Обмен углеводов в норме и патологии.ppt
- Количество слайдов: 29



Обмен углеводов в норме и патологии

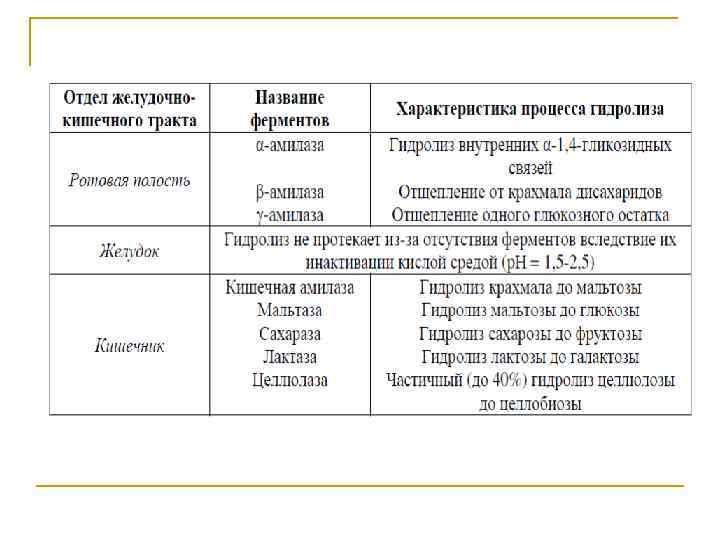
Переваривание и всасывание Переваривание углеводов в желудочно кишечном тракте это часть метаболизма углеводов, совокупность управляемых процессов, осуществляемых в системе пищеварения, которые представляют собой химическую переработку углеводов, поступающих в организм с пищей, для последующего их всасывания в кровь и в лимфу.

Пищевые вещества содержат крупномолекулярные белки, углеводы и липиды, которые не способны к всасыванию в кровь и лимфу из за больших размеров своих молекул. Химическая переработка пищи в желудочно кишечном тракте представляет собой последовательное ступенчатое ферментативное гидролитическое расщепление крупномолекулярных белков, углеводов и липидов до простых веществ, способных к всасыванию.

В числе углеводов потребляемой пищи содержатся полисахариды крахмал и гликоген. Расщепление этих углеводов начинается в полости рта и продолжается в желудке. Катализатором гидролиза является фермент α‑амилаза слюны. При расщеплении из крахмала и гликогена образуются декстрины и в небольшом количестве мальтоза. Пережеванная и смешанная со слюной пища проглатывается и попадает в желудок. Проглоченные пищевые массы со стороны поверхности полости желудка постепенно перемешиваются с желудочным соком, содержащим соляную кислоту.

Содержимое желудка с периферии приобретает значительную кислотность (p. H = 1, 5 ÷ 2, 5). Такая кислотность дезактивирует амилазу слюны. В то же время в толще массы желудочного содержимого амилаза слюны некоторое время продолжает действовать и происходит расщепление полисахаридов с образованием декстринов и мальтозы. Желудочный сок не содержит ферментов, расщепляющих сложные углеводы. Поэтому гидролиз углеводов с увеличением кислотности в желудке прерывается и возобновляется в двенадцатиперстной кишке.

В двенадцатиперстной кишке происходит наиболее интенсивное переваривание крахмала и гликогена с участием α‑амилазы сока поджелудочной железы. В двенадцатиперстной кишке кислотность значительно снижается. Среда становится практически нейтральной, оптимальной для максимальной активности α‑амилазы панкреатического сока. Поэтому гидролиз крахмала и гликогена с образованием мальтозы, который начинался в полости рта и в желудке с участием α‑амилазы слюны, в тонкой кишке завершается. Процессу гидролиза с участием α‑амилазы панкреатического сока дополнительно способствуют еще два фермента: амило 1, 6 глюкозидаза и олиго 1, 6 глюкозидаза (терминальная декстриназа).
")
Образовавшаяся в результате начальных стадий гидролиза углеводов мальтоза, гидролизуется с участием фермента мальтазы (α‑глюкозидазы) с образованием двух молекул глюкозы. Пищевые продукты могут содержать углевод сахарозу. Сахароза расщепляется при участии сахаразы фермента кишечного сока. При этом образуются глюкоза и фруктоза.
 могут содержать углевод лактозу. Лактоза гидролизуется с участием фермента кишечного сока")
Пищевые продукты (молоко) могут содержать углевод лактозу. Лактоза гидролизуется с участием фермента кишечного сока лактазы. В результате гидролиза лактозы образуются глюкоза и галактоза. Таким образом углеводы, содержащиеся в пищевых продуктах, расщепляются на составляющие их моносахариды: глюкозу, фруктозу и галактозу. Конечные стадии гидролиза углеводов осуществляются непосредственно на мембране микроворсинок энтероцитов в их гликокаликсе.
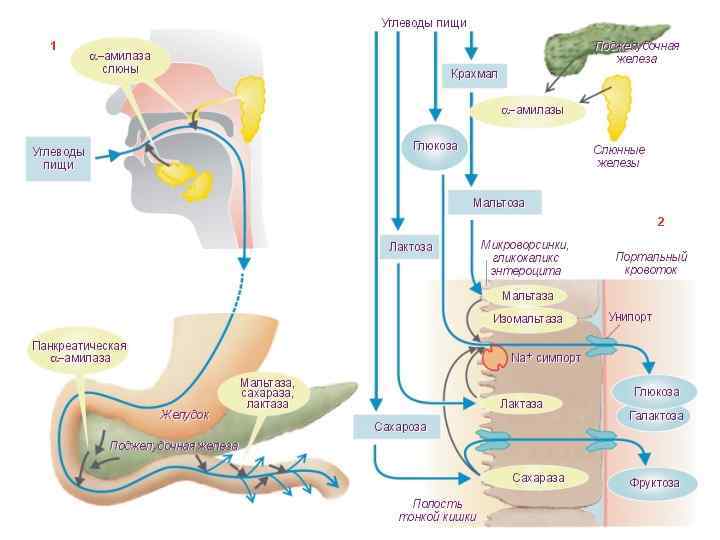

