

РЕАЛИЗАЦИЯ НАСЛЕДСТВЕННОЙ ИНФОРМАЦИИ.ppt
- Количество слайдов: 60

 Основные принципы наследования и наследственности
Основные принципы наследования и наследственности
 Наследственность - это способность организма передавать морфологические, биохимические и функциональные признаки своим потомкам.
Наследственность - это способность организма передавать морфологические, биохимические и функциональные признаки своим потомкам.
,") n Наследственная информация, заключенная в генах каждой особи (совокупность всех генов, присущих данной особи), носит название генотип, идиотип, или генетическая конституция, является итогом исторического развития данного вида и материальной основой будущей эволюции.
n Наследственная информация, заключенная в генах каждой особи (совокупность всех генов, присущих данной особи), носит название генотип, идиотип, или генетическая конституция, является итогом исторического развития данного вида и материальной основой будущей эволюции.
 n n Явление наследственности рассматривает ся как сложная молекулярная внутриклеточная система, обеспечивающая хранение и реализацию информации, в соответствии с которой осуществляются жизнь клетки, развитие особи и ее жизнедеятельности. Реализация наследственной информации, записанной с помощью генетического кода — чередования нуклеотидов в ДНК зиготы, происходит в результате непрерывных взаимовлияний ядра и цитоплазмы, межклеточных взаимодействий и гормональной регуляции активности генов.
n n Явление наследственности рассматривает ся как сложная молекулярная внутриклеточная система, обеспечивающая хранение и реализацию информации, в соответствии с которой осуществляются жизнь клетки, развитие особи и ее жизнедеятельности. Реализация наследственной информации, записанной с помощью генетического кода — чередования нуклеотидов в ДНК зиготы, происходит в результате непрерывных взаимовлияний ядра и цитоплазмы, межклеточных взаимодействий и гормональной регуляции активности генов.
 Наследственность обладает 2 свойствами: n 1. консерватизм или стабильность генетического аппарата, Основа стабильности генома: n n Дублированность его структурных элементов Матричный принцип биосинтеза Способность к репарации Регуляция генной активности
Наследственность обладает 2 свойствами: n 1. консерватизм или стабильность генетического аппарата, Основа стабильности генома: n n Дублированность его структурных элементов Матричный принцип биосинтеза Способность к репарации Регуляция генной активности
 n 2. изменчивость - гарантирует организму необходимую приспособляемость к условиям существования. Часть наследственной изменчивости, которая определяет не нормальные вариации признаков, а патологические, накопившаяся за время эволюции человека, составляет наследственную патологию.
n 2. изменчивость - гарантирует организму необходимую приспособляемость к условиям существования. Часть наследственной изменчивости, которая определяет не нормальные вариации признаков, а патологические, накопившаяся за время эволюции человека, составляет наследственную патологию.
 Каждый наследственный признак определяется отдельным наследственным") Основные принципы наследования признаков: n n n 1) Каждый наследственный признак определяется отдельным наследственным фактором, задатком; в современном представлении эти задатки соответствуют генам; 2) Гены сохраняются в чистом виде в ряду поколений, не утрачивая своей индивидуальности: это явилось доказательством основного положения генетики: ген относительно постоянен; 3) Оба пола в равной мере участвуют в передаче своих наследственных свойств потомству; 4) Редупликация равного числа генов и их редукция в мужских и женских половых клетках; это положение явилось генетическим предвидением существования мейоза; 5) Наследственные задатки являются парными, один – материнский, другой – отцовский; один из них может быть доминантным, другой – рецессивным; это положение соответствует открытию принципа аллелизма: ген представлен минимум двумя аллелями.
Основные принципы наследования признаков: n n n 1) Каждый наследственный признак определяется отдельным наследственным фактором, задатком; в современном представлении эти задатки соответствуют генам; 2) Гены сохраняются в чистом виде в ряду поколений, не утрачивая своей индивидуальности: это явилось доказательством основного положения генетики: ген относительно постоянен; 3) Оба пола в равной мере участвуют в передаче своих наследственных свойств потомству; 4) Редупликация равного числа генов и их редукция в мужских и женских половых клетках; это положение явилось генетическим предвидением существования мейоза; 5) Наследственные задатки являются парными, один – материнский, другой – отцовский; один из них может быть доминантным, другой – рецессивным; это положение соответствует открытию принципа аллелизма: ген представлен минимум двумя аллелями.
 наследственной детерминации признаков;") Законы наследственности n n n Первый закон – закон дискретной (генной) наследственной детерминации признаков; он лежит в основе теории гена. Второй закон – закон относительного постоянства наследственной единицы – гена. Третий закон – закон аллельного состояния гена (доминантность и рецессивность).
