

Lek_6_ugl.ppt
- Количество слайдов: 64
Триозы, образующиеся в процессе распада глюкозы,") ОСНОВНЫЕ УГЛЕВОДЫ И ИХ ХАРАКТЕРИСТИКА Физиологически важные моносахариды: 1)Триозы, образующиеся в процессе распада глюкозы, 2)Пентозы рибоза и дезоксирибоза, которые являются важными компонентами нуклеотидов, нуклеиновых кислот и коферментов. 3)Гексозы: глюкоза, галактоза, фруктоза и манноза.
ОСНОВНЫЕ УГЛЕВОДЫ И ИХ ХАРАКТЕРИСТИКА Физиологически важные моносахариды: 1)Триозы, образующиеся в процессе распада глюкозы, 2)Пентозы рибоза и дезоксирибоза, которые являются важными компонентами нуклеотидов, нуклеиновых кислот и коферментов. 3)Гексозы: глюкоза, галактоза, фруктоза и манноза.
 ДИСАХАРИДЫ: Состоят из 2 моносахаридных остатков, соединенных между собой гликозидной связью, которая образуется между двумя спиртами и является простым эфиром: R O R'. В первую очередь – это сахароза и лактоза.
ДИСАХАРИДЫ: Состоят из 2 моносахаридных остатков, соединенных между собой гликозидной связью, которая образуется между двумя спиртами и является простым эфиром: R O R'. В первую очередь – это сахароза и лактоза.
Гомополисахариды. Они состоят из моносахаров одного вида (глюкозы) •") • ПОЛИСАХАРИДЫ: Различают: • 1)Гомополисахариды. Они состоят из моносахаров одного вида (глюкозы) • Крахмал, декстрины, амилопектин, гликоген, клетчатка. • 2)Гетерополисахариды: Они состоят из цепей сложных углеводов, содержащих аминосахара и уроновые кислоты.
• ПОЛИСАХАРИДЫ: Различают: • 1)Гомополисахариды. Они состоят из моносахаров одного вида (глюкозы) • Крахмал, декстрины, амилопектин, гликоген, клетчатка. • 2)Гетерополисахариды: Они состоят из цепей сложных углеводов, содержащих аминосахара и уроновые кислоты.
 Крахмал представляет собой форму хранения питательных веществ в растениях. Это наиболее важный вид пищевых углеводов. Он содержится в злаках, картофеле, бобовых и др. Продукты гидролиза крахмала называют декстринами.
Крахмал представляет собой форму хранения питательных веществ в растениях. Это наиболее важный вид пищевых углеводов. Он содержится в злаках, картофеле, бобовых и др. Продукты гидролиза крахмала называют декстринами.
 Целлюлоза главный компонент структурной основы растений. Состоит из глюкозных остатков, соединенных, в отличие от крахмала, не α , а β(1 4) гликозидной связью. Многие млекопитающие, в том числе и человек не способны усваивать целлюлозу, т. к. они лишены фермента, расщепляющего такого рода гликозидную связь.
Целлюлоза главный компонент структурной основы растений. Состоит из глюкозных остатков, соединенных, в отличие от крахмала, не α , а β(1 4) гликозидной связью. Многие млекопитающие, в том числе и человек не способны усваивать целлюлозу, т. к. они лишены фермента, расщепляющего такого рода гликозидную связь.
 Гликоген полисахарид, в виде которого углеводы запасаются в организме животных. Его часто называют животным крахмалом. Гликоген характеризуется более разветвленной структурой, чем амилопектин. 1 6 связи в гликогене располагаются через каждые 8 11 глюкозных остатков, соединенных 1 4 связью.
Гликоген полисахарид, в виде которого углеводы запасаются в организме животных. Его часто называют животным крахмалом. Гликоген характеризуется более разветвленной структурой, чем амилопектин. 1 6 связи в гликогене располагаются через каждые 8 11 глюкозных остатков, соединенных 1 4 связью.
 ГЕТЕРОПОЛИСАХАРИДЫ: гликозаминогликаны или мукополисахариды: гиалуроновая кислота, хондроитинсульфаты, гепарин. Это основное скрепляющее вещество соединительной ткани. Их функция состоит в удержании большого количества воды и в заполнении межклеточного пространства. Они служат смягчающим и смазочным материалом для разного рода тканевых структур
ГЕТЕРОПОЛИСАХАРИДЫ: гликозаминогликаны или мукополисахариды: гиалуроновая кислота, хондроитинсульфаты, гепарин. Это основное скрепляющее вещество соединительной ткани. Их функция состоит в удержании большого количества воды и в заполнении межклеточного пространства. Они служат смягчающим и смазочным материалом для разного рода тканевых структур
 • ОБМЕН УГЛЕВОДОВ • Переваривание. • Большая часть утилизируемых углеводов поступает в организм с пищей в виде крахмалов (амилозы и амилопектина), гликогена, либо в виде дисахаридов (сахарозы, мальтозы или лактозы). Слизистая оболочка желудочно кишечного тракта играет роль естественного барьера, препятствующего поступлению в нормальный организм крупных чужеродных молекул.
• ОБМЕН УГЛЕВОДОВ • Переваривание. • Большая часть утилизируемых углеводов поступает в организм с пищей в виде крахмалов (амилозы и амилопектина), гликогена, либо в виде дисахаридов (сахарозы, мальтозы или лактозы). Слизистая оболочка желудочно кишечного тракта играет роль естественного барьера, препятствующего поступлению в нормальный организм крупных чужеродных молекул.
 • Переваривание это совокупность процессов ферментативного гидролиза больших молекул полисахаридов, белков, жиров, нуклеиновых кислот, поступающих с пищей, до более мелких молекул, которые могут затем всасываться и далее подвергаться дальнейшим метаболическим превращениям. Различают полостное пищеварение и пристеночное.
• Переваривание это совокупность процессов ферментативного гидролиза больших молекул полисахаридов, белков, жиров, нуклеиновых кислот, поступающих с пищей, до более мелких молекул, которые могут затем всасываться и далее подвергаться дальнейшим метаболическим превращениям. Различают полостное пищеварение и пристеночное.
