Membrane_2010.ppt
- Количество слайдов: 46



Общая физиология возбудимых тканей 1. Состав и свойства плазматической мембраны 2. Распределение ионов во внутри- и внеклеточных средах 3. Транспорт веществ через мембрану 4. Na/K АТФаза и вторично-активный транспорт 5. Ионные каналы 6. Потенциал покоя и равновесные потенциалы 7. Потенциал действия 8. Аккомодация 9. Следовые потенциалы 10. Кривая сила-длительность

- автономность по отношению к окружающей среде - связь с окружающей средой КЛЕТКА МЕМБРАНА Основные функции: 1. барьерная, 2. механическая, 3. матричная (распределение белков). Состав мембраны по сухой массе: - 20 -70% липиды (в среднем 30%), -20 -65% белки (в среднем 60%) (20% в миелине, а 80% в митохондриях), -10% углеводы (в составе гликолипидов и гликопротеинов).
 мембрана ЖИДКО-МОЗАИЧНАЯ МОДЕЛЬ (1972 г. ) ЖИДКО(i)-КРИСТАЛЛИЧЕСКОЕ (ii) СОСТОЯНИЕ: I) липиды в")
Плазматическая (поверхностная) мембрана ЖИДКО-МОЗАИЧНАЯ МОДЕЛЬ (1972 г. ) ЖИДКО(i)-КРИСТАЛЛИЧЕСКОЕ (ii) СОСТОЯНИЕ: I) липиды в жидком состоянии: латеральная диффузия - 5 мкм/с за 1 сек десятки млн. перестановок (t оседлости = 10 -7/-8 с) 10 -9 м = 10 нм II) В расположении липидов «дальний» порядок: -хвосты липидов ~ параллельны - головки однообразно ориентированы Белки: - л. диффузия – 0, 2 мкм/с, хотя некоторые белки заякорены - на 1 белок (протеин) приходится 50 молекул липидов
 глицерол Диацилглицерол")
Основные липиды мембраны: гликосфинголипиды глицеролипиды сфинголипиды стерины фосфатидная Кислота (0. 5 %) глицерол Диацилглицерол (ДАГ) С 16 - С 18 жирные кислоты ¾ ненасыщенные в кристалл сост. при низких Т 0 С холестерин (~5 р меньше, чем ФЛ) СФИНГОЗИН (С 18) R: 1. Фосфатидилхолин (30 %) С 16 -С 18 2. Фосфатидилэтаноламин (20%) насыщенная жирная кислота 3. Фосфатидилсерин (10%) 4. Фосфатидилинозитол (4%) ЦЕРАМИД R: Фосфохолин (13%) фосфоэтаноламин R: глюкоза галактоза олигосахариды
 Фосфатидилхолин (10 часов) Сфингимиелин (5 часов) Холестерин")
Распределение липидов в плазматической мембране Гликолипиды (----) Фосфатидилхолин (10 часов) Сфингимиелин (5 часов) Холестерин (2 мин) Фосфатидилэтаноламин Фосфатидилсерин пассивно: «флип-флоп» (в среднем 1 час) + Энерго-затратно: АТФ-транслоказы фосфолипидов
 более")
ФАЗОВЫЕ МЕМБРАННЫЕ ПЕРЕХОДЫ крио-повреждение Повышается проницаемость для ионов сквозные отверстия (1 -3 нм) более жидкая гель T 0 C или Са 2+ жидкий кристалл Выход воды и солей более плотная + холестерин если холестерина больше 50% фазовых переходов нет T 0 C ф. п. изменяется -20 до +60 в зависимости от количества ненасыщенных жирных к-т. T 1 При старении вязкость увеличивается T 1 > T 2 В клетках дистальной части ноги оленя много ненасыщенных липидов (T 0 C ф. п. ~ -20 С )