Законы наследственности n n n Первый закон – закон дискретной (генной) наследственной детерминации признаков; он лежит в основе теории гена. Второй закон – закон относительного постоянства наследственной единицы – гена. Третий закон – закон аллельного состояния гена (доминантность и рецессивность).

") Моногибридным называется скрещивание двух организмов, отличающихся друг от друга по одной паре альтернативных (взаимоисключающих) признаков.
Моногибридным называется скрещивание двух организмов, отличающихся друг от друга по одной паре альтернативных (взаимоисключающих) признаков.
 Альтернативные признаки
Альтернативные признаки
 G (гаметы) F 1 (потомки) P ♀АА х ♂аа АА -") Закон доминирования P(родители) G (гаметы) F 1 (потомки) P ♀АА х ♂аа АА - желтые G аа - зеленые А а F 1 Аа (желтые) » При скрещивании двух чистых линий у F 1 будет проявляться лишь один альтернативный признак.
Закон доминирования P(родители) G (гаметы) F 1 (потомки) P ♀АА х ♂аа АА - желтые G аа - зеленые А а F 1 Аа (желтые) » При скрещивании двух чистых линий у F 1 будет проявляться лишь один альтернативный признак.
 Моногибридное скрещивание у растений
Моногибридное скрещивание у растений
 Моногибридное скрещивание у животных
Моногибридное скрещивание у животных
 Неполное доминирование – доминантный ген не полностью подавляет рецессивный, а оба аллеля проявляют своё действие.
Неполное доминирование – доминантный ген не полностью подавляет рецессивный, а оба аллеля проявляют своё действие.
 Кроме нормальных доминантных и рецессивных признаков у человека известно большое количество аномальных признаков. Аномалии могут быть как рецессивными, так и доминантными. Примером доминантной аномалии – наследственная близорукость. Близорукость имеет много проявлений и наследуется различно.
Кроме нормальных доминантных и рецессивных признаков у человека известно большое количество аномальных признаков. Аномалии могут быть как рецессивными, так и доминантными. Примером доминантной аномалии – наследственная близорукость. Близорукость имеет много проявлений и наследуется различно.
 Цвет волос у человека определяется взаимодействием нескольких генов. Признак рыжих волос является рецессивным по отношению к нерыжим волосам и проявляется только в гомозиготном состоянии. На картине Гане Эворта «Семейство Кобхем» все члены этой семьи имеют рыжие волосы, т. е. являются гомозиготными по рецессивному признаку окраски волос.
Цвет волос у человека определяется взаимодействием нескольких генов. Признак рыжих волос является рецессивным по отношению к нерыжим волосам и проявляется только в гомозиготном состоянии. На картине Гане Эворта «Семейство Кобхем» все члены этой семьи имеют рыжие волосы, т. е. являются гомозиготными по рецессивному признаку окраски волос.


 Дигибридное скрещивание
Дигибридное скрещивание
 F 1 АА ВВ жёлтые, гладкие семена х аа bb зелёные,") Р G (гаметы) F 1 АА ВВ жёлтые, гладкие семена х аа bb зелёные, морщинистые семена АВ аb Аа Bb жёлтые, гладкие семена 100% Понятия: дигибридное скрещивание, гомозигота, гетерозигота, гаметы, доминантный признак, рецессивный признак, аллельные гены, решётка Пеннета
Р G (гаметы) F 1 АА ВВ жёлтые, гладкие семена х аа bb зелёные, морщинистые семена АВ аb Аа Bb жёлтые, гладкие семена 100% Понятия: дигибридное скрещивание, гомозигота, гетерозигота, гаметы, доминантный признак, рецессивный признак, аллельные гены, решётка Пеннета
 Аа Bb жёлтые, гладкие") F 1 Аа Bb х жёлтые, гладкие семена G (гаметы) Аа Bb жёлтые, гладкие семена AB Ab a. B AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb aabb 9 ж. гл. с. Aabb 3 ж. морщ. с. ab 3 зел. гл. с. 1 зел. морщ с.
F 1 Аа Bb х жёлтые, гладкие семена G (гаметы) Аа Bb жёлтые, гладкие семена AB Ab a. B AB AABb Aa. BB Aa. Bb Ab AABb AAbb Aa. Bb Aabb a. B Aa. Bb aa. BB aa. Bb ab Aa. Bb aabb 9 ж. гл. с. Aabb 3 ж. морщ. с. ab 3 зел. гл. с. 1 зел. морщ с.