 • ПОЛОСТНОЕ ПИЩЕВАРЕНИЕ • Переваривание полисахаридов начинается уже в ротовой полости. Здесь они подвергаются действию фермента амилазы слюны. Крахмал распадается на декстрины различной сложности. • Амилазу слюны активирует Na. Cl. Амилаза имеет оптимум р. Н в щелочной среде. Поэтому в желудке, где имеется резко кислая среда, переваривание крахмала возможно только в глубине пищевого комка.
• ПОЛОСТНОЕ ПИЩЕВАРЕНИЕ • Переваривание полисахаридов начинается уже в ротовой полости. Здесь они подвергаются действию фермента амилазы слюны. Крахмал распадается на декстрины различной сложности. • Амилазу слюны активирует Na. Cl. Амилаза имеет оптимум р. Н в щелочной среде. Поэтому в желудке, где имеется резко кислая среда, переваривание крахмала возможно только в глубине пищевого комка.
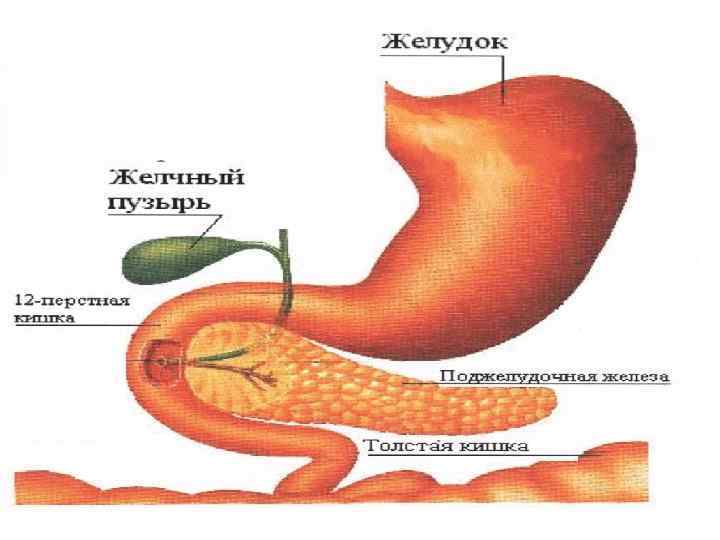
 • Далее пищевой комок переходит в кишечник, где он подвергается действию амилазы поджелудочной железы. Различают альфа -, бета- и гаммаамилазы. • Альфа-амилаза более широко распространена, она расщепляет крахмал до декстринов. • Бета-амилаза расщепляет дальше декстрины до дисахарида мальтозы. • Гамма-амилаза отличается тем, что отщепляет отдельные концевые молекулы глюкозы от молекулы
• Далее пищевой комок переходит в кишечник, где он подвергается действию амилазы поджелудочной железы. Различают альфа -, бета- и гаммаамилазы. • Альфа-амилаза более широко распространена, она расщепляет крахмал до декстринов. • Бета-амилаза расщепляет дальше декстрины до дисахарида мальтозы. • Гамма-амилаза отличается тем, что отщепляет отдельные концевые молекулы глюкозы от молекулы
 • • ПРИСТЕНОЧНОЕ ПИЩЕВАРЕНИЕ Дисахариды также подвергаются расщеплению в ЖКТ: на них действуют ферменты лактаза, сахараза и мальтаза. При этом образуются моносахариды глюкоза, галактоза, фруктоза. Гидролиз дисахаридов происходит не в просвете кишечника, а на поверхности клетках слизистой оболочки под специальной пленочкой гликокаликсом
• • ПРИСТЕНОЧНОЕ ПИЩЕВАРЕНИЕ Дисахариды также подвергаются расщеплению в ЖКТ: на них действуют ферменты лактаза, сахараза и мальтаза. При этом образуются моносахариды глюкоза, галактоза, фруктоза. Гидролиз дисахаридов происходит не в просвете кишечника, а на поверхности клетках слизистой оболочки под специальной пленочкой гликокаликсом
 • Лактоза поступает в организм в составе молока. Иногда в организме ребенка бывает недостаточная активность лактазы. • В этом случае лактоза, которой очень много в женском молоке, не успевает перевариваться, задерживается в кишечнике и может поддерживать развитие мощной и нежелательной микрофлоры кишечника.
• Лактоза поступает в организм в составе молока. Иногда в организме ребенка бывает недостаточная активность лактазы. • В этом случае лактоза, которой очень много в женском молоке, не успевает перевариваться, задерживается в кишечнике и может поддерживать развитие мощной и нежелательной микрофлоры кишечника.
 • При этом у ребенка развивается диспепсия ("несварение", приводящее к поносам, рвоте). Во всех этих случаях желателен переход на кормление коровьим молоком или "смесями", включающими сахарозу • Такие же явления бывают и у взрослых при недостаточности лактазы. • Большой осмотический эффект не всосавшейся лактозы вызывает приток жидкости в тонкий кишечник. Поэтому клиническими симптомами непереносимости лактозы (а стало быть и молока) является вздутие живота, тошнота, судороги, боль и понос.
• При этом у ребенка развивается диспепсия ("несварение", приводящее к поносам, рвоте). Во всех этих случаях желателен переход на кормление коровьим молоком или "смесями", включающими сахарозу • Такие же явления бывают и у взрослых при недостаточности лактазы. • Большой осмотический эффект не всосавшейся лактозы вызывает приток жидкости в тонкий кишечник. Поэтому клиническими симптомами непереносимости лактозы (а стало быть и молока) является вздутие живота, тошнота, судороги, боль и понос.
