МОРФОЛОГИЯ 3.ppt
- Количество слайдов: 47



Общая цитология и цитофизиология немембранных клеточных органелл

К мембранным органеллам относятся 1. митохондрии, 2. эндоплазматическая сеть, 3. аппарат Гольджи, 4. лизосомы. • • • Немембранные органеллы: свободные рибосомы и полисомы, микротрубочки, центриоли филаменты (микрофиламенты, промежуточные филаменты).

Рибосомы Элементарные аппараты синтеза белковых, полипептидных молекул — обнаруживаются во всех клетках. Рибосомы — это сложные рибонуклеопротеиды, в состав которых входят белки и молекулы РНК примерно в равных весовых отношениях. Размер функционирующей рибосомы эукариотических клеток 25 x 20 нм. Такая рибосома состоит из большой и малой субъединиц. Каждая из субъединиц построена из рибонуклеопротеидного тяжа, где р. РНК взаимодействует с разными белками и образует тело рибосомы.
. Рибосомы могут располагаться свободно в")
• Различают единичные рибосомы и комплексные рибосомы (полисомы). Рибосомы могут располагаться свободно в гиалоплазме или быть связанными с мембранами эндоплазматической сети. В малоспециализированных и быстрорастущих клетках в основном обнаруживаются свободные рибосомы. В специализированных клетках рибосомы располагаются в составе гранулярной эндоплазматической сети. Степень интенсивности синтетической деятельности свободных рибосом меньше, а образуемые белки используются в основном на собственные нужды клетки. Связанные рибосомы обеспечивают синтез белков «на экспорт» , т. е. на обеспечение нужд организма. Строение эукариотической клетки: 1 -плазмолема; 2 -гиалоплазма; 3 кариолема; 4 -хроматин; 5 -кариоплазма; 6 ядрышко; 7 -ядерные поры; 8 -агранулярная эндоплапзматическая сеть; 9 -гранулярная эндоплапзматическая сеть; 10 -рибосома; 11 - комплекс Гольджи; 12 -митохондрия; 13 вакуоль; 14 -лизосомы; 15 -пероксисомы; 16 клеточный центр (две центриоли); 17 микротрубочка; 18 -микрофибрилла.

Схема биосинтеза белка: 1 — т-РНК с аминокислотами, 2 — т. РНК, 3 — Аминокислота, 4 — Малая субъединица рибосомы, 5 — и-РНК, 6 — Кодон, 7 — Антикодон, 8 — Большая субъединица рибосомы

Субъединицы рибосом синтезируются в ядре клетки Схема синтеза рибосом в клетках эукариот. 1. Синтез м. РНК рибосомных белков РНК полимеразой II. 2. Экспорт м. РНК из ядра. 3. Узнавание м. РНК рибосомой и 4. синтез рибосомных белков. 5. Синтез предшественника р. РНК (45 S — предшественник) РНК полимеразой I. 6. Синтез 5 S p. РНК полимеразой III. 7. Сборка большой рибонуклеопротеидной частицы, включающей 45 S-предшественник, импортированные из цитоплазмы рибосомные белки, а также специальные ядрышковые белки и РНК, принимающие участие в созревании рибосомных субчастиц. 8. Присоединение 5 S р. РНК, нарезание предшественника и отделение малой рибосомной субчастицы. 9. Дозревание большой субчастицы, высвобождение ядрышковых белков и РНК. 10. Выход рибосомных субчастиц из ядра. 11. Вовлечение их в трансляцию.

• Полисома, или полирибосома - несколько рибосом, одновременно транслирующих одну молекулу и. РНК. Поскольку длина средней молекулы м. РНК значительно превышает количество нуклеотидов, занимаемых на РНК рибосомой, одну молекулу РНК, в зависимости от скорости инициации одновременно транслируют несколько рибосом. Образование и количество рибосом в полисоме зависит от скорости инициации, элонгации и терминации на данной конкретной РНК.

Опорно-двигательные структуры клетки. Цитоскелет • Микротрубочки представляют собой прямые, неветвящиеся длинные полые цилиндры. Их внешний диаметр составляет около 24 нм, внутренний просвет имеет ширину 15 нм, а толщина стенки — 5 нм. Стенка микротрубочек построена за счет плотно уложенных округлых субъединиц (из белка тубулина) величиной около 5 нм.