- локализация ферментов, - условия для сопряжения катализа и трансмембранного переноса, - доступность водонерастворимых субстратов, - оптимальное микроокружение
 1997, Кай Зимонс холестерол гликолипиды Перестройка липидных плотиков 1 2 DIC")
Липидные плотики (рафты) 1997, Кай Зимонс холестерол гликолипиды Перестройка липидных плотиков 1 2 DIC 18 твердая фаза BODY PC жидкая фаза 1+2 5 мкм Вирусы СПИДа, кори и гриппа, лихорадки Эбола – используют липидные плотики для проникновения

Ассиметричное распределение ионов во вне и внутриклеточных средах Na – более гидрофильный, а К – более гидрофобный Na. Cl ингибитор клеточных процессов -12 раз KA активатор клеточных процессов 30 раз 155 m. M органические - анионы (А ) -14 раз -25000 раз p. H 7. 4 р. Н 7. 2 8 m. M -3 раза НСО- 27 m. M 3 половина объема клетки – цитоплазма, в цитоплазме по массе 20 % белка состояние геля (цитозоль)
 A/d")
Природа ионной проницаемости мембран ДИФФУЗИЯ: Поток вещества = D (C 1 -C 2) A/d = P (C 1 - C 2) A D- коэффициент диффузии D /d = P d – «толщина мембраны» Р - проницаемость А – площадь обмена С 1 -С 2 – градиент концентраций D=URT U- подвижность частиц Т- температура С 1 d С 2 A диполь ПРОНИЦАЕМА Н 2 О -газы (O 2, CO 2, NO, CO, H 2 S) -жиры, -полярные молекулы малого размера (этанол, мочевина) H+ + HCOO- = HCOOH -ионы большого радиуса, т. к. малая плотность заряда (йод, мышьяк-) * Неполярные вещества (гексан) могут «застревать» в бислое НЕПРОНИЦАЕМА заряженные молекулы

Проницаемость для воды -движение ОСМОС воды через пассивный транспорт полупроницаемую мембрану из области низкой концентрации растворенного вещества в область высокой. полости (кинки) [сахар] Тепловое движение (вращение) -С-С- [сахар] за 1 мкс [сахар] растворенное вещество вода мембрана, непроницаемая для сахара
 d = 2. 8 Ао соответствует размеру")
Селективность: Каналы для воды - аквапорины (1) d = 2. 8 Ао соответствует размеру молекулы H 2 O (2) заряд Arg + 195 предотвращает проникновение + (1) (2) (3) в середине канала два + заряда разрушают водородные связи (3) первый AQ идентифицирован в 1992 г.
,")
1 1 1 Аквапорин – состоит из 4 -х субъединиц ( « 1+1+1+1» ), каждая из которых формирует водную пору. 1987 г. Питер Эгр (Нобелевская премия по химии, 2003)
 изменяется в ходе транспорта пассивные переносчики: ПО ГРАДИЕНТУ")
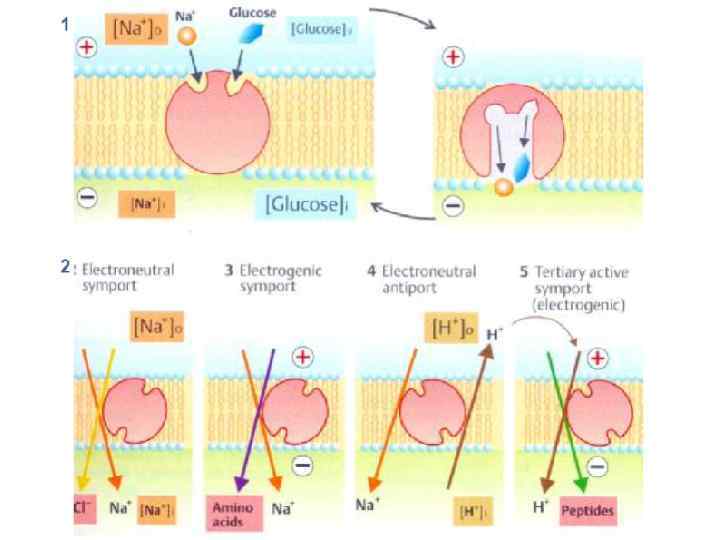
Переносчики – ТРАНСПОРТНЫЕ БЕЛКИ: конформация (строение) изменяется в ходе транспорта пассивные переносчики: ПО ГРАДИЕНТУ [С] (унипортеры) «активные» переносчики: ПРОТИВ ГРАДИЕНТА [C] I) первично-активные 1. Сопряженные с Red-Ox реакциями 2. Зависимые от поглощения света 3. АТФазы число оборотов: от 102 до 103 с-1 воротный, вращательный, эстафетный , челночный тип V облегченная диффузия II) вторично-активные 1. Симпортеры 2. Антипортеры Ферментативная реакция Vmax НАСЫЩЕНИЕ все переносчики задействованы АТФаза ½ Vmax Km число оборотов: от 102 до 105 с-1 C 1 -C 2 симпорт

