопитающих (схема капацитации) При капацитации углеводы отделяются от поверхности спермия, освобождая активные центры гликозилтрансфераз. Гликозилтрансферазы узнают N-ацетилглюкозаминовые остатки в молекуле гликопротеина, расположенного на поверхности прозрачной оболочки (рецептор спермия).

Проникновение спермия у млекопитающих: Блестящая оболочка яйцеклетки – образована гликопротеинами (ZP 1, ZP 2, ZP 3), которые соединяются в виде цепочек. Главным компонентом является ZP 3: обеспечивает первичное связывание спермия и индуцирует акросомную реакцию. Типы гликопротеинов (zona proteins, ZP): ZP 1 – сшивает белки ZP 2 и ZP 3, находясь перпендикулярно по отношению к ним; • ZP 2 – расположен параллельно поверхности яйца. Взаимодействует с акрозином (протеолитический фермент акросомы), лизирующим блестящую оболочку; • ZP 3 – расположен параллельно поверхности яйца. Необходим для взаимодействия с рецепторами спермия (терминальной галактозой, • N-ацетилглюкозамином, индуцирует акросомную реакцию

Проникновение спермия у млекопитающих: У млекопитающих активация сперматозоида не сопровождается образованием акросомного выроста. Акросомная реакция у млекопитающих: • сперматозоид контактирует с яйцом не вершиной, а боком; • диссоциация наружной мембраны головки сперматозоида и мембраны акросомы (на поверхности, расположенной вдоль головки сперматозоида); • ферменты акросомы растворяют клетки лучистого венца сперматозоид вступает в контакт с блестящей оболочкой с помощью МСР Membrane cofactor protein (MCP) экспрессируется на внутренней мембране акросомы.

Проникновение сперматозоида в яйцо: Реакция активации сперматозоида заканчивается слипанием задней мембраны акросомы спермия и мембраны яйцеклетки, их разрывом и соединением свободных концов. Формируется единая наружная мембрана, ограничивающая канал, через который ядро и проксимальная центриоль сперматозоида проникают в яйцо. У морского ежа контакт между сперматозоидом и яйцом вызывает полимеризацию актина и увеличение размеров микроворсинок яйца – формируется воспринимающий бугорок (гомологичен акросомному выросту). Актиновый цитоскелет ворсинок – дополнительный источник энергии для погружения спермия в яйцо. Mov 3 Проникновение спермия в яйцеклетку хомячка (по Yanagimachi, R. and Noda, Y. D. 1970. Electron microscope studies of sperm incorporation into the hamster egg. American Journal of Anatomy 138: 429 -462).

Блоки полиспермии. Быстрый блок полиспермии После контакта спермия с плазматической мембраной яйца она должна утрачивать способность связываться с другими спермиями. Быстрый блок полиспермии достигает этой цели путем изменения электрического потенциала плазматической мембраны яйца. В норме на мембране ооцита потенциал равен -70 м. В. Мембрана препятствует проникновению Na+ утечке К+ Через 0, 1 сек после прикрепления спермия мембрана яйца деполяризуется и мембранный потенциал становится положительным (+20 м. В).

Быстрый блок полиспермии Деполяризация является следствием резкого изменения Na+ проницаемости мембраны в результате чего положительно заряженные ионы натрия поступают внутрь клетки по градиенту концентрации, уничтожая отрицательный заряд внутренней стороны мембраны.

Открытие натриевых каналов в яйце индуцируется прикреплением к нему спермия. Проникновение спермиев через положительно заряженную мембрану невозможно!

Медленный блок полиспермии : Мембранный потенциал яйцеклетки остается положительным только около 1 мин, далее возвращается к норме. Поэтому нужен еще механизм, препятствующий полиспермии более длительное время. В большинстве яйцеклеток этот барьер создают вещества, которые освобождаются из кортикальных гранул в результате кортикальной реакции, запускаемой сильным повышением концентрации ионов Са 2+, высвобожденным из эндоплазматического ретикулума.
 обеспечивает содержимое кортикальных гранул, выделяемое при их экзоцитозе")
Медленный блок полиспермии (образование оболочки оплодотворения) обеспечивает содержимое кортикальных гранул, выделяемое при их экзоцитозе Повышение концентрации Са 2+ инициирует встраивание кортикальных гранул в мембрану яйца, далее они сливаются с ней и выделяют свое содержимое в область между мембраной яйцеклетки и желточной оболочкой, которая под действие выделяемых соединений претерпевает ряд изменений и называется оболочкой оплодотворения. Формирование оболочки оплодотворения (по Гилберту, 1993): 1 – оболочка оплодотворения; 2 – гиалиновый слой