n Итак, завершающие стадии гидролиза углеводов пищи и всасывание углеводов пищи тесно сопряжены (мембранное пищеварение). Моносахариды и небольшое количество дисахаридов и всасываются энтероцитами тонкой кишки и попадают в кровь. Интенсивность всасывания моносахаридов различна. Всасывание маннозы, ксилозы и арабинозы осуществляется преимущественно путем простой диффузии. Всасывание же большинства других моносахаридов происходит за счет активного транспорта. Легче других моносахаридов всасываются глюкоза и галактоза.

Мембраны микроворсинок энтероцитов содержат системы переносчиков, способных связывать глюкозу и Na+ и переносить их через цитоплазматическую мембрану энтероцита в его цитозоль. Энергия, необходимая для такого активного транспорта, образуется при гидролизе АТФ. Большая часть моносахаридов, всосавшихся в русло кишечных ворсинок, попадают с потоком крови через воротную вену в печень. Небольшое количество (~10%) моносахаридов поступает по лимфатическим сосудам в венозную систему. В печени значительная часть всосавшейся глюкозы превращается в гликоген. Гликоген резервируется в клетках печени (гепатоциты) в виде гранул.

Запомните !!! 1. В тонком кишечнике происходит всасывание только моносахаридов (глюкозы, фруктозы, галактозы и некоторых других), образующихся при гидролизе крахмала, гликогена и дисахаридов или поступающих в кишечник с пищей. 2. Всасывающиеся в кишечнике в небольшом количестве дисахариды не усваиваются организмом и выводятся с мочой в неизмененном виде. 3. Структурный полисахарид растений целлюлоза не расщепляется амилазой поджелудочной железы и не может служить источником углеводов для человека, хотя играет важную роль в обеспечении нормальной функции кишечника, в частности, его моторной функции.

Промежуточный обмен n n Основные биохимические превращения всосавшихся в кишечнике моносахаридов осуществляются в клетках печени и других органов. Здесь происходит превращение фруктозы и галактозы в глюкозу, которая включается затем в многочисленные ферментативные реакции, лежащие в основе энергообеспечения клеток и служащие источником ряда веществ, необходимых для синтеза белков, жиров и углеводов. Метаболизм глюкозы в клетках любого органа, в том числе печени, начинается с ее фосфорилирования и образования глюкозо 6 фосфата. Эта реакция протекает в цитоплазме клеток с использованием одной молекулы АТФ. В связи с неспособностью глюкозо 6 фосфата проходить через мембраны клеток, он накапливается в цитоплазме в достаточно больших количествах.

Распад углеводов в клетке происходит двумя путями: Аэробным – при достаточном обеспечении клетки кислородом n Анаэробным – при его недостатке. n
")
Основные пути распада углеводов Анаэробный распад Аэробный процесс Непрямой Прямой Глюкоза ( 6 С-атомов) Глюкоза-6 -фосфат (6 С-атомов) 2 молекулы пировиноградной кислоты (2 молекулы по 3 С-атомов) Молочная кислота(2 молекулы) Ацетил-Ко. А ( 2 молекулы) Пентозы (5 молекулы) Спирт( 2 молекулы) Н 2 О и СО 2 ( по 4 молекулы) НАДФН 2( 12 молекулы) Энергия – 2 АТФ (196, 8 к. Дж/моль) Энергия – 38 АТФ (2872, 1 к. Дж/моль) Энергия – 36 АТФ (2720, 9 к. Дж/моль)

