Обмен нуклеиновых кислот.ppt
- Количество слайдов: 18



Обмен нуклеиновых кислот


Общие особенности биосинтеза ДНК, РНК и белка n специфичность - обеспечение точности передачи наследственной информации от родителей к потомкам в виде определенной последовательности нуклеотидов. n матричный синтез - дочерний полимер создается по матрице родительской ДНК, т. е. является ее копией. n наличие трёх этапов синтеза: 1) инициация (начало) 2) элонгация (продолжение) 3) терминация (окончание синтеза)

Основные принципы репликации ДНК 1. Репликация ДНК - полуконсервативный процесс: из одной материнской биспиральной молекулы ДНК образуются две дочерние биспиральные молекулы ДНК, идентичные другу и исходной молекуле ДНК. Каждая из дочерних молекул содержит одну неизменную родительскую цепь ДНК и одну вновь синтезированную цепь, т. е. является полуконсервативной по составу (содержит 50% материнской ДНК и 50 % новообразованной ДНК).
. 3. Репликация ДНК инициируется «затравкой» - отрезками")
2. Для репликации необходима матричная ДНК (матрица). 3. Репликация ДНК инициируется «затравкой» - отрезками РНК праймерами, содержащими от 50 до 100 нуклеотидных остатков. Синтез «затравочного» олигорибонуклеотида, или праймера, на материнской цепи ДНК катализируется РНК-полимеразой. 4. Репликация ДНК - процесс ферментативный. В нём участвуют ДНК-полимеразы (I, III), праймаза (РНК-полимераза), эндонуклеаза, ДНК-лигаза и др. , а также белковые факторы, выполняющие специфические функции: - ДНК-связывающий белок, который, соединяясь с ДНК, резко ослабляет взаимодействие полидезоксирибонуклеотидных цепей в ее молекуле, - ДНК-раскручивающий белок (ДНК-гираза, или топоизомераза), обеспечивающий раскручивание суперспиральной молекулы ДНК, - хеликаза, расплетающая двойную спираль ДНК на две цепи.

5. За процесс репликации ДНК ответственна ДНК-полимераза III : она осуществляет сборку дезоксиполирибонуклеотидной цепи в направлении 5'—> 3' путём переноса остатка нуклеозидмонофосфата от нуклеозидтрифосфата на концевой нуклеотидный остаток растущей полинуклеотидной цепи (на место атома водорода гидроксильной группы при третьем углеродном атоме дезоксирибозы концевого нуклеотида). При этом выделяется пирофосфат. За 1 сек. 1 молекула ДНК-полимеразы присоединяет ~ 10 нуклеотидов. ДНК-полимеразы I и II достраивают поврежденные участки в молекуле ДНК - осуществляют репарацию ДНК. 6. Биосинтез (элонгация дочерней цепи ДНК) осуществляется ступенчато, путем последовательного присоединения нуклеотидных мономеров к растущей полинуклеотидной цепи в направлении 5'->3'. ДНК-полимераза III катализирует образование фосфодиэфирной связи только в том случае, если основание очередного нуклеотида комплементарно соответствующему основанию матричной цепи.
;")
7. Репликация ДНК идёт при наличии всех четырёх типов дезоксирибонуклеозид-5'-трифосфатов (АТФ, ГТФ, УТФ, ЦТФ); ионов магния; ДНК-полимеразы III, которая катализирует последовательное присоединение дезоксирибонуклеиновых звеньев к 3'-гидроксильному концу молекулы-затравки, и наличии матрицы ДНК.

8. Синтез новой ДНК сопряжён с одновременным раскручиванием родительской ДНК. Участок, где происходит одновременное расплетание материнской ДНК и синтез дочерней цепи ДНК при участии фермента хеликаза, называется репликационной вилкой. 9. В области репликационной вилки обе цепи родительской ДНК служат матрицей для синтеза дочерней цепи ДНК. Цепи родительской ДНК антипараллельны. 10. Одна цепь (ведущая) синтезируется в направлении 5'->3' без разрывов. Репликация другой цепи материнской ДНК (отстающая) осуществляется «челночным способом» - короткими фрагментами (фрагменты Оказаки), следующими один за другим.