 Цитологические основы
Цитологические основы
 – при") I закон Менделя (закон единообразия гибридов первого поколения или правило доминирования ) – при моногибридном скрещивании у гибридов первого поколения проявляются только доминантные признаки – оно фенотипически единообразно II закон Менделя (закон расщепления) – в потомстве, полученном от скрещивания гибридов первого поколения, наблюдается явление расщепления: четверть особей из гибридов второго поколения несёт рецессивный признак, три четверти – доминантный III закон Менделя (закон независимого расщепления или закон независимого комбинирования признаков) – при дигибридном скрещивании у гибридов каждая пара признаков наследуется независимо от других и даёт с ними разные сочетания. Образуются фенотипические группы, характеризующиеся отношением 9: 3: 3: 1 (расщепление по каждой паре генов идёт независимо от других пар генов)
I закон Менделя (закон единообразия гибридов первого поколения или правило доминирования ) – при моногибридном скрещивании у гибридов первого поколения проявляются только доминантные признаки – оно фенотипически единообразно II закон Менделя (закон расщепления) – в потомстве, полученном от скрещивания гибридов первого поколения, наблюдается явление расщепления: четверть особей из гибридов второго поколения несёт рецессивный признак, три четверти – доминантный III закон Менделя (закон независимого расщепления или закон независимого комбинирования признаков) – при дигибридном скрещивании у гибридов каждая пара признаков наследуется независимо от других и даёт с ними разные сочетания. Образуются фенотипические группы, характеризующиеся отношением 9: 3: 3: 1 (расщепление по каждой паре генов идёт независимо от других пар генов)
 Анализирующее скрещивание Используется для определения генотипа организма, имеющего фенотипическое проявление доминантного признака. Для этого скрещивают чисто рецессивную особь с исследуемой, генотип которой необходимо установить, так как по фенотипу нельзя установить генотип у особей с доминантными признаками. Например, при скрещивании мышей с черной окраской шерсти(доминантный признак) и коричневой (рецессивный признак) получено потомство: 50% мышей имеют черную окраску шерсти и 50%- коричневую. Требуется установить генотип мышей с черной окраской шерсти. Так как в процессе скрещивания происходит расщепление, то, следовательно, мыши с черной окраской- гетерозиготны ( Аа). Признак Ген P: ♀ черная × ♂ коричневая черная А Аа аа коричневая а G: A a a F: Aa aa 50% черные 50% коричневые по фенотипу 1: 1 по генотипу 1: 1
Анализирующее скрещивание Используется для определения генотипа организма, имеющего фенотипическое проявление доминантного признака. Для этого скрещивают чисто рецессивную особь с исследуемой, генотип которой необходимо установить, так как по фенотипу нельзя установить генотип у особей с доминантными признаками. Например, при скрещивании мышей с черной окраской шерсти(доминантный признак) и коричневой (рецессивный признак) получено потомство: 50% мышей имеют черную окраску шерсти и 50%- коричневую. Требуется установить генотип мышей с черной окраской шерсти. Так как в процессе скрещивания происходит расщепление, то, следовательно, мыши с черной окраской- гетерозиготны ( Аа). Признак Ген P: ♀ черная × ♂ коричневая черная А Аа аа коричневая а G: A a a F: Aa aa 50% черные 50% коричневые по фенотипу 1: 1 по генотипу 1: 1
 Связь генотипа и фенотипа: варианты проявления менделевских закономерностей
Связь генотипа и фенотипа: варианты проявления менделевских закономерностей
 n Наследственность всегда реализуется во взаимодействии генетических факторов и условий существования. При индивидуальном развитии организмов (их онтогенезе) наследственность определяет границы (норму реакции) изменчивости организма, т. е. набор тех возможных вариантов (фенотипов), которые допускает данный генотип при изменениях среды (модификационная, онтогенетическая изменчивость). При историческом развитии организмов (их филогенезе) наследственность, закрепляя изменения генетического материала (генотипическая изменчивость), создаёт предпосылки для эволюции организмов.
n Наследственность всегда реализуется во взаимодействии генетических факторов и условий существования. При индивидуальном развитии организмов (их онтогенезе) наследственность определяет границы (норму реакции) изменчивости организма, т. е. набор тех возможных вариантов (фенотипов), которые допускает данный генотип при изменениях среды (модификационная, онтогенетическая изменчивость). При историческом развитии организмов (их филогенезе) наследственность, закрепляя изменения генетического материала (генотипическая изменчивость), создаёт предпосылки для эволюции организмов.
 Хромосомная и цитоплазматическая наследственность n n Первенствующая роль ядра и хромосом в Н. не исключает передачи некоторых признаков и через цитоплазму, в которой обнаружены структуры, способные к самовоспроизведению ( цитоплазматическая наследственность). Единицы цитоплазматической (нехромосомной) наследственности отличаются от хромосомных тем, что они не расходятся при мейозе. Поэтому потомство при нехромосомной наследственности воспроизводит признаки только одного из родителей (чаще матери). Т. о. , различают ядерную наследственность, связанную с передачей наследственных признаков, находящихся в хромосомах ядра (иногда её называют хромосомной), и вне ядерную, зависящую от передачи самовоспроизводящихся структур цитоплазмы. Ядерная наследственность реализуется и при вегетативном размножении.