n Как видно из схемы, основным исходным веществом для дальнейших превращений является активированная глюкоза в виде глюкозо 6 фосфата. Распад ее в анаэробных условиях и аэробных протекает одинаково до стадии образования пировиноградной кислоты. Этот процесс называют гликолизом.
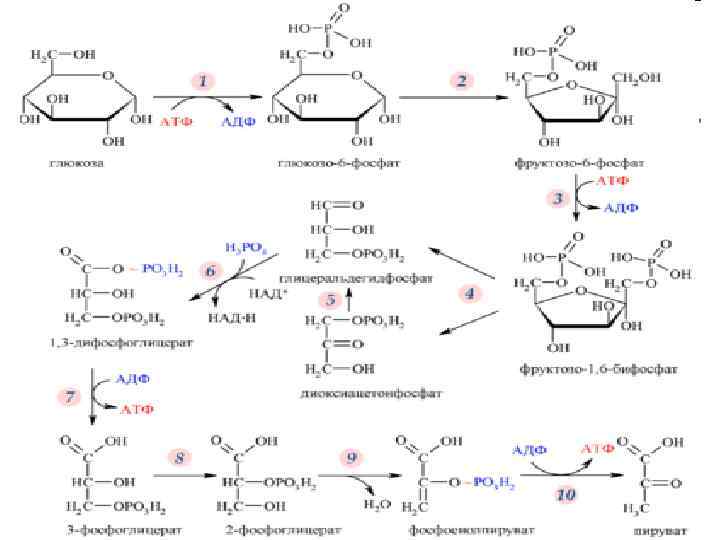

АНАЭРОБНЫЙ РАСПАД УГЛЕВОДОВ n n Ферментативная диссимиляция углеводов в анаэробных условиях, происходящая с выделением энергии и приводящая к образованию продуктов неполного окисления, называется брожением. При брожении акцептором водорода служат органические соединения, получающиеся в реакциях окисления (например, уксусный альдегид при спиртовом брожении); кислород в этих реакциях не участвует.

n 1. Образуются фосфорные эфиры Сахаров. Под действием фермента гексокиназы и адениловых кислот, являющихся донаторами и акцепторами фосфорной кислоты, глюкоза превращается в глюкопиранозо 6 фосфат. Адениловые кислоты в дрожжах содержатся в виде аденозинмонофосфата (АМФ), аденозиндифосфата (АДФ) и адеыозинтрифосфата (АТФ). Гексокиназа катализирует перенос одной фосфорной группы с АТФ на глюкозу. При этом АТФ превращается в АДФ, а остаток фосфорной кислоты присоединяется по месту шестого углеродного атома. Действие фермента активируется ионами магния. Подобным образом происходит превращение D фруктозы и D маннозы. Глюкокиназная реакция определяет скорость процесса брожения.

2. Глюкозо 6 фосфат под действием фермента глюкозофосфат — изомеразы подвергается изомеризации — превращению в фруктозо 6 фосфат. Реакция обратима и сдвинута в сторону фруктозо 6 фосфата. 3. Фруктозо 6 фосфат под действием фермента фосфофруктокпназы присоединяет по месту первого углеродного атома второй остаток фосфорной кислоты за счет АТФ и превращает в фруктозо 1, 6 дифосфат. Эта реакция практически необратима. Молекула сахара переходит в оксоформу и становится лабильной, способной к дальнейшему превращению, так как ослабляется связь между третьим и четвертым углеродными атомами.
")
4. Под действием фермента альдолазы (активируемой ионами Zn 2+, Со 2+ и Са 2+) фруктозе 1, 6 дифосфат распадается на две фосфотриозы — З фосфоглицериновый альдегид и фосфодиоксиацетон. Эта реакция обратима. 5. Между фосфотриозами происходит реакция изомеризации, катализируемая ферментом триозофосфатизомеразой. Равновесие устанавливается при 95% 3 фосфоглицеринового альдегида и 5% фосфодиоксиацетона.

6. В индукционный период, пока в качестве промежуточного продукта не образовался уксусный альдегид, между двумя молекулами 3 фосфоглицеринового альдегида под действием фермента альдегидмутазы при участии молекулы воды происходит реакция дисмутации. При этом одна молекула фосфоглицеринового альдегида восстанавливается, образуя фосфоглицерин, другая окисляется в 3 фосфоглицериновую кислоту. Фосфоглицерин в дальнейших реакциях не участвует н после отщепления фосфорной кислоты является побочным продуктом спиртового брожения.