Первично-активный транспорт: Са 2+ -АТФаза, Н+/K+-помпа, Na+/ K+ АТФаза 1957 г. Дж. Скоу выделил фермент из нерва краба (1997 г. Ноб. премия) β-субъединица -около 10% всех белков клетки -расходует 1/3 всей энергии клетки (иногда до 70%) места соединения с ингибитором оуабаином -3 формы α-субъединиц (8 -12 S) -3 формы β-субъединиц Na+/ K+ АТФаза α-субъединица снижает активность на 50% 10 -7 M Оуабаин (строфантин G) центр связывания ионов сайт связывания АТФ (сердечные гликозиды, препараты наперстянки)
![Блокада Na/K АТФазы Увеличение [Na]i Приток в клетку Cl- и воды Набухание клетки Конформационные](https://present5.com/presentation/21041214_179892527/image-16.jpg "Блокада Na/K АТФазы Увеличение [Na]i Приток в клетку Cl- и воды Набухание клетки Конформационные")
Блокада Na/K АТФазы Увеличение [Na]i Приток в клетку Cl- и воды Набухание клетки Конформационные состояния: Е 1 и Е 2 10 милисекунд на 1 цикл 10 m. V оуабаин МП на 10 м. В +АТФ - В клетках надпочечников синтезируется эндогенное оуабаин подобное соединение (особенно в ответ на АКТГ) - регуляторная γ- субъединица Na/K АТФазы снижает сродство фермента к Na и K Электрогенный (150 -600 зарядов в секунду)

Са 2+-насос 10 трансмембранных спиралей АТФ Са 2+ связывающий Са 2+-сайт P 3 - привод Mg нуклеотидсвязывающий домен Активаторы полиненасыщенные жирные кислоты, кислые фосфолипиды перенос Са 2+ сопровождается антипортом протонов H+
 Котранспорт: Na+ и аминокислот, Na+")
Вторично-активный транспорт Na+ Na+ пассивный унипорт (один вид молекул) Котранспорт: Na+ и аминокислот, Na+ и глюкозы, Na+ и остатка фосфорной кислоты Na+, К+, и Сl Na+ и Cl. Na+ и НСО 3 Н+ и олигопептидов Н+ и иона двухвалентного металла Н+ и RCOOK+ и Сl- Обменники: Na+ / H+ антипорт (защелачивает цитозоль), 3 Na+ /Ca 2+ - в 50 раз эффективнее удаляет избыток Са 2+ чем Са 2+АТФаза (т. к. его концентрация намного больше) Na+ / Сl 2 HCO 3 - и Na+ / Сl. Cl- / HCO 3 SO 42 - / анион-