Хромосомная и цитоплазматическая наследственность n n Первенствующая роль ядра и хромосом в Н. не исключает передачи некоторых признаков и через цитоплазму, в которой обнаружены структуры, способные к самовоспроизведению ( цитоплазматическая наследственность). Единицы цитоплазматической (нехромосомной) наследственности отличаются от хромосомных тем, что они не расходятся при мейозе. Поэтому потомство при нехромосомной наследственности воспроизводит признаки только одного из родителей (чаще матери). Т. о. , различают ядерную наследственность, связанную с передачей наследственных признаков, находящихся в хромосомах ядра (иногда её называют хромосомной), и вне ядерную, зависящую от передачи самовоспроизводящихся структур цитоплазмы. Ядерная наследственность реализуется и при вегетативном размножении.
 в хромосомах. Передача признаков") n n Хромосомная наследственность связана с распределением носителей наследственности (генов) в хромосомах. Передача признаков потомству особенно четко прослеживается при наследовании менделирующих признаков, т. е. таких наследственных признаков, которые в потомстве, расщепляются по моногенному типу наследования в соответствии с законами Менделя — эмпирическими правилами наследования, устанавливающими численные соотношения, в которых отдельные признаки и их сочетания проявляются в гибридном потомстве при половом размножении. Внехромосомная, или цитоплазматическая, наследственность заключается в наследовании признаков, которые контролируются факторами, локализованными у животных организмов в митохондриях, у растений — в митохондриях и пластидах, у бактерий — в плазмидах. Цитоплазматические элементы, обладающие свойством передачи наследственной информации, распределяются между дочерними клетками случайно, поэтому четкого менделевского расщепления в этих случаях не наблюдается. Все системы внехромосомной наследственности взаимодействуют с хромосомными генами или их продуктами.
n n Хромосомная наследственность связана с распределением носителей наследственности (генов) в хромосомах. Передача признаков потомству особенно четко прослеживается при наследовании менделирующих признаков, т. е. таких наследственных признаков, которые в потомстве, расщепляются по моногенному типу наследования в соответствии с законами Менделя — эмпирическими правилами наследования, устанавливающими численные соотношения, в которых отдельные признаки и их сочетания проявляются в гибридном потомстве при половом размножении. Внехромосомная, или цитоплазматическая, наследственность заключается в наследовании признаков, которые контролируются факторами, локализованными у животных организмов в митохондриях, у растений — в митохондриях и пластидах, у бактерий — в плазмидах. Цитоплазматические элементы, обладающие свойством передачи наследственной информации, распределяются между дочерними клетками случайно, поэтому четкого менделевского расщепления в этих случаях не наблюдается. Все системы внехромосомной наследственности взаимодействуют с хромосомными генами или их продуктами.
 Роль ядра в передаче наследственных признаков n Впервые мысль о роли ядра как носителя наследственности была сформулирована О. Гертвигом (1884) и Э. Страсбургером (1884) на основании изучения процесса оплодотворения. Т. Бовери (1887) установил индивидуальность хромосом и развил гипотезу о их качественном различии. Он же, а также Э. ван Бенеден (1883) установили уменьшение количества хромосом вдвое при образовании половых клеток в мейозе. Американский учёный У. Сеттон (1902) дал цитологическое объяснение закону Менделя о независимом наследовании признаков. Однако подлинное обоснование хромосомной теории Н. было дано в работах Т. Моргана и его школы (начиная с 1911), в которых было показано точное соответствие между генетическими и цитологическими данными.
Роль ядра в передаче наследственных признаков n Впервые мысль о роли ядра как носителя наследственности была сформулирована О. Гертвигом (1884) и Э. Страсбургером (1884) на основании изучения процесса оплодотворения. Т. Бовери (1887) установил индивидуальность хромосом и развил гипотезу о их качественном различии. Он же, а также Э. ван Бенеден (1883) установили уменьшение количества хромосом вдвое при образовании половых клеток в мейозе. Американский учёный У. Сеттон (1902) дал цитологическое объяснение закону Менделя о независимом наследовании признаков. Однако подлинное обоснование хромосомной теории Н. было дано в работах Т. Моргана и его школы (начиная с 1911), в которых было показано точное соответствие между генетическими и цитологическими данными.