 Их перенос в клетки слизистой оболочки") • Всасываются в кишечнике только моносахара. 1) Их перенос в клетки слизистой оболочки кишечника знтероциты может происходить по градиенту концентрации, т. е. из кишечника, где концентрация сахара выше в клетки кишечника, где она ниже. Этот вид транспорта называется пассивная диффузия. • 2) Перенос против градиента концентрации. Такой транспорт называется активным, он происходит с затратой энергии и с помощью специальных переносчиков. Глюкоза Белок–переносчик + АТФ
• Всасываются в кишечнике только моносахара. 1) Их перенос в клетки слизистой оболочки кишечника знтероциты может происходить по градиенту концентрации, т. е. из кишечника, где концентрация сахара выше в клетки кишечника, где она ниже. Этот вид транспорта называется пассивная диффузия. • 2) Перенос против градиента концентрации. Такой транспорт называется активным, он происходит с затратой энергии и с помощью специальных переносчиков. Глюкоза Белок–переносчик + АТФ
 • ДАЛЬНЕЙШИЕ ПРЕВРАЩЕНИЯ САХАРОВ В ОРГАНИЗМЕ. • В результате переваривания углеводного компонента пищи основным моносахаридом оказывается глюкоза. Она из кишечника поступает в кровь воротой вены, а затем с током крови в печень. • Большая часть глюкозы задерживается в печени (около 90%), а небольшая часть через общий кровоток может поступать в другие органы и ткани
• ДАЛЬНЕЙШИЕ ПРЕВРАЩЕНИЯ САХАРОВ В ОРГАНИЗМЕ. • В результате переваривания углеводного компонента пищи основным моносахаридом оказывается глюкоза. Она из кишечника поступает в кровь воротой вены, а затем с током крови в печень. • Большая часть глюкозы задерживается в печени (около 90%), а небольшая часть через общий кровоток может поступать в другие органы и ткани
 Пассивная диффузия, т.") • Транспорт глюкозы в клетки осуществляется несколькими способами. • 1) Пассивная диффузия, т. е. прохождение глюкозы через клеточную мембрану по градиенту концентрации по механизму простой диффузии • Благодаря пассивной диффузии глюкоза очень быстро переходит в печень, в крови концентрация глюкозы после приема пищи быстро снижается.
• Транспорт глюкозы в клетки осуществляется несколькими способами. • 1) Пассивная диффузия, т. е. прохождение глюкозы через клеточную мембрану по градиенту концентрации по механизму простой диффузии • Благодаря пассивной диффузии глюкоза очень быстро переходит в печень, в крови концентрация глюкозы после приема пищи быстро снижается.
 Облегченная диффузия. Отличается от пассивной диффузии тем, что в этом случае") • 2) Облегченная диффузия. Отличается от пассивной диффузии тем, что в этом случае перенос осуществляется за счет уникальных белков переносчиков. • Роль этих белков заключается в том, чтобы провести гидрофильное вещество через гидрофобный слой мембраны. Для этого вида транспорта характерна высокая избирательность. • Белки транспортеры делятся на 5 типов: • Глют 1 обеспечивает стабильный поток глюкозы в мозг.
• 2) Облегченная диффузия. Отличается от пассивной диффузии тем, что в этом случае перенос осуществляется за счет уникальных белков переносчиков. • Роль этих белков заключается в том, чтобы провести гидрофильное вещество через гидрофобный слой мембраны. Для этого вида транспорта характерна высокая избирательность. • Белки транспортеры делятся на 5 типов: • Глют 1 обеспечивает стабильный поток глюкозы в мозг.
 • Глют 2 обнаружен в клетках органов, выделяющих глюкозу в кровь: в энтероцитах и печени. • Глют 3 обладает более высоким сродством к глюкозе, чем Глют 1. Обеспечивает поток глюкозы в нервную ткань. • Глют 4 (инсулинзависимый) главный переносчик глюкозы в клетки мышцы и жировой тканей под влиянием инсулина. • Глют 5 встречается в энтероцитах, его функции известны недостаточно.
• Глют 2 обнаружен в клетках органов, выделяющих глюкозу в кровь: в энтероцитах и печени. • Глют 3 обладает более высоким сродством к глюкозе, чем Глют 1. Обеспечивает поток глюкозы в нервную ткань. • Глют 4 (инсулинзависимый) главный переносчик глюкозы в клетки мышцы и жировой тканей под влиянием инсулина. • Глют 5 встречается в энтероцитах, его функции известны недостаточно.
Активный транспорт. Он осуществляется также специфическими белками– переносчиками, но это энергозависимый процесс и требует") 3)Активный транспорт. Он осуществляется также специфическими белками– переносчиками, но это энергозависимый процесс и требует для своего осуществления участия Na+ и К+. Он происходит против градиента концентрации.
3)Активный транспорт. Он осуществляется также специфическими белками– переносчиками, но это энергозависимый процесс и требует для своего осуществления участия Na+ и К+. Он происходит против градиента концентрации.
 В период между приемами пищи концентрация глюкозы в крови в норме от 80 до 100 мг/100 мл или, в современных единицах измерения от 3, 3 до 5, 5 ммоль/л крови. После приема пищи, богатой углеводами, концентрация глюкозы в крови быстро увеличивается примерно на 30 50%, т. е. до 120 150 мг/100 мл (или до 6, 5 7 ммоль/л). Затем довольно быстро начинает снижаться и к 2 ч после приема пищи возвращается к исходному состоянию.
В период между приемами пищи концентрация глюкозы в крови в норме от 80 до 100 мг/100 мл или, в современных единицах измерения от 3, 3 до 5, 5 ммоль/л крови. После приема пищи, богатой углеводами, концентрация глюкозы в крови быстро увеличивается примерно на 30 50%, т. е. до 120 150 мг/100 мл (или до 6, 5 7 ммоль/л). Затем довольно быстро начинает снижаться и к 2 ч после приема пищи возвращается к исходному состоянию.
 Количество поступившей с пищей глюкозы, скорость и") От каких факторов зависит концентрация глюкозы 1) Количество поступившей с пищей глюкозы, скорость и регулярность поступления глюкозы. 2) Скорость поступления глюкозы в клетки, скорость транспорта. 3) Скорость утилизации глюкозы, т. е. : а) Скорость синтеза гликогена и скорость распада гликогена – гликогенолиза; б) Интенсивность гликолиза и глюконеогенеза
От каких факторов зависит концентрация глюкозы 1) Количество поступившей с пищей глюкозы, скорость и регулярность поступления глюкозы. 2) Скорость поступления глюкозы в клетки, скорость транспорта. 3) Скорость утилизации глюкозы, т. е. : а) Скорость синтеза гликогена и скорость распада гликогена – гликогенолиза; б) Интенсивность гликолиза и глюконеогенеза
 ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ УГЛЕВОДНОГО ОБМЕНА Гормоны надпочечников АДРЕНАЛИН и ГЛЮКОКОРТИКОИДНЫЕ гормоны, гормон щитовидной железы ТИРОКСИН, гормоны поджелудочной железы ИНСУЛИН и ГЛЮКАГОН Только один из этих гормонов СНИЖАЕТ концентрацию сахара в крови. Это ИНСУЛИН. Все остальные из перечисленных гормонов являются КОНТРГОРМОНАМИ по отношению к инсулину и обладают противоположным действием, т. е. ПОВЫШАЮТ содержание сахара в крови, но каждый с помощью своего собственного механизма действия.