содержимое кортикальных гранул, выделяемое при их экзоцитозе • вителлиновая деламиназа: отделяет желточную оболочку от цитоплазматической мембраны яйцеклетки; • сперморецепторная гидролаза: освобождает поверхность яйца от осевших на желточной оболочке сперматозоидов, лизируя сайты их соединения; мукополисахариды: создают осмотический градиент, обусловливающий поступление воды из цитоплазмы яйца в пространство между желточной оболочкой и плазматической мембраной (перивителлиновое пространство); • пероксидаза: вызывает затвердевание оболочки оплодотворения (образование поперечных связей между остатками тирозина соседствующих белков); • гиалин: поддерживает бластомеры в период дробления. • карбоангидраза отщепляет некоторые углеводные компоненты (задубливание) Формирование оболочки оплодотворения (по Гилберту, 1993): 1 – оболочка оплодотворения; 2 – гиалиновый слой
")
У млекопитающих кортикальная реакция вызывает преобразования прозрачной оболочки (реакция прозрачной оболочки или zona reaction)

• • • У млекопитающих, высвободившиеся из кортикальных гранул протеолитические ферменты, разрушают белки, связывающие плазмалемму с блестящей оболочкой. Происходит протеолиз или модификация ZP-рецепторов спермиев, что надежно блокирует полиспермию. Этот процесс изменения свойств рецепторов спермиев носит название реакции прозрачной оболочки (реакция зоны) Выделенные из кортикальных гранул гликозаминогликаны создают осмотический градиент, вызывающий поступление воды из цитоплазмы в область между мембраной и прозрачной оболочкой, образуется перивителлиновое пространство, прозрачная оболочка отделяется от плазмалеммы и называется оболочкой оплодотворения

Механизм кортикальной реакции: • прикрепление спермия к рецептору плазматической мембраны яйца • активация G-белка • стимуляция активности фосфолипазы С – расщепляет фосфатидилинозитол-4, 5 бифосфат (PIP 2) на диацилглицерол – DAG (связан с мембраной) и инозитолтрифосфат – IP 3 (диффундирует в цитоплазму, стимулирует выход Са 2+) • высвобождение Ca 2+ (из внутриклеточных депо) – посредством IP 3 , приводит к экзоцитозу кортикальных гранул (медленный блок полиспермии), активации протеинкиназы С, НАД-киназы (превращение НАД в НАДФ+ (синтез липидных компонентов мембраны de novo). • активация Na+/H+-антипортера (DAG, посредством протеинкиназы С) приводит к сдвигу р. Н в щелочную сторону, что вызывает усиление белкового синтеза, репликацию ДНК, перемещение морфогенетических детерминант в цитоплазме. • Mov 7
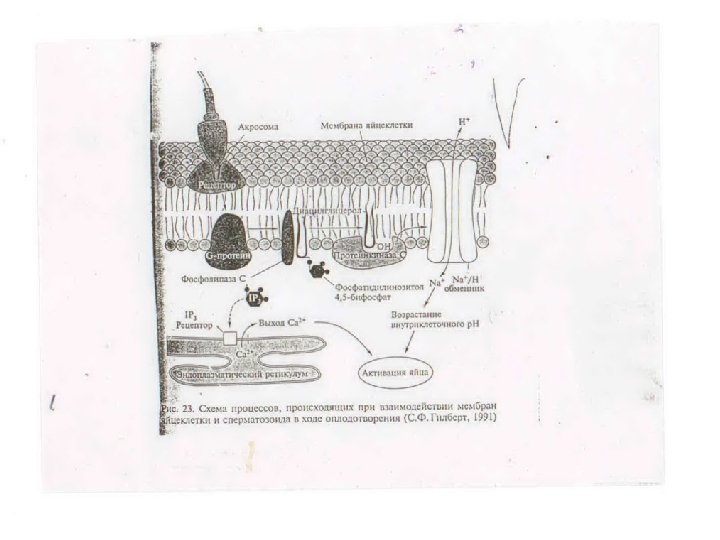

Механизм кортикальной реакции • • Прикрепление спермия к плазматической мембране яйца, через активацию находящегося в мембране G-белка, стимулирует активность мембраносвязанного фермента фосфолипазы С. Она расщепляет находящийся здесь же фосфатидилинозитол-4, 5 -бифосфат на диацилглицерол(ДАГ, (DAG) и диффундирующий в цитоплазму инозитолтрифосфат (ИТФ, IP 3). ИТФ вызывает открытие Са 2+ каналов эндоплазматического ретикулума, что приводит к высвобождению кальция из внутриклеточного депо. ДАГ посредством протеинкиназы С (С-киназа), активация которой возможна только в присутствии ионов Са 2+, активирует Na+/H+-антипортер, что приводит к увеличению внутриклеточного р. Н (натрий входит в клетку, а протон ее покидает). Сдвиг р. Н в щелочную сторону вызывает усиление белкового синтеза, репликацию ДНК, перемещение морфогенетических детерминант в цитоплазме при участии Са 2+. Высвобождение Са 2+ из внутриклеточных депо приводит к экзоцитозу кортикальных гранул и включению медленного блока полиспермии, а также образованию гиалинового слоя вокруг яйца. Другим Са 2+-зависимым эффектом является активация НАД-киназы, которая катализирует превращение НАД в НАДФ+, что в конечном итоге обеспечивает синтез новых липидных компонентов плазматической мембраны, необходимых для последующего протекания процессов дробления.