ТРАНСПОРТНЫЕ БЕЛКИ: Каналы -изменения структуры касается только открытия / закрытия канала закрыт открыт пора 106 -8 ионов в 1 сек V Скорость транспорта Проводимость канала зависит от : Диффузия через канал - проницаемости (свойство канала), -концентрации ионов около устья канала. разность концентраций C 1 -C 2
 Хотя есть и неселективные каналы заряд иона размер")
Ионные каналы, свойства - избирательность (селективность) Хотя есть и неселективные каналы заряд иона размер иона (молекулярное сито) -селективный фильтр (3 А для K+) спонтанно периодически изменяются фиксированные заряды гидратные (водные) оболочки пора проницаема для иона, если ее диаметр точно соответствует размеру одной из гидратных оболочек
 конформационные перестройки Открытое / закрытое состояния канала")
Ионные каналы, свойства -воротный механизм (способность открываться/закрываться) конформационные перестройки Открытое / закрытое состояния канала модель «шара на цепочке» Открытое / закрытое состояния канала Среднее время открытия – величина постоянная для данного вида канала (обычно – 1 мс)

Ионные каналы, свойства Каналы - покоя Воротные каналы -управление воротами Активация канала – это увеличение вероятности открытия канала, собственный сенсор а не увеличение времени открытого состояния канала потенциал–чувствительные хемо(лиганд)–зависимые механо-чувствительные сенсор разобщен с каналом рецептор-управляемые ионные каналы стимул инактивация десенситизация Физические изменения в мембране активируют канал: - потенциал-активируемые каналы - механочувствительные каналы Химические вещества активируют канал (лиганд-активируемые): - ионотропные рецепторы (отвечают на внеклеточные стимулы), - каналы, активируемые с внутриклеточной стороны

МЕХАНОЧУВСт. ВИТЕЛЬНОСТЬ Стретч – активируемые ионные каналы + некоторые селективные каналы имеют свойства стретч-каналов обычно неселективные: пропускают Na+, K+, Ca 2+ Механо – активируемые ионные каналы Рецептор –управляемые механо-чувствительные каналы