• Полагают, что в клетке тубулины существуют в двух формах — свободной и связанной. Сдвиг равновесия между этими формами может привести или к диссоциации (распаду) микротрубочек, или к их росту. Ни тубулины в чистом виде, ни построенные из них микротрубочки не способны к сокращению, они не обладают АТФ-азной активностью. Они выполняют роль жестких каркасных структур. В клетках микротрубочки принимают участие в создании ряда временных (цитоскелет интерфазных клеток, веретено деления) или постоянных структур (центриоли и базальные тельца, реснички, жгутики).

• Микротрубочки интерфазных клеток • Практически во всех эукариотических клетках в гиалоплазме можно видеть длинные неветвящиеся микротрубочки. В больших количествах они обнаруживаются в цитоплазматических отростках нервных клеток, фибробластов и других изменяющих свою форму клеток. Они могут быть выделены сами или можно выделить образующие их белки: это те же тубулины со всеми их свойствами. • Главное функциональное значение таких микротрубочек цитоплазмы заключается в создании эластичного, но одновременно устойчивого внутриклеточного каркаса (цитоскелета), необходимого для поддержания формы клетки.
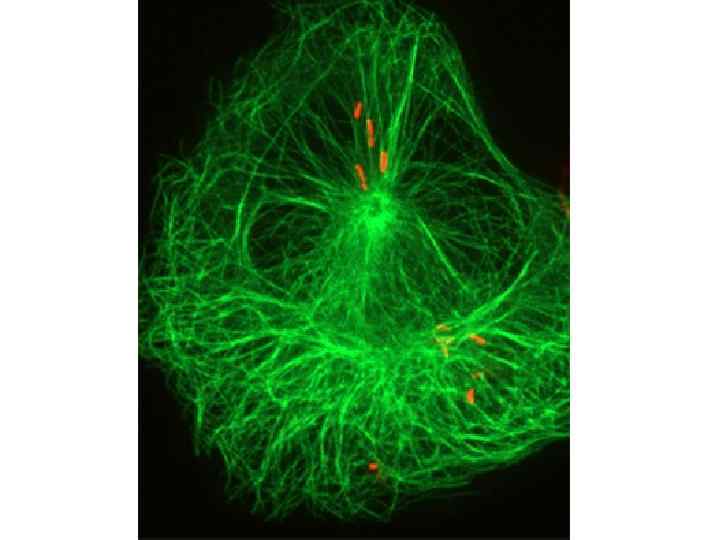

• Создавая внутриклеточный скелет, микротрубочки могут быть факторами ориентированного движения клетки в целом и ее внутриклеточных компонентов, задавать своим расположением векторы для направленных потоков разных веществ и для перемещения крупных структур. Разрушение микротрубочек, например колхицином, нарушает транспорт веществ в аксонах нервных клеток, приводит к блокаде секреции и другим нарушениям транспорта веществ. С цитоплазматическими микротрубочками связаны специальные белки, участвующие в механическом переносе отдельных внутриклеточных компонентов: микровакуолей, рибосом, митохондрий, других органелл. Перемещение кинезином меланосомы (клеточной органеллы) вдоль микротрубки

Транспорт в направлении к аппарату Гольджи осуществляет моторный белок динеин, доставку созревших в аппарате Гольджи белков по отходящим от центросомы микротрубочкам во все части клетки осуществляет моторный белок
 клетке система микротрубочек развивается в связи с особой клеточной")
• В неделящейся (интерфазной) клетке система микротрубочек развивается в связи с особой клеточной органеллой — центриолью, которая является местом, где происходит начальная полимеризация тубулинов и рост микротрубочек цитоскелета. Обычно в интерфазных клетках всегда присутствуют две центриоли, располагающиеся рядом друг с другом, образуя диплосому. В диплосоме центриоли располагаются под прямым углом по отношению друг к другу. Из двух центриолей различают материнскую и дочернюю. Обе центриоли сближены и расположены так, что конец дочерней центриоли направлен к поверхности материнской центриоли.