 Экспериментальные доказательства роли ДНК в передаче наследственной информации
Экспериментальные доказательства роли ДНК в передаче наследственной информации
 Эксперимент Т. Бовери 1889 г. n В опытах по гибридизации двух видов морских ежей (Psammechinus и Sphaerichinus), имеющих морфологические различия, впервые было показано, что особи, развивающиеся после оплодотворения энуклеированных (безъядерных) яиц Sphaerichinus спермой ежа Psammechinus, развиваются в личинки с анатомическим строением исключительно отцовского организма (т. е. Psammechinus).
Эксперимент Т. Бовери 1889 г. n В опытах по гибридизации двух видов морских ежей (Psammechinus и Sphaerichinus), имеющих морфологические различия, впервые было показано, что особи, развивающиеся после оплодотворения энуклеированных (безъядерных) яиц Sphaerichinus спермой ежа Psammechinus, развиваются в личинки с анатомическим строением исключительно отцовского организма (т. е. Psammechinus).
 n Результат обмена ядрами между") Эксперимент Дж. Геммерлинга (30 -х гг. XX в. ) n Результат обмена ядрами между двумя видами ацетобулярии; шляпка удалена, ядра пересажены в энуклеированные ризоиды. Структуры, принадлежащие А. mediterranea , выделены серым цветом, а А. сrenulata – белым цветом
Эксперимент Дж. Геммерлинга (30 -х гг. XX в. ) n Результат обмена ядрами между двумя видами ацетобулярии; шляпка удалена, ядра пересажены в энуклеированные ризоиды. Структуры, принадлежащие А. mediterranea , выделены серым цветом, а А. сrenulata – белым цветом
 Эксперимент Астаурова Б. Л. по андрогенезу n Суть опытов заключалась в получении яиц тутового шелкопряда (Bombix mori) с инактивированными ионизирующим облучением ядрами и последующего их осеменения сперматозоидами. Использовали яйца самки, гусеницы которых были черными, а сперматозоиды - от самца с рыжими гусеницами.
Эксперимент Астаурова Б. Л. по андрогенезу n Суть опытов заключалась в получении яиц тутового шелкопряда (Bombix mori) с инактивированными ионизирующим облучением ядрами и последующего их осеменения сперматозоидами. Использовали яйца самки, гусеницы которых были черными, а сперматозоиды - от самца с рыжими гусеницами.
 и растений (табак, кукуруза) в") n n Андрогенез наблюдается у отдельных видов животных (шелкопряд) и растений (табак, кукуруза) в тех случаях, когда материнское ядро погибает до оплодотворения, которое при этом является ложным, то есть женское и мужское ядра не сливаются и в дроблении участвует только мужское ядро. Андрогенез — особый случай девственного развития, или партеногенеза; иногда его называют «мужской партеногенез» .
n n Андрогенез наблюдается у отдельных видов животных (шелкопряд) и растений (табак, кукуруза) в тех случаях, когда материнское ядро погибает до оплодотворения, которое при этом является ложным, то есть женское и мужское ядра не сливаются и в дроблении участвует только мужское ядро. Андрогенез — особый случай девственного развития, или партеногенеза; иногда его называют «мужской партеногенез» .
 получены только у тутового шелкопряда и наездника Habrobracon juglandis.") n Половозрелые животные (всегда самцы) получены только у тутового шелкопряда и наездника Habrobracon juglandis. При этом Б. Л. Астаурову и В. П. Остряковой удалось на животном впервые осуществить (1956) при скрещивании двух видов шелкопряда полный межвидовой андрогенез. Несколько случаев полного андрогенеза наблюдалось у растений при отдалённых скрещиваниях разных видов табака, скерды и кукурузы. Во всех случаях полного андрогенеза как растений, так и животных андрогенные потомки оказались сходными с отцовским видом, что указывает на ведущее значение клеточного ядра в наследственности.
n Половозрелые животные (всегда самцы) получены только у тутового шелкопряда и наездника Habrobracon juglandis. При этом Б. Л. Астаурову и В. П. Остряковой удалось на животном впервые осуществить (1956) при скрещивании двух видов шелкопряда полный межвидовой андрогенез. Несколько случаев полного андрогенеза наблюдалось у растений при отдалённых скрещиваниях разных видов табака, скерды и кукурузы. Во всех случаях полного андрогенеза как растений, так и животных андрогенные потомки оказались сходными с отцовским видом, что указывает на ведущее значение клеточного ядра в наследственности.
 Опыты Дж. Гердона с африканской шпорцевой лягушкой Xenopus laevis n Используя шпроцевую лягушку, ученый добился развития взрослой лягушки из энуклеированной яйцеклетки, в которую было пересажено ядро из эпителиальной клетки кожи лягушки или кишечника головастика, т. е. из соматической дифференцированной клетки.