ГОРМОНАЛЬНАЯ РЕГУЛЯЦИЯ УГЛЕВОДНОГО ОБМЕНА Гормоны надпочечников АДРЕНАЛИН и ГЛЮКОКОРТИКОИДНЫЕ гормоны, гормон щитовидной железы ТИРОКСИН, гормоны поджелудочной железы ИНСУЛИН и ГЛЮКАГОН Только один из этих гормонов СНИЖАЕТ концентрацию сахара в крови. Это ИНСУЛИН. Все остальные из перечисленных гормонов являются КОНТРГОРМОНАМИ по отношению к инсулину и обладают противоположным действием, т. е. ПОВЫШАЮТ содержание сахара в крови, но каждый с помощью своего собственного механизма действия.
 ИНСУЛИН. Инсулин белковый гормон. Молекулярная масса = 6000, содержит 51 аминокислоту и состоит из двух цепей, соединенных дисульфидными мостиками: ! S S ! глицин 6 7 11 20 А (21 АК) ! ! 6, 7, 11 и 20 цистеин S S ! ! 7 и 19 цистеин Фен 7 19 В (30 АК)
ИНСУЛИН. Инсулин белковый гормон. Молекулярная масса = 6000, содержит 51 аминокислоту и состоит из двух цепей, соединенных дисульфидными мостиками: ! S S ! глицин 6 7 11 20 А (21 АК) ! ! 6, 7, 11 и 20 цистеин S S ! ! 7 и 19 цистеин Фен 7 19 В (30 АК)
 ИНСУЛИН ТРАНСПОРТ ГЛЮКО НЕОГЕНЕЗ ГЛЮКО КИНАЗА ГЛИКОЛИЗ СИНТЕЗ ГЛИКОГЕНА ГЛИКОГЕ НОЛИЗ ПФП ЛИПОГЕНЕЗ СИНТЕЗ БЕЛКА ЛИПОЛИЗ
ИНСУЛИН ТРАНСПОРТ ГЛЮКО НЕОГЕНЕЗ ГЛЮКО КИНАЗА ГЛИКОЛИЗ СИНТЕЗ ГЛИКОГЕНА ГЛИКОГЕ НОЛИЗ ПФП ЛИПОГЕНЕЗ СИНТЕЗ БЕЛКА ЛИПОЛИЗ
 Дальнейшая судьба глюкозы в организме • В клетках глюкоза прежде всего подвергается реакции фосфорилирования. • В клетках печени существует два изофермента, которые могут фосфорилировать глюкозу: это глюкокиназа и гексокиназа. В других органах есть только гексокиназа. • Под влиянием глюкокиназы происходит фосфорилирование образованием Г 6 Ф. глюкозы с
Дальнейшая судьба глюкозы в организме • В клетках глюкоза прежде всего подвергается реакции фосфорилирования. • В клетках печени существует два изофермента, которые могут фосфорилировать глюкозу: это глюкокиназа и гексокиназа. В других органах есть только гексокиназа. • Под влиянием глюкокиназы происходит фосфорилирование образованием Г 6 Ф. глюкозы с
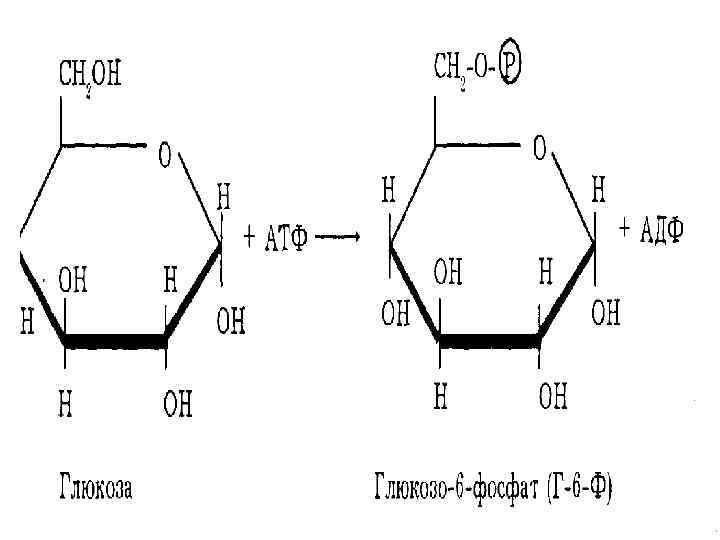
 глюкозо 6 фосфат + АДФ:") глюкоза + АТФ Глюкокиназа (Гексокиназа) глюкозо 6 фосфат + АДФ:
глюкоза + АТФ Глюкокиназа (Гексокиназа) глюкозо 6 фосфат + АДФ:
 ГЛИКОЛИЗ Реакции этого пути протекают цитоплазме клетки. в 1 я реакция гликолиза это получение Г 6 -Ф. с помощью фермента гексокиназы или глюкокиназы. При этом затрачивается 1 молекула АТФ. В другом случае, когда глюкоза отщепляется от гликогена в виде Г 1 Ф, она превращается в Г 6 Ф с помощью фермента фосфоглюкомутазы, и в этом случае расхода АТФ нет.
ГЛИКОЛИЗ Реакции этого пути протекают цитоплазме клетки. в 1 я реакция гликолиза это получение Г 6 -Ф. с помощью фермента гексокиназы или глюкокиназы. При этом затрачивается 1 молекула АТФ. В другом случае, когда глюкоза отщепляется от гликогена в виде Г 1 Ф, она превращается в Г 6 Ф с помощью фермента фосфоглюкомутазы, и в этом случае расхода АТФ нет.