По мере репликации фрагменты Оказаки соединяются под действием ДНК-лигазы (катализирует образование фосфодиэфирной связи между 3'- и 5'-концами фрагментов Оказаки, закрепленных на комплементарной им непрерывной цепи матричной ДНК в присутствии АТФ):

Современная гипотетическая схема репликации ДНК
 n n n В качестве матрицы фермент РНК-полимераза использует")
Синтез РНК на ДНК-матрице (транскрипция) n n n В качестве матрицы фермент РНК-полимераза использует двухцепочечную ДНК, причем в клетке транскрибируется только одна цепь ДНК. Область связывания (специфическая последовательность нуклеотидов ДНК) РНК-полимеразы с матрицей называется промотором. Синтез молекулы РНК идёт комплементарно от 5'-конца к 3'концу матричной цепи ДНК. По мере продвижения РНК-полимеразы впереди её идёт разделение, а позади – восстановление двойной спирали ДНК. Временно образовавшийся «глазок» движется вдоль ДНК вместе с полимеразой. Синтез завершается, когда РНК-полимераза достигает терминирующей последовательности нуклеотидов (сайта терминации). Белковый фактор терминации облегчает отделение транскрипта от матрицы и транскрипция завершается. Участок ДНК, ограниченный промотором и сайтом терминации, представляет собой единицу транскрипции — транскриптон. У эукариот в состав транскриптона входит, как правило, один ген (участок ДНК, кодирующий синтез одного полипептида).
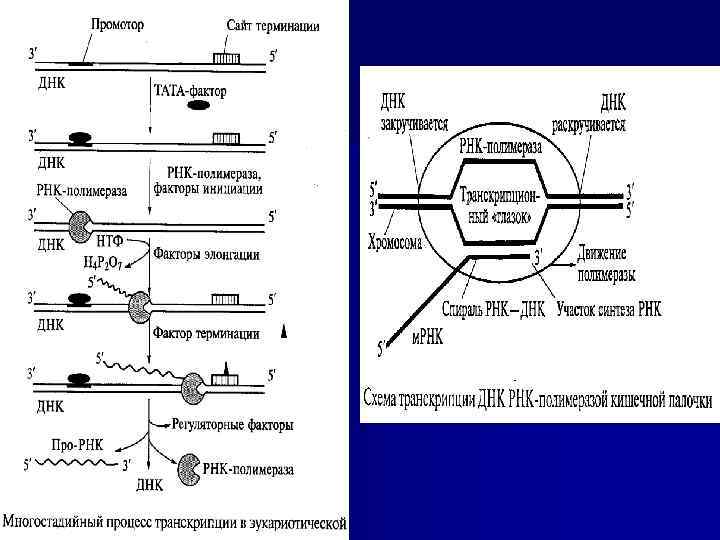

n n n Механизм синтеза РНК на ДНК является консервативным: продукт синтеза не включает каких-либо компонентов матрицы. РНК, синтезированная в присутствии различных ДНК-матриц, характеризуется нуклеотидным составом, комплементарным одной из цепей матрицы ДНК. Количество синтезированной РНК может в сотни раз превосходить количество ДНК, взятой для затравки, т. е. одна и та же матрица многократно используется для синтеза РНК. С помощью РНК-полимеразы in vivo синтезируются различные м. РНК, т. РНК и р. РНК, причем у эукариот основная масса РНК синтезируется в ядре клетки. Как правило, все виды РНК синтезируются в виде РНКпредшественников (про-РНК). Они имеют большую молекулярную массу. В дальнейшем в процессе «созревания» (процессинга) утрачивается неинформативная (не несущая функциональной нагрузки) часть РНК-предшественника, изменяется его информативная часть путем ее метилирования (особенно в случае т. РНК) или путем присоединения полиаденилатного фрагмента (в случае м. РНК). В результате возникают функционально активные РНК.

Синтез РНК во многих отношениях подобен синтезу ДНК. Однако есть отличия механизмов синтеза РНК и ДНК: не нужна «затравка; n ДНК-матрица сохраняется; n РНК- полимераза не обладает нуклеазной активностью, т. е. не проверяет правильность новообразованной полинуклеотидной цепи. В связи с этим надёжность транскрипции значительно ниже, чем надёжность репликации. Это объясняется тем, что в клетке с 1 гена синтезируется много копий РНК. n Синтез и функционирование м. РНК у эукариот разделены во времени и пространстве: - синтез осуществляется в ядре, а функционирует м. РНК в цитоплазм; - между двумя этими событиями происходит процессинг (в ядре) и транспорт функционально зрелой м. РНК в цитоплазму. n

Пути распада НК В клетках происходит постоянный обмен нуклеотидов. Нуклеиновые кислоты в реакциях, катализируемых нуклеазами, распадаются на мононуклеотиды, которые подвергаются дальнейшему ферментативному гидролизу. Конечным продуктом гидролиза нуклеотидов являются моносахариды, которые включаются в реакции обмена, характерные для углеводов, и пуриновые и пиримидиновые основания.

Пурины расщепляются до мочевой кислоты, которая выводится из организма.

Пиримидины расщепляются до мочевины и аммиака. Сложные молекулы нуклеотидов в процессе распада в живых организмах превращаются в относительно простые соединения, которые либо выводятся из организма (аммиак, углекислый газ и др. ), либо участвуют в ресинтезе НК.
Обмен нуклеиновых кислот.ppt