Потенциал -чувствительный канал Родоначальники K-каналы, путем удвоения генов из них образовались Na+ и Ca 2+ Потенциал (напряжение) - электрическая работа, необходимая для перемещения заряда (без трения) из одной точки пространства в другую U (напряжение), Вольт q (заряд), Кл А (работа), Дж
 канал")
1 семейство, гены 2, 3, 17 хромосом, 9 изоформ. Потенциал управляемый натриевый (Na+) канал модулирует активацию / инактивацию 36 к. Да связывается с внеклеточным матриксом 260 к. Да 33 к. Да выстилка поры α-субъединица: 4 домена, каждый из 6 сегментов модуль, контролирующий открытие/закрытие в S 4 много +заряженных а/к (Lys и Arg) двое ворот Р –сегменты, окружают пору
 К-каналы задержанного выпрямления (фаза реполяризации в ПД,")
Калиевые каналы с 6 -трансмембранными сегментами: 1) К-каналы задержанного выпрямления (фаза реполяризации в ПД, инактивируются очень медленно), 2) быстро инактивирующиеся А-каналы (генерация ритмической активности, латентный период), 3) медленно активирующиеся К каналы и 4) Са активируемые К каналы (следовая гиперполяризация), 5) Shaker каналы (трепетание при эфирной анестезии). быстрые медленные (десятки милисек) +S 0 (реполяризация) Х модулирует воротный механизм Инактивационные N -ворота, открывающиеся при деполяризации 4 гомо- и гетеротетрамерные ионы «проталкивают» друга - O- O O из 20 а/к на N-конце «решетка» из карбоновых групп долгая деполяризация
 инактивационные ворота типа «шар на")
дегидратация иона К+ селективный фильтр активационные ворота (конформационный тип) инактивационные ворота типа «шар на цепочке» ионы К + подталкивают друга Na+ H 20
 открываются при снижении")
Калиевые каналы с 2 -трансмембранными сегментами: 1. АТФ -чувствительные К-каналы (KATP) открываются при снижении концентрации АТФ в клетке 2. K-каналы внутреннего выпрямления (KIR), движение К внутрь клетки (когда мембранный потенциал становится отрицательнее чем калиевый равновесный потенциал блокируютcя полиаминами /спермин, спермидин/, Mg 2+, Cs, Ba, ТЭА) 3. Управляемые G-белками К каналы (KGIR), рецептор -управляемые каналы (отрываются в ответ на связывание с Gβγ – комплексом ) Калиевые каналы с 4 -трансмембранными сегментами: К-каналы «утечки» - при МПП открыты Х 4 нет S 4 сенсора напряжения 2 поры Х 2 аритмия, глухота, эпилепсия, диабет, гипертония, гипотония… менее выраженные свойства внутреннего выпрямления у человека обнаружено более 100 генов, кодирующих калиевые каналы в каждой ткани набор К-каналов сильно гетерогенен
 каналы Лиганд –зависимые (Ry. R, IP 3")
Депо -управляемые Критерии классификации: Кальциевые (Са 2+) каналы Лиганд –зависимые (Ry. R, IP 3 R, NDMA-R ) Потенциал -управляемые - величина сдвига мембранного потенциала необходимая для активации низко-(high-voltage activated)пороговые высоко-(low voltage activated)пороговые - чувствительность к фармакологическим агентам L (long lasting)-тип дигидропиридины R(residual)-тип [никель] Т(transient)-тип -структурная гомология 3 семейства либефразил N (neuronal)-тип 70% сходства P(Purkinje)/Q-тип яд воронковых пауков только в скелетных мышцах сходна с α-субъединицей Na-канала 4 домена из 6 сегментов, 10 генов отвечает за инактивацию 4 гена
")
Хлорные каналы Лиганд-управляемые потенциал –управляемые, Сl. C-тип - 9 видов, двупоровые димеры (ассиметричные поры) Са 2+ -активируемые Сl-каналы (регуляция объема клетки) Макси(310 п. С) Мини(42 -69 п. С) СFTR (cystic fibrosis transmembrane conductance regulator) Ионотропные рецепторы (ГАМКА, ГАМКС, Гли) 2 типа ворот (Сl. C-0) медленные (секунды) Сl- открывают обе поры быстрые (милисек) независимые в каждой поре, способствуют закрыванию при гиперполяризации Сl. C-0, Cl. C-1 (в скелетных мышцах) – открыты при МП ~ 0 м. В, закрываются в ответ на гиперполяризацию Сl. С-2 (во всех тканях), открываются в ответ на гиперполяризацию, реагирует на разбухание клеток Сl. C-0, Cl. C-1, Сl. C-0, Cl. C-4, Cl. C-5 (в почках) чувствительны к р. Н среды Сl. C-3 (в синаптических везикулах) миотония, почечная патология (болезнь Dent’s), дегенерация сетчатки, остеопороз, Bartter's синдром
![Калиевый равновесный потенциал (E K+) внеклеточная среда [Na+] = [Cl-] 1. Вне: [Na+]=[Cl-] Внутри:](https://present5.com/presentation/21041214_179892527/image-32.jpg "Калиевый равновесный потенциал (E K+) внеклеточная среда [Na+] = [Cl-] 1. Вне: [Na+]=[Cl-] Внутри:")
Калиевый равновесный потенциал (E K+) внеклеточная среда [Na+] = [Cl-] 1. Вне: [Na+]=[Cl-] Внутри: [K+]=[A-] 2. Высокая проницаемость для К+, [K+]внутри>>> [K+]вне Мембрана А- АК+ К+ К+ А- 0. 1 -1% каналов открыто К+ АК+ 3. Выход K+ из клетки К+ градиент концентрации К+ К+ К+ А- А- АК+ А- К+ Аэлектрический градиент Аосм = R T ln[K+]0/[K+]i концентрации Ф. Нернста Аосм = А эл электрический градиент 4. Формируется разность потенциалов, способствующая обратному потоку К+ Аэл = n F Eion 5. Ситуация, когда выход К+ уравновешен входом К+, обозначается равновесный потенциал для ионов К+ R- газовая постоянная (кинетическая энергия 1 моля ионов при Т=1 К) Т- температура в Кельвинах n – валентность ионы F- число Фарадея (заряд 1 моля одновалентных ионов) Е – потенциал