 2 я реакция гликолиза это изомеризация Г 6 Ф во фруктозо 6 Ф (Ф 6 Ф). Фермент – глюкозофосфатизомераза. 3 я реакция гликолиза второе фосфорилирование. Ф 6 Ф фосфорилируется с образованием Ф 1, 6 дифосфата под влиянием фермента фосфофруктокиназы. При этом затрачивается еще одна молекула АТФ.
2 я реакция гликолиза это изомеризация Г 6 Ф во фруктозо 6 Ф (Ф 6 Ф). Фермент – глюкозофосфатизомераза. 3 я реакция гликолиза второе фосфорилирование. Ф 6 Ф фосфорилируется с образованием Ф 1, 6 дифосфата под влиянием фермента фосфофруктокиназы. При этом затрачивается еще одна молекула АТФ.
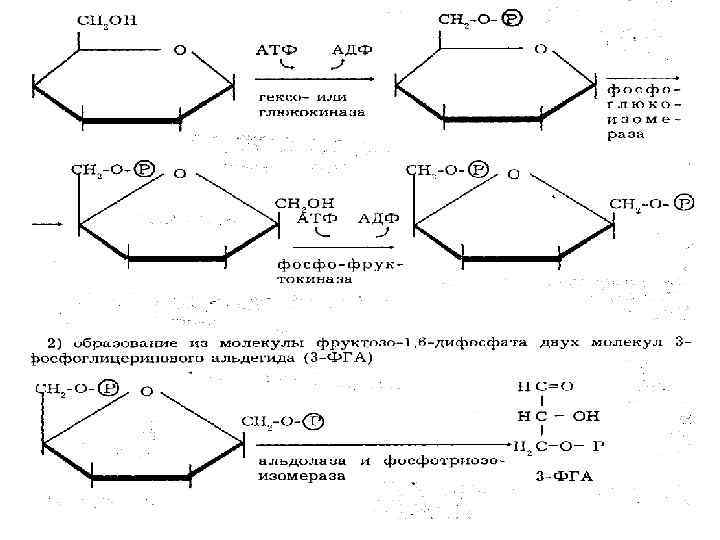
 Весь ход гликолиза лимитируется уровнем активности этого фермента. Это аллостерический фермент. Его активность ингибируется АТФ, лимонной кислотой, жирными кислотами. Активация под влиянием АДФ и АМФ. Образованием Ф 1, 6 ДФ первая стадия заканчивается. гликолиза
Весь ход гликолиза лимитируется уровнем активности этого фермента. Это аллостерический фермент. Его активность ингибируется АТФ, лимонной кислотой, жирными кислотами. Активация под влиянием АДФ и АМФ. Образованием Ф 1, 6 ДФ первая стадия заканчивается. гликолиза
 • Далее под влиянием фермента альдолазы происходит расщепление Ф 1, 6 ДФ с образованием двух триоз: • 3 Ф глицеральдегида (3 ФГА) и диоксиацетон Ф (ДОФ). • В дальнейшем в реакциях гликолиза уже участвуют не 6 ти углеродные, а трехуглеродные соединения.
• Далее под влиянием фермента альдолазы происходит расщепление Ф 1, 6 ДФ с образованием двух триоз: • 3 Ф глицеральдегида (3 ФГА) и диоксиацетон Ф (ДОФ). • В дальнейшем в реакциях гликолиза уже участвуют не 6 ти углеродные, а трехуглеродные соединения.
 Диоксиацетон Ф превращается в 3 Ф Глицеральдегид. • СН 2 ОН НСО • СО НСОН • СН 2 ОРО 3 Н 2 • дигидрооксиацетон Ф 3 Ф глицеральдегид Таким образом, из 1 молекулы Ф 1, 6 ДФ образуются две молекулы 3 Ф глицеральдегид.
Диоксиацетон Ф превращается в 3 Ф Глицеральдегид. • СН 2 ОН НСО • СО НСОН • СН 2 ОРО 3 Н 2 • дигидрооксиацетон Ф 3 Ф глицеральдегид Таким образом, из 1 молекулы Ф 1, 6 ДФ образуются две молекулы 3 Ф глицеральдегид.
 • П Е Р Ы В
• П Е Р Ы В
 • Начиная с третьей стадии, происходят реакции, которые направлены на получение энергии. • 6 я реакция это ключевая реакция гликолиза. Здесь происходит окисление 3 Ф глицеральдегида до 1, 3 дифосфоглицериновой кислоты.
• Начиная с третьей стадии, происходят реакции, которые направлены на получение энергии. • 6 я реакция это ключевая реакция гликолиза. Здесь происходит окисление 3 Ф глицеральдегида до 1, 3 дифосфоглицериновой кислоты.
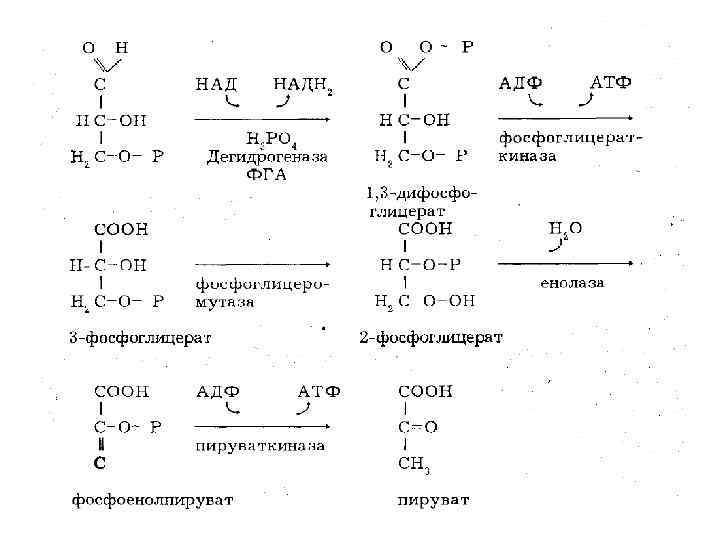
 • Cледующая реакция – также чрезвычайно важная, т. к. здесь происходит образование АТФ без участия окислительного фосфорилирования. Это пример фосфорилирования на субстратном уровне.