• Центриоли • Этот термин был предложен Т. Бовери в 1895 г. для обозначения очень мелких телец, размер которых находится на границе разрешающей способности светового микроскопа. В некоторых объектах удавалось видеть, что мелкие плотные тельца — центриоли (centriolum), обычно расположенные в паре — т. н. диплосома (diplosoma), окружены зоной более светлой цитоплазмы, от которой отходят радиально тонкие фибриллы (т. н. центросфера). Совокупность центриолей и центросферы называют клеточным центром (cytocentrum). Эти органеллы в делящихся клетках принимают участие в формировании веретена деления и располагаются на его полюсах. В неделящихся (интерфазных) клетках центриоли часто определяют полярность клеток, например, эпителия, и располагаются вблизи комплекса Гольджи.
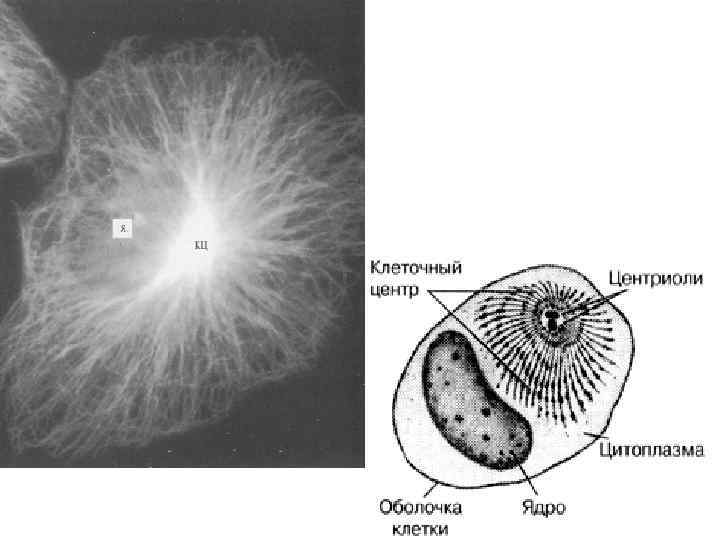

а—ультраструктурные особенности клеточного центра (/— центриоль; 2 — комплекс Гольджи; 3— десмосома; 4 — митохондрия; 5 — мембрана митохондрии; 6 — микротрубочки; 7 — шероховатый эндоплазматический ретикулум; 8 — гладкий эндо-плазматический ретикулум; 9 — межклеточное пространство) б — схема организации клеточного центра и центриоли (/ — центриоли; 2 — триплеты микротрубочек; 3 — микротрубочки; 4 — сателлиты)

• Тонкое строение центриолей удалось изучить только с помощью электронного микроскопа. Основой строения центриолей являются расположенные по окружности 9 триплетов микротрубочек (triplomicrotubuli), образующих таким образом полый цилиндр. Его ширина около 0, 2 мкм, а длина — 0, 3— 0, 5 мкм (хотя встречаются центриоли, достигающие в длину нескольких микрометров)

• Кроме микротрубочек в состав центриоли входят дополнительные структуры — «ручки» , соединяющие триплеты. Соединительные «ручки» построены из белка динеина, обладающего АТФ-азной активностью и обеспечивающего движение микротрубочек друг относительно друга. Системы микротрубочек центриоли можно описать формулой: (9 x 3) + 0, подчеркивая отсутствие микротрубочек в ее центральной части.

• При подготовке клеток к митотическому делению происходит удвоение центриолей. Этот процесс у различных объектов происходит в разное время — в течение синтеза ядерной ДНК или после него. Он заключается в том, что две центриоли в диплосоме расходятся и около каждой из них возникает заново по одной новой дочерней, так что в клетке перед делением обнаруживаются две диплосомы, т. е. четыре попарно связанные центриоли. Этот способ увеличения числа центриолей был назван дупликацией. Важно отметить, что увеличение числа центриолей не связано с их делением, почкованием или фрагментацией, а происходит путем образования зачатка, процентриоли, вблизи и перпендикулярно к исходной центриоли.
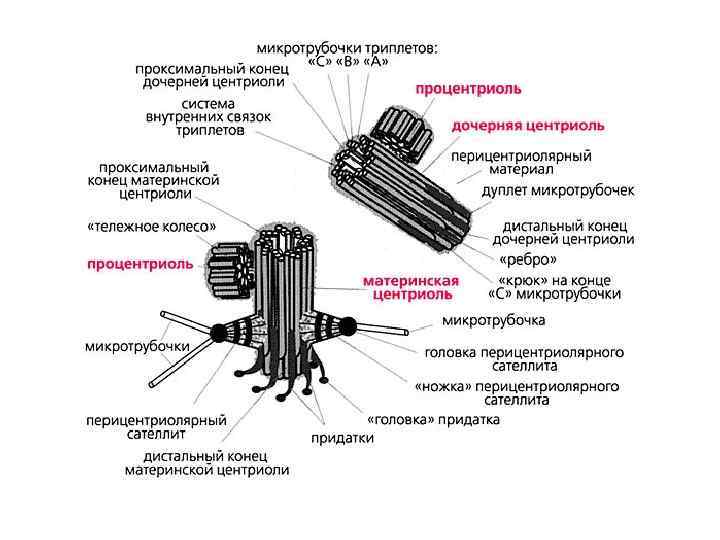