Cl. Na+ Cl- Cl- Na+ Na+ Na+ Cl- Электрогенный эффект Na/K насоса Na+ от - 1 до - 10 м. В в зависимости от активности 0 м. В градиент концентрации Cl. Cl- Na+ АТФ Cl. Na+ электрический градиент Na+ Na+ Cl. Cl- Cl- Na+ Na+ К+ К+ E Na + 55 -70 м. В (в нейронах) / + 70 -80 м. В (в скелетных мышцах) Na+ Na+ на 20 -30 Na+ каналов : 1000 К+ каналов E к = - 75 / -95 м. В E Cl = - 80 -90 м. В в покое: Рк : PNa : PCl = 1 : 0, 04 : 0, 45
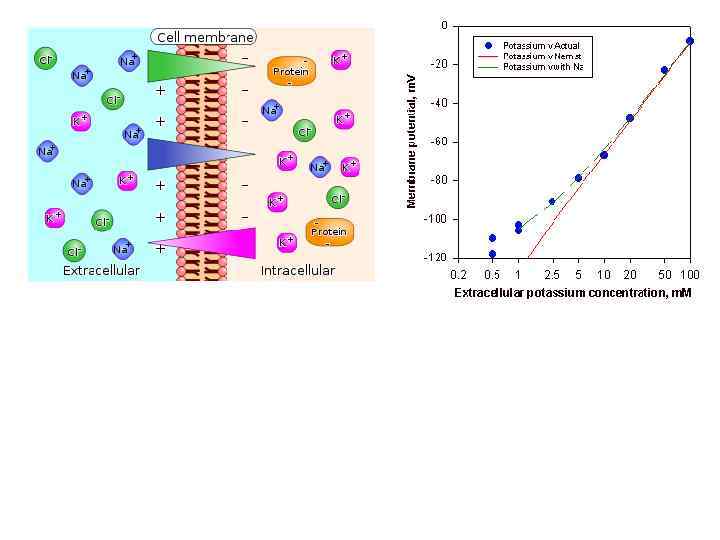

Потенциал покоя К+проводимость > Сl- проводимость > вклад Na/K насоса > Na проводимость > другие системы транспорта Нейроны – 40 – 70 м. В, ППМ – 90 м. В, ГМК – 30 м. В. деполяризация выходящий ток - входящий в клетку ток E Na = +55 м. В - 60 м. В - 50 м. В де 0 м. В -60 м. В гиперполяризация - выходящий из клетки ток гип -75 м. В E к = - 75 м. В ер- - 60 м. В - 70 м. В -75 м. В
 – возбудимые Нейроны Мышечные")
клетки способные к возбуждению ( то есть генерации электрического ответа) – возбудимые Нейроны Мышечные клетки некоторые Железистые клетки Потенциал – чувствительный (управляемый) Na-канал потенциал покоя деполяризация ТТХ деполяризация 2 активационные ворота, m (быстрые) инактивационные ворота, h (медленные) 3 t = 0, 7 мс деполяризация усиливается На 1 мкм 2 – 1 до 50 Na каналов, за 1 мс через канал переносятся 6000 ионов Na+ изменения мембранного потенциала на 100 м. В Блокаторы: 1. тетродотоксин (из рыбы фуга) cакситоксин (цианобактерии) Активаторы: 2. вератридин, грайанотоксин (полипептиды из растений) батрахотоксин (кожный секрет колумбийских лягушек) 3. Анемонотоксины I, III (морская змея анемона) α-токсины скорпиона

Мембрана деполяризована деполяризация покой Мембрана деполяризована реполяризация Мембрана в процессе реполяризации