• Cледующая реакция – также чрезвычайно важная, т. к. здесь происходит образование АТФ без участия окислительного фосфорилирования. Это пример фосфорилирования на субстратном уровне.
 • О • СОФ СНОН • 2 СНОН +АДФ 2 СНОН + АТФ • СН 2 ОФ • 1, 3 ДФГК 3 ФГК • При этом 1, 3 ДФГК под влиянием фосфоглицерокиназы теряет один остаток фосфорной кислоты, который уходит на образование АТФ, а сама превращается в 3 Фосфоглицериновую кислоту (3 ФГК)
• О • СОФ СНОН • 2 СНОН +АДФ 2 СНОН + АТФ • СН 2 ОФ • 1, 3 ДФГК 3 ФГК • При этом 1, 3 ДФГК под влиянием фосфоглицерокиназы теряет один остаток фосфорной кислоты, который уходит на образование АТФ, а сама превращается в 3 Фосфоглицериновую кислоту (3 ФГК)
 • СООН • СНОН глицеромутаза НСО Ф • СН 2 О Ф СН 2 ОН В следующей реакции 3 фосфоглицерат превращается в 2 фосфоглицерат под влиянием фермента фосфоглицеромутазы. При этом происходит переход фосфорильной группы из 3 его положения во 2 е:
• СООН • СНОН глицеромутаза НСО Ф • СН 2 О Ф СН 2 ОН В следующей реакции 3 фосфоглицерат превращается в 2 фосфоглицерат под влиянием фермента фосфоглицеромутазы. При этом происходит переход фосфорильной группы из 3 его положения во 2 е:
 • 9 я реакция состоит в образовании фосфоенолпирувата в результате дегидратации 2 фосфоглицерата. Фермент енолаза. • СООН • 2 НСО Ф 2 НОН 2 СО Ф • СН !! • СН 2 •
• 9 я реакция состоит в образовании фосфоенолпирувата в результате дегидратации 2 фосфоглицерата. Фермент енолаза. • СООН • 2 НСО Ф 2 НОН 2 СО Ф • СН !! • СН 2 •
 • СООН • 2 СО Ф + 2 АДФ 2 С=О + АТФ • !! пируваткиназа СН 3 • СН 2 • ФЭП ПВК • 10 я, последняя, реакция гликолиза образование пирувата с одновременным образованием АТФ. Перенос фосфорильной группы от фосфоенолпирувата к АДФ катализируется пируваткиназой. Это также реакция субстратного фосфорилирования.
• СООН • 2 СО Ф + 2 АДФ 2 С=О + АТФ • !! пируваткиназа СН 3 • СН 2 • ФЭП ПВК • 10 я, последняя, реакция гликолиза образование пирувата с одновременным образованием АТФ. Перенос фосфорильной группы от фосфоенолпирувата к АДФ катализируется пируваткиназой. Это также реакция субстратного фосфорилирования.
 Таким образом, в ходе превращения глюкозы в ПВК происходит образование 4 молекул АТФ: 2 молекулы АТФ освобождаются при окислении двух молекул 1, 3 дифосфоглицерата в 3 фосфоглицерат, и еще 2 молекулы АТФ образуются при превращении фосфоенолпирувата в ПВК.
Таким образом, в ходе превращения глюкозы в ПВК происходит образование 4 молекул АТФ: 2 молекулы АТФ освобождаются при окислении двух молекул 1, 3 дифосфоглицерата в 3 фосфоглицерат, и еще 2 молекулы АТФ образуются при превращении фосфоенолпирувата в ПВК.
 • Но из этих 4 молекул АТФ нужно вычесть 1 молекулу АТФ, которая затрачивается на фосфорилирование глюкозы, и еще 1 молекулу АТФ, которая затрачивается на образование Ф 1, 6 ДФ. Поэтому общий итог 2 АТФ. Это и есть выигрыш гликолиза. При распаде гликогена выигрыш 3 молекулы АТФ, т. к. нет расхода на фосфорилирование глюкозы.
• Но из этих 4 молекул АТФ нужно вычесть 1 молекулу АТФ, которая затрачивается на фосфорилирование глюкозы, и еще 1 молекулу АТФ, которая затрачивается на образование Ф 1, 6 ДФ. Поэтому общий итог 2 АТФ. Это и есть выигрыш гликолиза. При распаде гликогена выигрыш 3 молекулы АТФ, т. к. нет расхода на фосфорилирование глюкозы.
 ЖИРЫ БЕЛКИ УГЛЕВОДЫ Аминокислоты ГЛЮКОЗА ПВК ГЛИЦЕРИН ПВК ВЖК Ацетил Ко. А КЕТОКИСЛОТЫ Ацетил Ко. А Цикл Кребса
ЖИРЫ БЕЛКИ УГЛЕВОДЫ Аминокислоты ГЛЮКОЗА ПВК ГЛИЦЕРИН ПВК ВЖК Ацетил Ко. А КЕТОКИСЛОТЫ Ацетил Ко. А Цикл Кребса
 • В матриксе содержатся ферменты окислительного декарбоксилирования ПВК и ферменты ЦТК. • Ферменты же дыхательной цепи встроены во внутреннюю митохондриальную мембрану и важны для всего процесса терминального окисления и, в особенности, для окислительного фосфорилирования.
• В матриксе содержатся ферменты окислительного декарбоксилирования ПВК и ферменты ЦТК. • Ферменты же дыхательной цепи встроены во внутреннюю митохондриальную мембрану и важны для всего процесса терминального окисления и, в особенности, для окислительного фосфорилирования.
 • Рассмотрим каждый из этапов в отдельности. I. В реакциях промежуточного обмена наибольшее значение имеет процесс образования ацетил Ко. А из пировиноградной кислоты в окислительного декарбоксилирования • реакции
• Рассмотрим каждый из этапов в отдельности. I. В реакциях промежуточного обмена наибольшее значение имеет процесс образования ацетил Ко. А из пировиноградной кислоты в окислительного декарбоксилирования • реакции
 • Реакция протекает в матриксе митохондрий, служит связующим звено между реакциями окисления глюкозы и циклом Кребса. Суммарно реакцию можно записать так: ПВК + Ко. А + НАД+ Ацетил Ко. А + СО 2 + НАДН. • Катализируется пируват дегидрогеназным комплексом. Это типичный пример мультиферментного комплекса.
