ОБМЕН АММИАКА.ppt
- Количество слайдов: 61



ОБМЕН АММИАКА ИСТОЧНИКИ АММИАКА В КЛЕТКАХ

Катаболизм аминокислот в тканях происходит постоянно со скоростью 100 г/сут. При этом в результате дезаминирования аминокислот освобождается большое количество аммиака. Значительно меньшие количества его образуются при дезаминировании биогенных аминов и нуклеотидов.

Основные источники аммиака Источник Аминокислоты Процесс Ферменты Локализация процесса Непрямое дезаминирование (основной путь дезаминирования аминокислот) Аминотрансферазы, ПФ Глутаматдегидрогеназа, NAD+ Все ткани Окислительное дезаминирование глутамата Глутаматдегидрогеназа, NAD+ Все ткани Неокислительное дезаминирование Гис, Сер, Тре Гистидаза-Серин, треониндегидратазы, ПФ Преимущественно печень Окислительное дезаминирование аминокислот (малозначимый путь дезаминирования) Оксидаза Lаминокислот, FMN Печень и почки Биогенные амины Окислительное дезаминирование (путь инактивации биогенных аминов) Аминооксидазы, FAD Все ткани АМФ Гидролитическое дезаминирование АМФ-дезаминаза Интенсивно работающие мышцы

Часть аммиака образуется в кишечнике в результате действия бактерий на пищевые белки (гниение белков в кишечнике) и поступает в кровь воротной вены. Концентрация аммиака в крови воротной вены существенно больше, чем в общем кровотоке. В печени задерживается большое количество аммиака, что поддерживает низкое содержание его в крови. Концентрация аммиака в крови в норме редко превышает 0, 4 -0, 7 мг/л (или 25 -40 мкмоль/л).

В крови и цитозоле клеток при физиологических значениях р. Н аммиак переходит в ион аммония - NH 4+, количество неионизированного NH 3 невелико (~ 1%). Аммиак - токсичное соединение. Даже небольшое повышение его концентрации оказывает неблагоприятное действие на организм, и, прежде всего, на ЦНС.

Механизм токсического действия аммиака на мозг и организм в целом, очевидно, связан с действием его на несколько функциональных систем. Аммиак легко проникает через мембраны в клетки и в митохондриях сдвигает реакцию, катализируемую глутаматдегидрогеназой, в сторону образования глутамата: α-Кетоглутарат + NADH + Н+ + NH 3 → Глутамт + NAD+. Уменьшение концентрации α-кетоглутарата вызывает: • угнетение обмена аминокислот (реакции трансаминирования) и, следовательно, синтеза из них нейромедиаторов (ацетилхолина, дофамина и др. ); • гипоэнергетическое состояние в результате снижения скорости ЦТК.

Недостаточность α-кетоглутарата приводит к снижению концентрации метаболитов ЦТК, что вызывает ускорение реакции синтеза оксалоацетата из пирувата, сопровождающейся интенсивным потреблением СО 2. Усиленное образование и потребление диоксида углерода при гипераммониемии особенно характерны для клеток головного мозга.
. Это,")
Повышение концентрации аммиака в крови сдвигает р. Н в щелочную сторону (вызывает алкалоз). Это, в свою очередь, увеличивает сродство гемоглобина к кислороду, что приводит к гипоксии тканей, накоплению СО 2 и гипоэнергетическому состоянию, от которого главным образом страдает головной мозг. Высокие концентрации аммиака стимулируют синтез глутамина из глутамата в нервной ткани (при участии глутаминсинтетазы): Глутамат + NH 3 + АТФ → Глутамин + АДФ + Н 3 Р 04.

Накопление глутамина в клетках нейроглии приводит к повышению осмотического давления и в больших концентрациях может вызвать отёк мозга. Снижение концентрации глутамата нарушает обмен аминокислот и нейромедиаторов, в частности синтез Ɣ-аминомасляной кислоты (ГАМК), основного тормозного медиатора. При недостатке ГАМК и других медиаторов нарушается проведение нервного импульса, возникают судороги. Ион NH 4+ практически не проникает через цитоплазматические и митохондриальные мембраны. Избыток иона аммония в крови способен нарушать трансмембранный перенос одновалентных катионов Na+ и К+, конкурируя с ними за ионные каналы, что также влияет на проведение нервных импульсов.
 аммиака Высокая интенсивность процессов дезаминирования аминокислот в тканях и очень низкий уровень")
Связывание (обезвреживание) аммиака Высокая интенсивность процессов дезаминирования аминокислот в тканях и очень низкий уровень аммиака в крови свидетельствуют о том, что в клетках активно происходит связывание аммиака с образованием нетоксичных соединений, которые выводятся из организма с мочой. Эти реакции можно считать реакциями обезвреживания аммиака. В разных тканях и органах обнаружено несколько типов таких реакций. Универсальным механизмом обезвреживания аммиака, протекающей во всех тканях организма, является синтез глутамина под действием глутаминсинтетазы:

Глутаминсинтетаза обладает высоким сродством к аммиаку. Фермент локализован в митохондриях клеток, для его работы необходим кофактор - ионы Mg 2+. Глутамин легко переносится через клеточные мембраны путём облегчённой диффузии и поступает из тканей в кровь. Основными поставщиками глутамина являются мышцы, мозг и печень. С током крови глутамин транспортируется в кишечник и почки.

В клетках кишечника под действием фермента глутаминазы происходит гидролитическое освобождение амидного азота в виде аммиака:

Образовавшийся аммиак частично удаляется через кишечник, небольшая его часть через воротную вену попадает в печень, остальная часть поступает в почки и выводится в виде солей аммония. Глутамат подвергается трансаминированию с пируватом под действием аланинаминотрансферазы. αаминогруппа глутаминовой кислоты включается в состав аланина, который поступает из кишечника в кровь воротной вены и поглощается печенью.

В почках происходит гидролиз глутамина под действием глутаминазы с образованием аммиака. Этот процесс является одним из механизмов регуляции кислотно-щелочного равновесия в организме и сохранения важнейших катионов для поддержания осмотического давления. Глутаминаза почек значительно индуцируется при ацидозе, образующийся аммиак нейтрализует кислые продукты обмена и в виде аммонийных солей экскретируется с мочой. Таким образом, организм защищается от излишней потери ионов Na+ и К+, которые также могут использоваться для выведения анионов и утрачиваться. При алкалозе количество глутаминазы в почках снижается. В почках образуется и выводится около 0, 5 г солей аммония в сутки.

Высокий уровень глутамина в крови и лёгкость его поступления в клетки обусловливают использование глутамина во многих анаболических процессах. Глутамин - основной донор азота в организме. Амидный азот глутамина используется для синтеза пуриновых и пиримидиновых нуклеотидов, аспарагина, аминосахаров и других соединений.

Ещё одной реакцией обезвреживания аммиака в тканях можно считать синтез аспарагина под действием аспарагинсинтетазы.

В мозге обезвреживание аммиака происходит путем восстановительного аминирования α-кетоглутарата под действием глутаматдегидрогеназы. Однако эта реакция идет с незначительной скоростью, так как глутаматдегидрогеназа в тканях используется для непрямого дезаминирования глутамата. Хотя, если учитывать последующее образование глутамина, реакция выгодна для клеток, так как способствует связыванию сразу 2 молекул NH 3.

Из мышц и кишечника избыток аммиака выводится преимущественно в виде аланина. Этот механизм необходим, так как активность глутаматдегидрогеназы в мышцах невелика и непрямое дезаминирование аминокислот малоэффективно. Поэтому в мышцах существует ещё один путь выведения азота. Аминогруппы разных аминокислот посредством реакций трансаминирования переносятся на пируват, основным источником которого служит процесс окисления глюкозы.

Мышцы выделяют особенно много аланина в силу их большой массы, активного потребления глюкозы при физической работе, а также потому, что часть энергии они получают за счёт распада аминокислот. Образовавшийся аланин поступает в печень, где подвергается непрямому дезаминированию. Выделившийся аммиак обезвреживается, а пируват включается в глюконеогенез. Глюкоза из печени поступает в ткани и там, в процессе гликолиза, опять окисляется до пирувата.

Глюкозо-аланиновый цикл Образование аланина в мышцах, его перенос в печень и перенос глюкозы, синтезированной в печени, обратно в мышцы составляют глюкозо -аланиновый цикл.