7. фермента 8. действием Под отдачи результате В 9. При участии фермента фосфотрансферазы остаток фосфорной кислоты, содержащий макроэргическую связь, передается с 1, 3 дифосфоглицериновой кислоты на АДФ с образованием АТФ и 3 фосфоглицериновой кислоты. Энергия, освобождающаяся при окислении фосфоглицеринового альдегида, резервируется в АТФ. фосфоглицеромутазы 3 фосфоглицериновая кислота изомеризуется в 2 фосфоглицериновую кислоту. вызываемой перераспределением внутримолекулярной энергии, 2 фосфоглицериновая кислота превращается в фосфоэнолпировиноградную кислоту, содержащую макроэргическую связь. Реакцию катализирует энолаза, 2+, Мn 2+ и Zn 2+. активируемая ионами Мg

10. Под действием фермента фосфотрансферазы в присутствии ионов К+ ос таток осфорной кислоты ф передается от фосфоэнолпировиноградной кислоты на АДФ, резервируя энергию в АТФ. 11. Образовавшаяся энолпировшюградная кислота превращается в более ста бильную кетоформу. 12. Под действием фермента карбоксилазы от пировиноградной кислоты от щепляется диоксид углерода и образуется уксусный альдегид. 13. Фермент алкогольдегидрогеназа переносит водород с восстановленного НАД Н 2 на уксусный альдегид, в результате чего образуется этиловый спирт и регенерируется НАД.

АЭРОБНЫЙ РАСПАД УГЛЕВОДОВ n В условиях аэробиоза распад углеводов до образования пировиноградной кислоты происходит так же, как и при анаэробиозе, но в отличие от него пировиноградная кислота полностью окисляется до диоксида углерода и воды в цикле трикарбоновых кислот — ЦТК (цикле Кребса, лимоннокислотном цикле). В этом цикле последовательно протекают окислительно восстановительные реакции, в которых под действием специфических дегидрогеназ происходит перенос водорода на молекулярный кислород — конечный его акцептор. Однако перенос осуществляется не непосредственно, ; а через молекулы переносчики, образующие так называемую дыхательную цепь. Схема химических превраще ний при аэробном распаде глюкозы приведена ниже.

При катаболизме глюкозы образуются две молекулы пировиноградной кис лоты. Вначале одна из них подвергается реакциям окислительного декарбоксилирования, в результате которых образует ся цетил Ко. А а (активированная уксусная кислота): СНз·СО · СООН + Ко. АSН + НАД — СНз СО ~ Ко. АSН + НАД · Н 2 + СО 2 Вторая молекула пировиноградной кислоты под действием фермента пируваткарбоксилазы конденсируется с молекулой диоксида углерода с образова нием авелевоуксусной щ кислоты: СHз · COOH + CO 2 + АТФ ↔ HOOC · CH 2 · COOH + АДФ + Ф
. Собственно ЦТК начинается с кон")
При установившемся цикле щавелевоуксусная кислота образуется из яблочной (малата). Собственно ЦТК начинается с кон денсации цетил Ко. А а с молекулой щавелевоуксусной кислоты (оксалоацетата), катализируемой ферментом цитратсинтазой. Продуктами реакции являются лимонная кислота (цитрат) и свободный кофермент А. Дальнейшие превращения видны из схемы на рисунке. За один оборот молекулы пировиноградной кислоты присоединяется 3 молекулы Н 2 О, выделяется 5 Н 2 и образуется 3 молекулы СО 2: СНз · СООН + ЗН 2 О— ЗС 02 + 10 Н. В ЦТК «сжигаются» не только углеводы, но и жирные кислоты (после пред варительной еградации до ацетил Ко. А), а д также многие аминокислоты (после удаления аминогруппы в реакциях дезаминирования или переаминирования).

В ЦТК «сжигаются» не только углеводы, но и жирные кислоты (после предварительной деградации до ацетил Ко. А), а также многие аминокислоты (после удаления аминогруппы в реакциях дезаминирования или переаминирования). Аэробный и анаэробный распад углеводов доставляет дрожжам энергию и обеспечивает процессы синтеза биомассы различными предшественниками. Из щавелевоуксусной и а кетоглутаровой кис лот в результате восстановительного аминирования и переаминиро вания образуются соответственно аспарагиновая и глютаминовая кислоты. Аспарагиновая кислота может образовываться также из фумаровой кислоты. Синтез этих двух аминокислот занимает глав ное место в синтезе белков из углеводов. При конденсации фос фодиоксиацетона с альдегидами могут образовываться пентозы, гексозы и различные полисахариды. Для синтеза биомассы дрожжи используют и другие — анаплеротические — пути, например пентозофосфатный путь. Пентозофосфаты являются предшественниками нуклеотидов и нуклеиновых кислот. Так как при полном окислении сахара значительно больше освобождается энергии и образуется реакционноспособных метаболи тов для синтетических процессов, то возрастает скорость размно жения и увеличивается биомасса дрожжей.
Обмен углеводов в норме и патологии.ppt