• Полагают, что центриоли участвуют в индукции полимеризации тубулина при образовании микротрубочек. Так, в интерфазе именно в связи с центриолью происходит рост микротрубочек клеточного каркаса. Перед митозом центриоль является одним из центров полимеризации микротрубочек веретена клеточного деления. Центриоль — центр роста микротрубочек аксонемы ресничек или жгутиков. Наконец, она сама индуцирует полимеризацию тубулинов новой процентриоли, возникающей при ее дупликации.

Реснички и жгутики • • Это специальные органеллы движения, встречающиеся в некоторых клетках различных организмов. В световом микроскопе эти структуры выглядят как тонкие выросты клетки. Длина ресничек 5— 10 мкм, а длина жгутиков может достигать 150 мкм. В единственном типе клеток человека имеющих жгутик сперматозоидах содержится по одному жгутику длиной 50 -70 мкм Схематическое изображение эпителия трахеи человека (по Rhodin, 1966). Четыре типа клеток: 1 — реснитчатые; 2 — бокаловидные; 3 — промежуточные и 4 — базальные. Многорядный эпителий слизистой оболочки полости носа: / - мерцательные реснички; 2 -- бокаловидные клетки; 3 --- замещающие клетки; 4 ■-- базальнан мембрана

Ресничка представляет собой тонкий цилиндрический вырост цитоплазмы с постоянным диаметром 200 нм. Этот вырост от основания до самой его верхушки покрыт плазматической мембраной. Внутри выроста расположена аксонема ( «осевая нить» ) — сложная структура, состоящая в основном из микротрубочек. Проксимальная часть реснички (базальное тело) погружена в цитоплазму. Диаметры аксонемы и базального тельца одинаковы (около 150 нм). Базальное тельце по своей структуре очень сходно с центриолью. Оно также состоит из 9 триплетов микротрубочек, имеет динеиновые «ручки» . Часто в основании реснички лежит пара базальных телец, располагающихся под прямым углом друг к другу подобно диплосоме. Аксонема (filamentum axiale) в своем составе имеет в отличие от базального тельца или центриоли 9 дублетов микротрубочек с динеиновыми «ручками» , образующих стенку цилиндра аксонемы. Кроме периферических дублетов микротрубочек, в центре аксонемы располагается пара центральных микротрубочек. В целом систему микротрубочек реснички описывают как (9 x 2) + 2 в отличие от (9 x 3) + 0 системы Ресничка — тонкий вырост на поверхности клетки. Стержень реснички центриолей и базальных телец. Базальное тельце и аксонема структурно связаны друг с образован аксонемой — системой микротрубочек 9+2. В основании другом и составляют единое целое: две реснички расположено базальное тельце, служащее матрицей для формирования аксонемы. микротрубочки триплетов базального тельца являются микротрубочками дублетов аксонемы.

Основной белок ресничек — тубулин — не способен к сокращению, укорочению. Вероятным кандидатом на роль сократимого белка считается белок «ручек» — динеин, так как он обладает АТФ-азной активностью. В последние годы для объяснения способа движения ресничек и жгутиков используется гипотеза «скользящих нитей» . Известно, что сокращение мышечных волокон происходит за счет встречного скольжения фибрилл двух мышечных белков: миозина и актина; при этом также не происходит собственно укорачивания или сокращения отдельных мышечных белковых фибрилл. Предполагается, что незначительные смещения дублетов микротрубочек друг относительно друга могут вызвать изгиб всей реснички, а если такое локальное смещение будет происходить вдоль жгутика, то может возникнуть волнообразное его движение. Аксонема состоит из комплекса микротрубочек и связанных с ними белков. 9 пар микротрубочек расположено по окружности, одна пара находится в центре. Каждая периферическая пара образована субфибриллой A и субфибриллой B. Субфибриллы состоят из протофиламентов. Обладающий АТФазной активностью белок динеин — компонент тубулин-динеинового хемомеханического преобразователя — входит в состав ручек, связанных с субфибриллой A
 Содержит 2 центриоли (4 -5) От одной из них")
Шейка, или связующий отдел (а) Содержит 2 центриоли (4 -5) От одной из них начинается аксонема (7), или осевая нить хвоста, образованная микротрубочками по схеме (9? 2) + 2. Промежуточная часть (б) В этой части вокруг аксонемы - 9 наружных фибрилл, митохондриальная спиральная оболочка (6) и плазмолемма. Главная, или основная часть (в) Здесь вокруг аксонемы фибриллярное влагалище (8) (9 наружных фибрилл и волокнистая оболочка), а также плазмолемма. Концевая часть (г) В этой части вокруг аксонемы остаётся только плазмолемма.