Опыты Дж. Гердона с африканской шпорцевой лягушкой Xenopus laevis n Используя шпроцевую лягушку, ученый добился развития взрослой лягушки из энуклеированной яйцеклетки, в которую было пересажено ядро из эпителиальной клетки кожи лягушки или кишечника головастика, т. е. из соматической дифференцированной клетки.
 Опыты Гёрдона n 1 -2 % экспериментальных зародышей превращались во взрослых лягушек. Абсолютно точного копирования не было.
Опыты Гёрдона n 1 -2 % экспериментальных зародышей превращались во взрослых лягушек. Абсолютно точного копирования не было.
 ОПЫТ ГРИФФИТСА n Впервые на явление трансформации обратил внимание Ф. Гриффитс в 1928 г. , изучая пневмококки (Streptococcus pneumoniae), которые образуют капсулу в организме, а на агаре растут в виде гладких (S-форма) и блестящих колоний. При введении белым мышам убитых нагреванием вирулентных капсульных пневмококков ІІІ типа вместе с авирулентными бескапсульными пневмококками ІІ типа (R-форма) лабораторное животное через несколько дней погибало, а из ее крови высевались живые капсульные пневмококки ІІ серотипу. Следовательно, происходила глубокая перестройка генетического аппарата клетки, которая приобретала способность синтезировать капсулу.
ОПЫТ ГРИФФИТСА n Впервые на явление трансформации обратил внимание Ф. Гриффитс в 1928 г. , изучая пневмококки (Streptococcus pneumoniae), которые образуют капсулу в организме, а на агаре растут в виде гладких (S-форма) и блестящих колоний. При введении белым мышам убитых нагреванием вирулентных капсульных пневмококков ІІІ типа вместе с авирулентными бескапсульными пневмококками ІІ типа (R-форма) лабораторное животное через несколько дней погибало, а из ее крови высевались живые капсульные пневмококки ІІ серотипу. Следовательно, происходила глубокая перестройка генетического аппарата клетки, которая приобретала способность синтезировать капсулу.
 In vitro Эвери, Мак-Леод, Мак-Карти 1944") Трансформация у бактерий Pneumococcus (Гриффитс, 1928 г. ) In vitro Эвери, Мак-Леод, Мак-Карти 1944 ДНК-азная обработка
Трансформация у бактерий Pneumococcus (Гриффитс, 1928 г. ) In vitro Эвери, Мак-Леод, Мак-Карти 1944 ДНК-азная обработка
 штамма мыши погибают. Капсульные пневмококки размножаются.") Опыты Гриффита После заражения пневмококками капсульного патогенного (вирулентного) штамма мыши погибают. Капсульные пневмококки размножаются.
Опыты Гриффита После заражения пневмококками капсульного патогенного (вирулентного) штамма мыши погибают. Капсульные пневмококки размножаются.
 Опыты Гриффита При инфицировании пневмококками бескапсульного непатогенного штамма животные не погибают. Бескапсульные пневмококки не размножаются.
Опыты Гриффита При инфицировании пневмококками бескапсульного непатогенного штамма животные не погибают. Бескапсульные пневмококки не размножаются.
 штамма, предварительно убитыми нагреванием, мыши не") Опыты Гриффита После заражения пневмококками капсульного патогенного (вирулентного) штамма, предварительно убитыми нагреванием, мыши не погибают. Пневмококки не размножаются.
Опыты Гриффита После заражения пневмококками капсульного патогенного (вирулентного) штамма, предварительно убитыми нагреванием, мыши не погибают. Пневмококки не размножаются.
 ТРАНСФОРМАЦИЯ n В 1944 г. О. Евери, К. Маклеод и М. Маккарти смоделировали этот феномен in vitro, выделили и очистили трансформирующий агент. Им оказалась молекула ДНК. Трансформация происходит только в тех клетках, которые способны к ней. Такое состояние определяется понятием компетентности, природа которой окончательно не выяснена. Считается, что она предопределяется наличием особенного белка - компонента клеточной мембраны, способного расщеплять некоторые структурные элементы клеточной поверхности. Таким образом высвобождаются рецепторные участки, с которыми взаимодействует ДНК. Состояние компетентности формируется на определенных стадиях развития бактериальной клетки.
ТРАНСФОРМАЦИЯ n В 1944 г. О. Евери, К. Маклеод и М. Маккарти смоделировали этот феномен in vitro, выделили и очистили трансформирующий агент. Им оказалась молекула ДНК. Трансформация происходит только в тех клетках, которые способны к ней. Такое состояние определяется понятием компетентности, природа которой окончательно не выяснена. Считается, что она предопределяется наличием особенного белка - компонента клеточной мембраны, способного расщеплять некоторые структурные элементы клеточной поверхности. Таким образом высвобождаются рецепторные участки, с которыми взаимодействует ДНК. Состояние компетентности формируется на определенных стадиях развития бактериальной клетки.