• Реакция протекает в матриксе митохондрий, служит связующим звено между реакциями окисления глюкозы и циклом Кребса. Суммарно реакцию можно записать так: ПВК + Ко. А + НАД+ Ацетил Ко. А + СО 2 + НАДН. • Катализируется пируват дегидрогеназным комплексом. Это типичный пример мультиферментного комплекса.
 На") • Превращение пирувата в ацетил Ко. А происходит в 4 стадии 1) На первой стадии пируват соединяется с тиаминпирофосфатом (ТПФ) и затем подвергается декарбоксилированию. В результате образуется оксиэтил ТПФ. Фермент • пируват дегидрогеназа
• Превращение пирувата в ацетил Ко. А происходит в 4 стадии 1) На первой стадии пируват соединяется с тиаминпирофосфатом (ТПФ) и затем подвергается декарбоксилированию. В результате образуется оксиэтил ТПФ. Фермент • пируват дегидрогеназа
 На второй стадии оксиэтил ТПФ окисляется с образованием ацетильной группы и") • 2) На второй стадии оксиэтил ТПФ окисляется с образованием ацетильной группы и одновременно переносится на липоевую кислоту (липоамид). •
• 2) На второй стадии оксиэтил ТПФ окисляется с образованием ацетильной группы и одновременно переносится на липоевую кислоту (липоамид). •
 На третьей стадии ацетильная группа переносится с ацетиллипоамида на Ко. А,") • 3) На третьей стадии ацетильная группа переносится с ацетиллипоамида на Ко. А, образуя ацетил~Ко. А. Процесс также катализируется дигидролипоилтрансацетилазой. При переходе ацетильной группы на Ко. А сохраняется богатая энергией тиоэфирная связь: ацетил~Ко. А
• 3) На третьей стадии ацетильная группа переносится с ацетиллипоамида на Ко. А, образуя ацетил~Ко. А. Процесс также катализируется дигидролипоилтрансацетилазой. При переходе ацетильной группы на Ко. А сохраняется богатая энергией тиоэфирная связь: ацетил~Ко. А
 На четвертой, завершающей стадии происходит восстановление • окисленной формы липоамида. Реакция") • 4) На четвертой, завершающей стадии происходит восстановление • окисленной формы липоамида. Реакция катализируется дигидролипоил дегидрогеназой. Окислителем в ней служит НАД+, а роль кофермента выполняет ФАД: • SH S • ЛК + ФАДН 2 • SH S • • ФАДН 2 + НАД+ ФАД + НАДН 2 •
• 4) На четвертой, завершающей стадии происходит восстановление • окисленной формы липоамида. Реакция катализируется дигидролипоил дегидрогеназой. Окислителем в ней служит НАД+, а роль кофермента выполняет ФАД: • SH S • ЛК + ФАДН 2 • SH S • • ФАДН 2 + НАД+ ФАД + НАДН 2 •
В") Биологическое значение окислительного декарбоксилирования ПВК определяется тем, какова будет дальнейшая судьба его продуктов: а)В процессе декарбоксилирования образуется НАДН 2, который может генерировать энергию. 1 мол НАДН 2 вызывает образование 3 мол. АТФ.
Биологическое значение окислительного декарбоксилирования ПВК определяется тем, какова будет дальнейшая судьба его продуктов: а)В процессе декарбоксилирования образуется НАДН 2, который может генерировать энергию. 1 мол НАДН 2 вызывает образование 3 мол. АТФ.
 Дальнейший путь самого ацетил Ко. А может идти по двум направлениям: 1) ацетил") б) Дальнейший путь самого ацетил Ко. А может идти по двум направлениям: 1) ацетил Ко. А включается в цикл трикарбоновых кислот и сгорает в нем до СО 2. При этом образуется огромное количество энергии, либо 2) ацетил Ко. А включается в процесс синтеза липидов
б) Дальнейший путь самого ацетил Ко. А может идти по двум направлениям: 1) ацетил Ко. А включается в цикл трикарбоновых кислот и сгорает в нем до СО 2. При этом образуется огромное количество энергии, либо 2) ацетил Ко. А включается в процесс синтеза липидов
 Цикл начинается с конденсации ЩУК и ацетил Ко. А, при этом образуется 1") 1) Цикл начинается с конденсации ЩУК и ацетил Ко. А, при этом образуется 1 молекула цитрата, HS Co. A и Н+ • • • СООН С=О + СН 3 CО~Ко. А + НОН СН 2 НООС СОН СООН СН 2 (ЩУК) СООН (цитрат) НS~Ко. А + Н+
1) Цикл начинается с конденсации ЩУК и ацетил Ко. А, при этом образуется 1 молекула цитрата, HS Co. A и Н+ • • • СООН С=О + СН 3 CО~Ко. А + НОН СН 2 НООС СОН СООН СН 2 (ЩУК) СООН (цитрат) НS~Ко. А + Н+
Следующая реакция изомеризации лимонной кислоты. При этом сначала под влиянием фермента цисаконитазы образуется цисаконитовая") 2)Следующая реакция изомеризации лимонной кислоты. При этом сначала под влиянием фермента цисаконитазы образуется цисаконитовая форма лимонной кислоты. Затем из цисаконитата образуется изоцитрат. Далее идут реакции, в которых образуются энергетические эквиваленты НАД. Н и ФАД. Н
2)Следующая реакция изомеризации лимонной кислоты. При этом сначала под влиянием фермента цисаконитазы образуется цисаконитовая форма лимонной кислоты. Затем из цисаконитата образуется изоцитрат. Далее идут реакции, в которых образуются энергетические эквиваленты НАД. Н и ФАД. Н
Третья реакция ЦТК: окислительное декар боксилирование изоцитрата, (фермент – изоцитратдегидрогеназа). Промежуточный продукт щавелевоянтарная кислота") 3)Третья реакция ЦТК: окислительное декар боксилирование изоцитрата, (фермент – изоцитратдегидрогеназа). Промежуточный продукт щавелевоянтарная кислота (оксалосукцинат), • которая быстро теряет СО 2 с образованием альфа кетоглутаровой кислоты:
3)Третья реакция ЦТК: окислительное декар боксилирование изоцитрата, (фермент – изоцитратдегидрогеназа). Промежуточный продукт щавелевоянтарная кислота (оксалосукцинат), • которая быстро теряет СО 2 с образованием альфа кетоглутаровой кислоты:
 реакция окислительное декарбоксилирование альфа кетоглутаровой кислоты (мультиферментный альфа кетоглутарат дегидрогеназный комплекс) с") Следующая (4) реакция окислительное декарбоксилирование альфа кетоглутаровой кислоты (мультиферментный альфа кетоглутарат дегидрогеназный комплекс) с образованием сукцинил Ко. А с высокоэнергетической фосфатной связью. (5). Сукцинил~Ко. А сукцинат под влиянием сукцинил~Ко. А синтетазы. Эта реакция сопряжена с фосфорилированием гуанозиндифосфата. Это пример субстратного фосфорилирования. ГТФ легко превращается в АТФ.