Сl 1 - 3 2 Na+ Na+ цитоплазма A цитоплазма - 4 5 6 Na+ K+ 3 х 7 K+ К- каналы задержанного выпрямления(Kdr) Na+ АТФ цитоплазма 2 х K+ Блокаторы: - тетраэтиламмоний, - 4 -аминопиридин во время ПД 1/10 млн. часть K+ покидает клетку
 Фазы ПД: 1 – быстрая деполяризация (0. 5 -1 мс) 2")
Потенциал действия (спайк) Фазы ПД: 1 – быстрая деполяризация (0. 5 -1 мс) 2 - реполяризация (I) (III) овершут (+30 -50 м. В) К-каналы Если dr-типа, утечки – то медленно Если А-типа –то быстро 1 (II) 2 порог (I) Na-каналы (в некоторых клетках Са) (III) (II) Интенсивность стимула (I) потенциал действия Рк : PNa : PCl = 1 : 20 : 0, 45 покой Рк : PNa : PCl = 1 : 0, 04 : 0, 45

Элетротонический потенциал - сдвиг мембранного потенциала клетки, вызываемый действием постоянного электрического тока. Пассивная реакция клетки на электрический раздражитель; состояние ионных каналов и транспорт ионов при этом не изменяется. Локальный ответ – активная реакция клетки на электрический раздражитель, однако состояние ионных каналов и транспорт ионов при этом изменяется незначительно. Его называют местным возбуждением, так как это возбуждение не распространяется по мембранам возбудимых клеток. Потенциал действия – возбуждение клетки, проявляющееся в форме скоротечного изменения мембранного потенциала, которое распространяется по мембране. Лишь небольшое количество ионов проходит через мембрану во время потенциала действия, и концентрация ионов Na+ и K+ практически не изменяется.
 (g. Ca 2+ ) открывается несколько раз в течение 5")
Рефрактерность (периоды «молчания» ) (g. Ca 2+ ) открывается несколько раз в течение 5 мс й но ль те инактивация Na+ каналов си но от ой тн лю со аб за время деполяризации открывается 1 раз на 0. 7 мс периоды рефрактерности: (невосприимчивости) Проводимость – определяется количеством открытых каналов высокая g. K+

1963 г. Нобелевская премия по физиологии и медицине, совместно с Джоном С. Эклсом и Аланом Ходжкином
,")
Феномен аккомодации медленно быстро очень медленно постепенная инактивация Nа каналов!!! чем меньше порог (Ек), тем больше возбудимость увеличение К проницаемости Если у клеток мембранный потенциал менее электроотрицательный, чем – 40 м. В, то свойство возбудимости утрачивается у двигательных нервных волокон скорость аккомодации, как правило, значительно выше, чем у чувствительных волокон.
 2) следовая гиперполяризация")
Следовые потенциалы реполяризация быстрая деполяризация следовая деполяризация – замедление реполяризации 1) 2) следовая гиперполяризация Еm E k+ 1) – снижение проницаемости для ионов К + через К-каналы задержанного выпрямления и быстрые Са-активируемые К-каналы (блоратор: харибдотоксин скорпиона) 2) – медленно активирующиеся К-каналы, Ks-каналы (4 -аминопиридин) - медленные Са 2+ активируемые К-каналы Na+ -(ибериотоксин, апамин, бревитоксин-В) - возросшая активность Na/K АТФазы Cl 3 Na+ А- K+ 2 K+

Зависимость между временем действия раздражителя и его силой -для функциональной оценки нервно-мышечной системы человека минимальная величина силы раздражителя – реобаза (абсолютный порог силы) минимальное время в течение которого должен действовать раздражитель пороговой силы – полезное время высокочастотный ток (10 к. Гц) прогревание Хронаксия – время в течение которого должен действовать стимул удвоенной реобазы чем меньше хронаксия, тем больше возбудимость гипербола абсолютный порог времени

Membrane_2010.ppt