 МЕХАНИЗМ ТРАНСФОРМАЦИИ n Механизм явления трансформации заключается в том, что сначала на поверхности реципиента клетки адсорбируется небольшой фрагмент двониткової ДНК (1/250 -1/500 часть хромосомы донора клетки). Впоследствии он проникает внутрь клетки, где одна нить ДНК переваривается эндонуклеазами, а другая - монтируется в клеточную хромосому. Наступает последняя фаза процесса - экспрессия рекомбинантов. Такой процесс интеграции происходит очень быстро. Исследовано, что для появления рекомбинантов достаточно пятидесятиминутного контакта реципиента клетки с донорской ДНК, а сам процесс завершается через 2 часа.
МЕХАНИЗМ ТРАНСФОРМАЦИИ n Механизм явления трансформации заключается в том, что сначала на поверхности реципиента клетки адсорбируется небольшой фрагмент двониткової ДНК (1/250 -1/500 часть хромосомы донора клетки). Впоследствии он проникает внутрь клетки, где одна нить ДНК переваривается эндонуклеазами, а другая - монтируется в клеточную хромосому. Наступает последняя фаза процесса - экспрессия рекомбинантов. Такой процесс интеграции происходит очень быстро. Исследовано, что для появления рекомбинантов достаточно пятидесятиминутного контакта реципиента клетки с донорской ДНК, а сам процесс завершается через 2 часа.
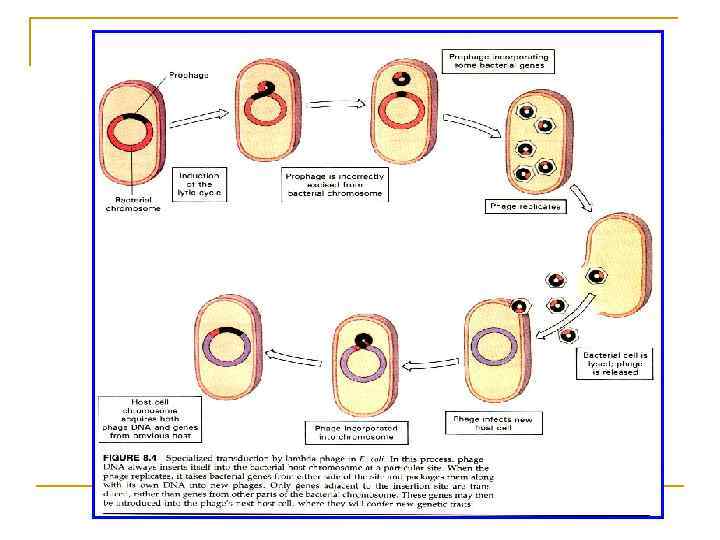
 ТРАНСДУКЦИЯ n В 1952 г. Н. Циндер и Д. Ледерберг, изучая процесс конъюгации между разными штаммами сальмонел, обратили внимание, что иногда обмен генетическим материалом происходит не в результате конъюгации, а в результате высвобождения из родительских штаммов умеренного бактериофага. Явление обмена генетической информацией у бактерий путем переноса фагами фрагментов ДНК от донора клетки к реципиенту клетки получило название трансдукції. Она часто происходит в энтеробактерий, псевдомонадах, стафилококках и бациллах. Считают, что большинство видов микроорганизмов несут в своем геноме профаг, потому трансдукция может быть чрезвычайно распространенным явлением в микробном мире.
ТРАНСДУКЦИЯ n В 1952 г. Н. Циндер и Д. Ледерберг, изучая процесс конъюгации между разными штаммами сальмонел, обратили внимание, что иногда обмен генетическим материалом происходит не в результате конъюгации, а в результате высвобождения из родительских штаммов умеренного бактериофага. Явление обмена генетической информацией у бактерий путем переноса фагами фрагментов ДНК от донора клетки к реципиенту клетки получило название трансдукції. Она часто происходит в энтеробактерий, псевдомонадах, стафилококках и бациллах. Считают, что большинство видов микроорганизмов несут в своем геноме профаг, потому трансдукция может быть чрезвычайно распространенным явлением в микробном мире.