Свободные клетки, имеющие реснички и жгутики, обладают способностью двигаться, а неподвижные клетки движением ресничек могут перемещать жидкость и частицы. Траектория движения ресничек очень разнообразна. В различных клетках это движение может быть маятникообразным, крючкообразным, воронкообразным или волнообразным. • Движения жгутика (хвоста) сперматозоида (А) и реснички мерцательного эпителия (Б)

Микрофиламенты • встречаются практически во всех типах клеток. По строению и функциям они бывают разные, однако отличить их морфологически друг от друга трудно. Располагаются микрофиламенты в кортикальном слое цитоплазмы, непосредственно под плазмолеммой, пучками или слоями.

• Сеть микрофиламентов выявлена в большинстве клеток. Они отличаются по химическому составу. В зависимости от их химического состава они могут выполнять функции цитоскелета и участвовать в обеспечении движения. • С помощью иммунофлюоресцентных методов четко показано, что в состав микрофиламентов кортикального слоя и пучков входят сократительные белки: актин, миозин, тропомиозин, аактинин. Следовательно, микрофиламенты не что иное, как внутриклеточный сократительный аппарат, обеспечивающий не только подвижность клеток при активном амебоидном их перемещении, но, вероятно, и большинство внутриклеточных движений, таких как токи цитоплазмы, движение вакуолей, митохондрий, деление клетки.

Организация микроворсинки в апикальной части каёмчатой клетки. Около 30 параллельно идущих микрофиламентов образуют стержень микроворсинки. (+)– Концы двух переплетённых нитей F– актина микрофиламентов направлены к вершине микроворсинки. Микрофиламенты заякорены цитоплазматическими концами в терминальной сети. Терминальная сеть — густое сплетение молекул спектрина, сшивающих примембранные микрофиламенты. Непосредственно под терминальной сетью расположено сплетение промежуточных филаментов. Микрофиламенты скреплены актинсвязывающими белками фимбрином и фасцином. К внутренней поверхности плазматической мембраны микрофиламенты присоединены при помощи минимиозина
")
Промежуточные филаменты, или микрофибриллы • • • тоже белковые структуры. Это тонкие (10 нм) неветвящиеся, часто располагающиеся пучками нити. Характерно, что их белковый состав различен в разных тканях. Роль промежуточных микрофиламентов скорее всего опорно-каркасная, однако эти фибриллярные структуры не так лабильны, как микротрубочки. В последнее время с помощью иммуноморфологических методов стало возможным определить тканевое происхождение тех или иных опухолей именно по белкам их промежуточных филаментов, что очень важно для правильного выбора типа химиоте-рапевтических противоопухолевых препаратов.

• • • • • Тип I — кератины Данный тип белков делится на 2 подсемейства: кислые кератины, нейтральные и основные кератины. Димер кератина состоит из одного кислого и одного основного кератина. Среди многочисленных изоформ кератина выделяют две основные группы — эпителиальные кератины (цитокератин), включающую около 20 видов кератинов, и кератины волос (примерно 10 видов), из которых построены также ногти, рога и чешуя пресмыкающихся. Тип II виментин - характерный для клеток мезенхимного происхождения: входит в состав клеток соединительной ткани, эндотелия, клеток крови; десмин; глиальный фибриллярный кислый белок; периферин. Тип III Альфа-интернексин Белки нейрофиламентов Нестин Синемин Синкойлин Общая схема Тип IV структурной организации Ядерные ламины промежуточных филаментов