Основной источник аммиака - аминокислоты. Большая часть образовавшегося аммиака обезвреживается в орнитиновом цикле в печени и выделяется в виде мочевины. Основной реакцией обезвреживания аммиака в тканях является синтез глутамина, который затем используется в анаболических процессах и для обезвреживания веществ в печени. Ферменты глутаматдегидрогеназа и глутаминсинтетаза являются регуляторными и обусловливают скорость процессов образования и обезвреживания аммиака.

Орнитиновый цикл

Мочевина - основной конечный продукт азотистого обмена, в составе которого из организма выделяется до 90% всего выводимого азота. Экскреция мочевины в норме составляет 25 г/сут. Количество азотсодержащих веществ в моче (%) при нормальном белковом питании.

Синтез мочевины представляет собой циклический процесс, состоящий из нескольких стадий, ключевым соединением которого, замыкающим цикл, является орнитин. Поэтому процесс синтеза мочевины получил название "орнитиновый цикл", или "цикл Кребса-Гензелейта".
 - полный амид угольной кислоты - содержит 2 атома")
Реакции синтеза мочевины Мочевина (карбамид) - полный амид угольной кислоты - содержит 2 атома азота. Источником одного из них является аммиак, который в печени связывается с диоксидом углерода с образованием карбамоилфосфата под действием карбамоилфосфатсинтетазы I.

Далее под действием орнитинкарбамоилтрансферазы карбамоильная группа карбамоилфосфата переносится на α-аминокислоту орнитин, и образуется другая αаминокислота - цитруллин.
. Этот")
В следующей реакции аргининосукцинатсинтетаза связывает цитруллин с аспартатом и образует аргининосукцинат (аргининоянтарную кислоту). Этот фермент нуждается в ионах Mg 2+. В реакции затрачивается 1 моль АТФ, но используется энергия двух макроэргических связей. Аспартат - источник второго атома азота мочевины.
 расщепляет аргининосукцинат на аргинин и фумарат, при этом аминогруппа аспартата")
Далее фермент аргининосукцинатлиаза (аргининосукциназа) расщепляет аргининосукцинат на аргинин и фумарат, при этом аминогруппа аспартата оказывается в молекуле аргинина.

Аргинин подвергается гидролизу под действием аргиназы, при этом образуются орнитин и мочевина. Кофакторами аргиназы являются ионы Са 2+ или Мn 2+. Высокие концентрации орнитина и лизина, являющихся структурными аналогами аргинина, подавляют активность этого фермента: Образующийся орнитин взаимодействует с новой молекулой карбамоилфосфата, и цикл замыкается.

Первые две реакции процесса происходят в митохондриях гепатоцитов. Затем цитруллин, являющийся продуктом этих реакций, транспортируется в цитозоль, где и осуществляются дальнейшие превращения.

Окислительное дезаминирование глутамата происходит в митохондриях. Ферменты орнитинового цикла распределены между митохондриями и цитозолем. Поэтому необходим трансмембранный перенос глутамата, цитруллина и орнитина с помощью специфических транслоказ.

")
На схеме показаны пути включения азота двух разных аминокислот (аминокислота 1 и аминокислота 2) в молекулу мочевины: • одна аминогруппа - в виде аммиака в матриксе митохондрии; • вторую аминогруппу поставляет аспартат цитозоля. Суммарное уравнение синтеза мочевины: СО 2 + NH 3 + Аспартат + 3 АТФ + 2 Н 2 О → Мочевина + Фумарат + 2 (АДФ + Н 3 Р 04) + АМФ + H 4 P 2 O 7.