 и Альфред") Эксперимент Альфреда Херши и Марты Чейз. 1952 г. Марта Чейз (1927– 2003) и Альфред Херши (1908– 1997)
Эксперимент Альфреда Херши и Марты Чейз. 1952 г. Марта Чейз (1927– 2003) и Альфред Херши (1908– 1997)
") Бактериофаг T 2 – один из наиболее изученных фагов кишечной палочки (Escherichia coli)
Бактериофаг T 2 – один из наиболее изученных фагов кишечной палочки (Escherichia coli)
 Эксперимент Альфреда Херши и Марты Чейз. 1952 г. Суть опыта: Фаги, у которых белковая оболочка была мечена радиоактивной серой (S 35), а ДНК - радиоактивным фосфором (Р 32), инкубировали с бактериями. Затем бактерии отмывали. В смывных водах не обнаруживали Р 32, а в бактериях - S 35 Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих и белковую оболочку, и ДНК.
Эксперимент Альфреда Херши и Марты Чейз. 1952 г. Суть опыта: Фаги, у которых белковая оболочка была мечена радиоактивной серой (S 35), а ДНК - радиоактивным фосфором (Р 32), инкубировали с бактериями. Затем бактерии отмывали. В смывных водах не обнаруживали Р 32, а в бактериях - S 35 Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих и белковую оболочку, и ДНК.
 Эксперимент Альфреда Херши и Марты Чейз. Фаги, у которых белковая оболочка была мечена радиоактивной серой (S 35), а ДНК радиоактивным фосфором (Р 32), инкубировали с бактериями.
Эксперимент Альфреда Херши и Марты Чейз. Фаги, у которых белковая оболочка была мечена радиоактивной серой (S 35), а ДНК радиоактивным фосфором (Р 32), инкубировали с бактериями.
 Эксперимент Альфреда Херши и Марты Чейз. Бактерии отмывали. В смывных водах не обнаруживали Р 32, а в бактериях - S 35. В то же время большая часть Р 32 оказалась внутри бактериальных клеток, а вся S 35 - в окружающей среде.
Эксперимент Альфреда Херши и Марты Чейз. Бактерии отмывали. В смывных водах не обнаруживали Р 32, а в бактериях - S 35. В то же время большая часть Р 32 оказалась внутри бактериальных клеток, а вся S 35 - в окружающей среде.
 Эксперимент Альфреда Херши и Марты Чейз. Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих немеченую белковую оболочку, и ДНК, меченую Р 32.
Эксперимент Альфреда Херши и Марты Чейз. Следовательно, внутрь попала только ДНК. Через несколько минут из бактерии выходили десятки полноценных фагов, содержащих немеченую белковую оболочку, и ДНК, меченую Р 32.
 Эксперимент Альфреда Херши и Марты Чейз. Отсюда следовал однозначный вывод о том, что именно ДНК выполняет генетическую функцию - несет информацию, как о создании новых копий ДНК, так и о синтезе фаговых белков.
Эксперимент Альфреда Херши и Марты Чейз. Отсюда следовал однозначный вывод о том, что именно ДНК выполняет генетическую функцию - несет информацию, как о создании новых копий ДНК, так и о синтезе фаговых белков.
 Опыты Френкеля-Конрада 1957 г. ФРЭНКЕЛЬ-КОНРАД Хейнц 1910 - 1999
Опыты Френкеля-Конрада 1957 г. ФРЭНКЕЛЬ-КОНРАД Хейнц 1910 - 1999
. ВТМ") Опыты Френкеля-Конрада 1957 г. Суть опыта: Френкель-Конрат работал с вирусом табачной мозаики (ВТМ). ВТМ В вирусе ВТМ содержится РНК, а не ДНК. Разные штаммы вируса вызывают разную картину поражения листьев табака. После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком.
Опыты Френкеля-Конрада 1957 г. Суть опыта: Френкель-Конрат работал с вирусом табачной мозаики (ВТМ). ВТМ В вирусе ВТМ содержится РНК, а не ДНК. Разные штаммы вируса вызывают разную картину поражения листьев табака. После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком.
 Опыты Френкеля-Конрада РНК покрыта белковой облочкой. Вирус можно "разобрать" и "собрать" снова, но уже так, что РНК вируса одного штамма будет окружена белковой оболочкой вируса другого штамма.
Опыты Френкеля-Конрада РНК покрыта белковой облочкой. Вирус можно "разобрать" и "собрать" снова, но уже так, что РНК вируса одного штамма будет окружена белковой оболочкой вируса другого штамма.
 Опыты Френкеля-Конрада После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком (на рисунке - штамм 1).
Опыты Френкеля-Конрада После смены белковой оболочки "переодетые" вирусы вызывали картину поражения, характерную для того штамма, чья РНК была покрыта чужим белком (на рисунке - штамм 1).
 Опыты Френкеля-Конрада Не только ДНК, но и РНК может служить носителем генетической информации.
Опыты Френкеля-Конрада Не только ДНК, но и РНК может служить носителем генетической информации.