• Белки промежуточных филаментов различных клеток • • • Белок • • Десмин Клетки мышечных тканей Структурируют сократительные органеллы миофибриллы. Маркёры опухолей мышечного происхождения(рабдомиосаркома) • • Виментин Клетки мезенхимного генеза: Формируют сетевидную структуру вокруг ядра клетки фибробласты, ГМК, эндотелиальные, Маркёры опухолей соединительной ткани хондробласты, остеобласты, макрофаги (фибросаркома, липосаркома, ангиосаркома, хондросаркома, остеосаркома) • • • Глиальный фибриллярный кислый белок Астроциты, олигодендроциты, Структурная поддержка цитоплазмы шванновские клетки Маркёры опухолей, возникающих . из глиальных клеток (астроцитомы) • • Белки нейрофиламентного триплета Нейроны Поддерживают структуру и форму отростков нейрона (аксонов и дендритов) • • • Ламины A, B и C Ядерная пластинка Организуют ядерную пластинку и лежащий около неё хроматин Локализация Некоторые специальные функции Цитокератин (29 изоформ) Эпителиальные клетки Создают тянущее усилие, взаимодействуют с десмосомами и полудесмосомами. Маркёры опухолей эпителиального происхождения (плоскоклеточная карцинома, аденокарцинома)
 — необязательные компоненты клетки, возникающие и исчезающие")
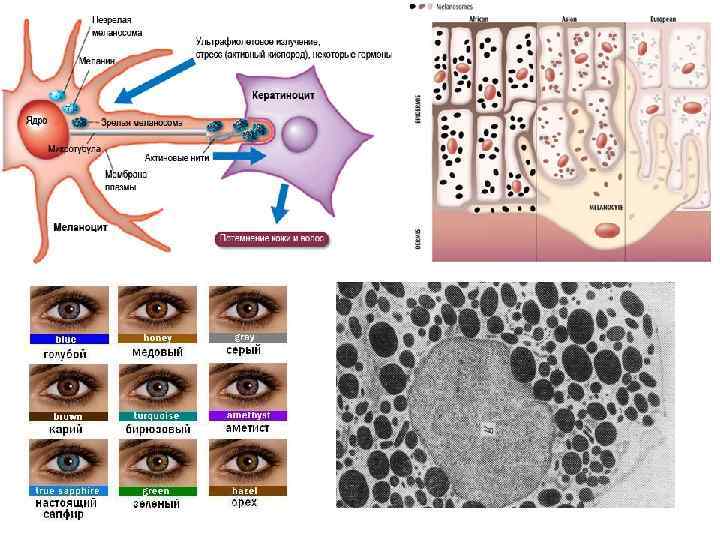
• • Включения цитоплазмы (inclusiones cytoplasmicae) — необязательные компоненты клетки, возникающие и исчезающие в зависимости от метаболического состояния клеток. Различают включения трофические, секреторные, экскреторные и пигментные. К трофическим включениям относятся капельки нейтральных жиров, которые могут накапливаться в гиалоплазме. В случае недостатка субстратов для жизнедеятельности клетки эти капельки могут резорбироваться. Другим видом включений резервного характера является гликоген — полисахарид, откладывающийся также в гиалоплазме (рис. 15). Отложение запасных белковых гранул обычно происходит в связи с активностью эндоплазматической сети. Так, запасы белка вителлина в яйцеклетках амфибии накапливаются в вакуолях эндоплазматической сети. Секреторные включения — обычно округлые образования различных размеров, содержащие биологически активные вещества, образующиеся в клетках в процессе жизнедеятельности. Экскреторные включения не содержат каких-либо ферментов или других активных веществ. Обычно это продукты метаболизма, подлежащие удалению из клетки. Пигментные включения могут быть экзогенные (каротин, пылевые частицы, красители и др. ) и эндогенные (гемоглобин, гемосидерин, билирубин, меланин, липофусцин). Наличие их в цитоплазме может изменять цвет ткани органа временно или постоянно. Нередко пигментация (или депигментация) ткани служит диагностическим признаком.