Следующая (4) реакция окислительное декарбоксилирование альфа кетоглутаровой кислоты (мультиферментный альфа кетоглутарат дегидрогеназный комплекс) с образованием сукцинил Ко. А с высокоэнергетической фосфатной связью. (5). Сукцинил~Ко. А сукцинат под влиянием сукцинил~Ко. А синтетазы. Эта реакция сопряжена с фосфорилированием гуанозиндифосфата. Это пример субстратного фосфорилирования. ГТФ легко превращается в АТФ.
 оследняя стадия ЦТК включает 3 реакции, благодаря которым происходит образование ЩУК из сукцината через фумарат и малат: 6) Сначала сукцинат окисляется в фумарат с помощью фермента сукцинат ДГ (СДГ). Акцептором водорода здесь является ФАД. Н. Гидратация фумарата под влиянием фермента фумаратгидратазы с образованием L малата. 8) Яблочная кислота окисляется с помощью малат дегидрогеназы (МДГ), где роль акцептора электронов выполняет НАД+. Продукты реакции ЩУК, НАДН и Н+:
оследняя стадия ЦТК включает 3 реакции, благодаря которым происходит образование ЩУК из сукцината через фумарат и малат: 6) Сначала сукцинат окисляется в фумарат с помощью фермента сукцинат ДГ (СДГ). Акцептором водорода здесь является ФАД. Н. Гидратация фумарата под влиянием фермента фумаратгидратазы с образованием L малата. 8) Яблочная кислота окисляется с помощью малат дегидрогеназы (МДГ), где роль акцептора электронов выполняет НАД+. Продукты реакции ЩУК, НАДН и Н+:
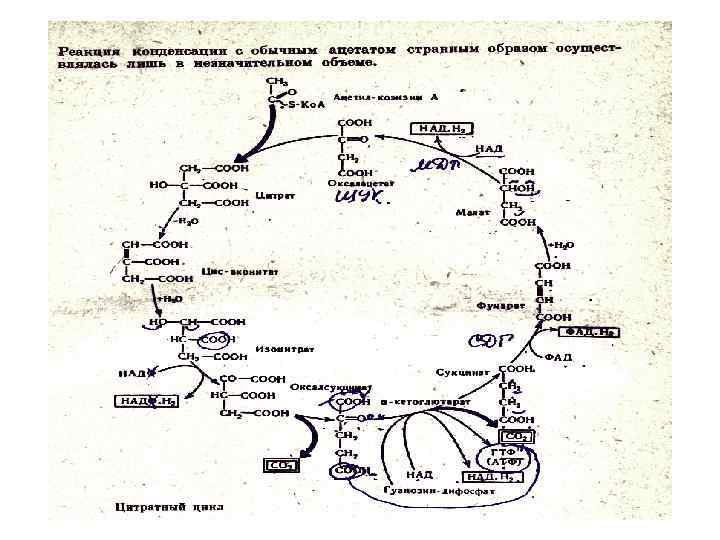
 Суммарно уравнение ЦТК будет выглядеть следующим образом: CH 3 CO~SKo. A + ЩУК +ГДФ +Н 3 РО 4 + 3 НАД+ +ФАД + 2 Н 2 О ЩУК+2 СО 2+ГТФ(АТФ)+3 НАДН+ФАДН При сгорании 1 мол Ацетил Ко. А образуется 12 мол АТФ.
Суммарно уравнение ЦТК будет выглядеть следующим образом: CH 3 CO~SKo. A + ЩУК +ГДФ +Н 3 РО 4 + 3 НАД+ +ФАД + 2 Н 2 О ЩУК+2 СО 2+ГТФ(АТФ)+3 НАДН+ФАДН При сгорании 1 мол Ацетил Ко. А образуется 12 мол АТФ.
 • Какова энергетика ЦТК? В ЦТК за период одного цикла сгорает 1 молекула ацетил~Ко. А. При этом образуются три молекулы НАДН 2 и одна молекула ФАДН 2. Эти восстановительные эквиваленты передаются в дыхательную цепь, локализованную во внутренней митохондриальной мембране. При прохождении по дыхательной цепи НАДН 2 образуются три молекулы АТФ из АДФ. За счет ФАДН 2 образуются только две молекулы АТФ
• Какова энергетика ЦТК? В ЦТК за период одного цикла сгорает 1 молекула ацетил~Ко. А. При этом образуются три молекулы НАДН 2 и одна молекула ФАДН 2. Эти восстановительные эквиваленты передаются в дыхательную цепь, локализованную во внутренней митохондриальной мембране. При прохождении по дыхательной цепи НАДН 2 образуются три молекулы АТФ из АДФ. За счет ФАДН 2 образуются только две молекулы АТФ
 • Еще один высокоэнер гетический фосфат производится на субстратном уровне при превращении сукцинил~Ко. А в сукцинат. Таким образом, всего за период каждого цикла образуется 12 молекул АТФ.
• Еще один высокоэнер гетический фосфат производится на субстратном уровне при превращении сукцинил~Ко. А в сукцинат. Таким образом, всего за период каждого цикла образуется 12 молекул АТФ.