Слияние генетического материала • После попадания ядра спермия в цитоплазму яйцеклетки оно утрачивает свою оболочку. • Спермийные гистоны фосфорилируются и заменяются гистонами стадии дробления. Ядерная оболочка сперматического ядра распадается на мелкие пузырьки, что делает возможным воздействие цитоплазмы яйца на компактный хроматин спермия. • Одновременно с преобразованием структуры хроматина происходит сборка ламинов и возникает новая ядерная оболочка. Образовавшееся гаплоидное ядро называется мужским пронуклеусом.

• Ядро яйца после завершения второго деления созревания и приобретения пузыревидной формы называется женским пронуклеусом

Слияние генетического материала Митохондрии спермия и жгутик разрушаются и в цитоплазме развивающихся взрослых организмов не обнаруживаются (митохондрии передаются от материнского организма, а центриоли от отцовского, хотя в яйцеклетках млекопитающих они изначально присутствуют в яйцеклетке). После проникновения в цитоплазму мужской пронуклеус поворачивается на 180 о (центриоль спермия оказывается расположенной между мужским и женским пронуклеусом). Ее микротрубочки удлиняются, вступая в контакт с женским пронуклеусом, после чего пронуклеусы перемещаются навстречу другу. Mov 8
: У млекопитающих процесс сближения пронуклеусов продолжается около 12 часов (у")
Слияние генетического материала (2): У млекопитающих процесс сближения пронуклеусов продолжается около 12 часов (у морского ежа – всего 1 час). Репликация ДНК у млекопитающих происходит в пронуклеусах в период миграции. Контакт пронуклеусов приводит к разрушению их ядерных оболочек, конденсации хроматина и образованию видимых хромосом (располагаются на общем митотическом веретене первого деления дробления ). У млекопитающих истинно диплоидное ядро появляется у 2 -х клеточного эмбриона. Схема процесса оплодотворения у хомячка (по Гилберту, 1993): А – спермий прикрепляется к яйцу; Б – прикрепление спермия к яйцу и взмахи его хвоста вызывают вращение яйца; В – плазматические мембраны спермия и яйца сливаются; Г – формируются пронуклеусы; Д – начинается первое деление дробления; Е – двух клеточный зародыш

Преобразование цитоплазмы яйца • После проникновения сперматозоида начинаются интенсивные перемещения цитоплазмы яйцеклетки, усиление метаболических процессов в 70 -80 раз. Иногда при этом происходит расслоение (отмешивание) различных частей ооплазмы, что обозначается как ооплазматическая сегрегация. • В ходе этого процесса намечаются основные, хотя далеко не все, элементы пространственной организации зародыша. Поэтому данный этап называют проморфогенезом, так как расставляются вехи для будущих морфогенетических процессов. •

• Перемещения цитоплазмы приводят к ее биохимической неоднородности. Попадая в определенные участки зиготы, разные и. РНК и белки обеспечивают биохимическую неоднородность этих локусов. • При активации метаболизма отмечается всплеск синтеза новых белковых продуктов, которые сами могут становиться морфогенами и усиливают биохимическую неоднородность. В дальнейшем, попадая в разные бластомеры, эти детерминанты будут участвовать в активации и репрессии разных генов, а значит в приобретении клеткой определенных свойств, в ее дифференцировке.

Преобразование цитоплазмы яйца • У некоторых кишечнополостных сегрегация ограничивается процессом разделения ооплазмы на внешний ободок эктоплазмы (бедный питательным матриалом и иногда окрашенный различными пигментами) и внутренний - эндоплазму (богата желтком и другими питательными компонентами), что определяет в яйцах радиальную установку веретен делений дробления. Самыми наглядными примерами могут служить яйца асцидий и амфибий. • Ооплазматическая сегрегация влияет не на конечную дифференцировку клеток, а на ближайшие этапы развития – дробление и гаструляцию (проморфогенез).

Таким образом, перераспределение цитоплазматических масс произошло, пронуклеусы встретились, ДНК реплицируется, новые белки транслируются. Достигнута стадия, с которой может начинаться развитие многоклеточного организма. Это стадия дробления

• Спасибо за внимание!
d3ce7c91aa6e58bcb122a69298548792.ppt