• Жир и гликоген в печени

Рис. 1. Гладкая мышечная клетка кишечника. Рис. 2. Жировая клетка из подкожной клетчатки крысы. Рис. 3. Митохондрии и секреторные гранулы о клетках поджелудочной железы. Рис. 4. Аппарат Гольджи в чувствительной клетке спинномозгового ганглия. Рис. 5. Аппарат Гольджи и секреторные гранулы в клетках поджелудочной железы.
. Слева - секреторные клетки частично заполнены молоком,")
• Альвеолы молочной железы (упрощенное изображение). Слева - секреторные клетки частично заполнены молоком, справа - клетки после выделения молока в полость альвеолы (изображение молока условное)
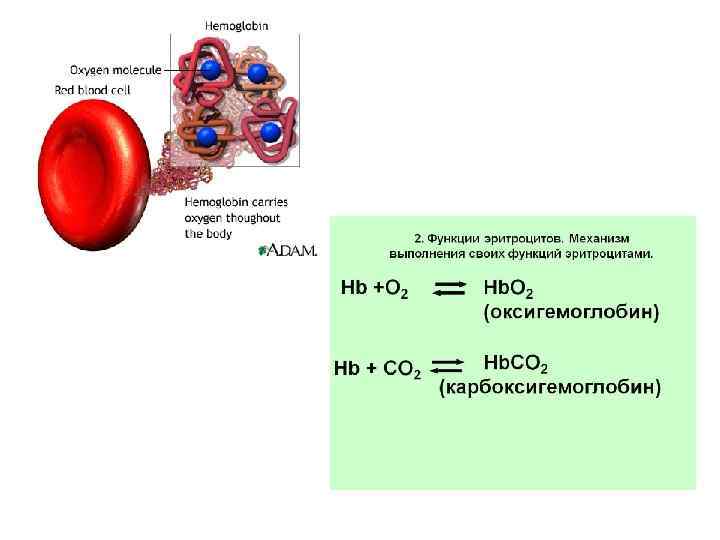

• Гемосидерин — это пигмент темно-желтого цвета, представляющий собой гидроокись железа, связанную с белками, гликозаминогликанами и липидами клетки. Он образуется при распаде гемоглобина на гем и ферритин, с последующей денатурацией и депротеинизацией белка ферритина. Наличие железа в гемосидерине позволяет с помощью специфичных реакций (например, реакция Перлса с берлинской лазурью) выявить и дифференцировать его от сходных пигментов (гемомеланин, липофусцин, меланин). Гемосидерин обнаруживается в ретикулярных и эндотелиальных клетках селезенки, печени, костного мозга, лимфатических узлах. В межклеточном веществе гемосидерин подвергается фагоцитозу сидерофагами. Гемосидерин образуется при кровоизлияниях. В небольших кровоизлияниях обнаруживается только гемосидерин, а в крупных - по периферии, среди живой ткани образуется гемосидерин, а в центре - кровоизлияния, где аутолиз происходит без доступа кислорода и участия клеток, появляются кристаллы гематоидина. Накопление гемосидерина в тканях и органах происходит при различных заболеваниях.

• Билирубин – это желто-зеленый пигмент, который образуется в результате распада гемоглобина, и содержится в сыворотке крови и в желчи. В обмене билирубина большую роль играет печень. Повышение уровня билирубина в крови сопровождается желтухой (пожелтение склер глаз, кожи тела и слизистых оболочек) и чаще всего является симптомом некоторых заболеваний печени (гепатит) или крови (гемолитическая анемия).

миоцит
 – заболевание, вызванное отложением в различных органах и тканях организма серебра. Является")
Аргироз (аргирия) – заболевание, вызванное отложением в различных органах и тканях организма серебра. Является следствием длительного приема препаратов серебра или его попадания в организм в связи с профессиональными вредностями. • Пол Карасон (Paul Karason) из Калифорнии посинел в результате многолетнего употребления коллоидного серебра, которым он лечил дерматит.

• Во многих клетках органеллы могут принимать участие в образовании особых структур, характерных для специализированных клеток. Так, реснички и жгутики образуются за счет центриолей и плазматической мембраны

микроворсинки — это выросты плазматической мембраны с гиалоплазмой и микрофиламентами

акросома спермиев — это производное элементов аппарата Гольджи,
 и колбочки (Б)")
• «эллипсоид» зрительных клеток — скопления митохондрий Строение палочки (А) и колбочки (Б) сетчатки позвоночных 1, 2— наружный и внутренний сегменты, 3 — масляная капля, 4 — эллипсоид (митохондрии), 5 — параболоид, 6 — ядро, 7 — синаптическая область. К П
МОРФОЛОГИЯ 3.ppt