Аммиак, используемый карбамоилфосфатсинтетазой I, поставляется в печень с кровью воротной вены. Аспартат, необходимый для синтеза аргининосукцината, образуется в печени путём трансаминирования аланина с оксалоацетатом. Аланин поступает главным образом из мышц и клеток кишечника. Источником оксалоацетата, необходимого для этой реакции, можно считать превращение фумарата, образующегося в реакциях орнитинового цикла. Фумарат в результате двух реакций цитратного цикла превращается в оксалоацетат, из которого путём трансаминирования образуется аспартат. Таким образом, с орнитиновым циклом сопряжён цикл регенерации аспартата из фумарата. Пируват, образующийся в этом цикле из аланина, используется для глюконеогенеза.
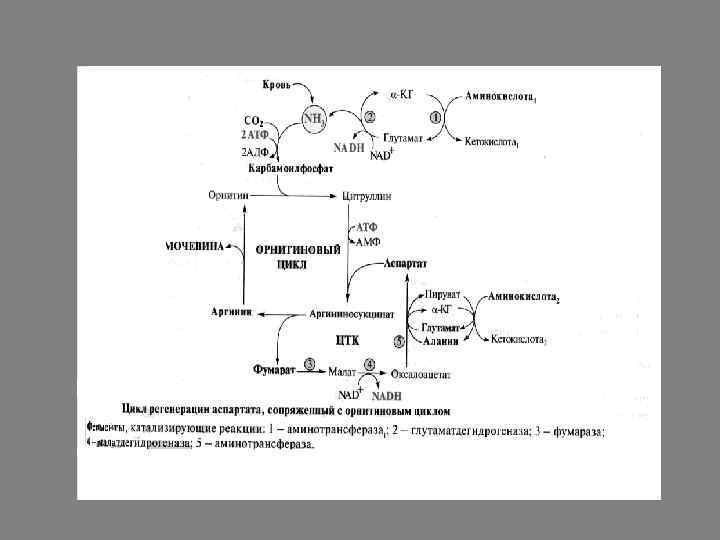

Энергетический баланс процесса В реакциях орнитинового цикла расходуются четыре макроэргических связи трёх молекул АТФ на каждый оборот цикла. Однако процесс превращения аминокислот в безазотистые остатки и мочевину имеет пути компенсации энергозатрат: • при включении фумарата в ЦТК на стадии дегидрирования малата образуется NADH, который обеспечивает синтез 3 молекул АТФ; • при окислительном дезаминировании глутамата в разных органах также образуется NADH, соответственно - ещё 3 молекулы АТФ.

Взаимосвязь орнитинового цикла и общего пути катаболизма. Фумарат, образующийся в результате расщепления аргининосукцината, превращается в малат, который затем переносится в митохондрии, включается в ЦТК и дегидрируется с образованием оксалоацетата. Эта реакция сопровождается выделением 3 молекул АТФ, которые и компенсируют затраты энергии на синтез одной молекулы мочевины.

Первые две реакции орнитинового цикла происходят в митохондриях, а последующие три - в цитозоле. Цитруллин, образующийся в митохондрии, должен быть перенесён в цитозоль, а орнитин, образующийся в цитозоле, необходимо транспортировать в митохондрию. Кроме того, в почках перенос мочевины из крови в мочу происходит путём активного транспорта за счёт градиента ионов натрия, создаваемого К+, Nа+АТФ-азой, что тоже сопряжено с энергозатратами.

Биологическая роль орнитинового цикла Кребса-Гензелейта Орнитиновый цикл в печени выполняет 2 функции: • превращение азота аминокислот в мочевину, которая экскретируется и предотвращает накопление токсичных продуктов, главным образом аммиака; • синтез аргинина и пополнение его фонда в организме.

Регуляторные стадии процесса - синтез карбамоилфосфата, синтез цитруллина и заключительная стадия, катализируемая аргиназой. Эффективность работы орнитинового цикла при нормальном питании человека и умеренных физических нагрузках составляет примерно 60% его мощности. Запас мощности необходим для избежания гипераммониемии при изменениях количества белка в пище. Увеличение скорости синтеза мочевины происходит при длительной физической работе или длительном голодании, которое сопровождается распадом тканевых белков.
, также сопровождаются активацией орнитинового")
Некоторые патологические состояния, характеризующиеся интенсивным распадом белков тканей (сахарный диабет), также сопровождаются активацией орнитинового цикла. При избыточном белковом питании количество ферментов орнитинового цикла в печени увеличивается, что приводит к интенсификации синтеза мочевины.
 или наследственный дефект ферментов обезвреживания аммиака могут вызвать повышение")
Гипераммониемия Заболевания печени (гепатит, цирроз) или наследственный дефект ферментов обезвреживания аммиака могут вызвать повышение содержания аммиака в крови - гипераммониемию. Известно пять наследственных заболеваний, обусловленных дефектом пяти ферментов орнитинового цикла.

Наследственные нарушения орнитинового цикла и основные их проявления Заболевание Дефект фермента Тип наследования Метаболиты Клинические проявления кровь моча Гипераммониемия, тип I Карбамоилфосфатсинтетаза I Аутосомнорецессивный В течение 24 -48 ч после рождения кома, смерть Глн Ала NH 3 Оротат Гипераммониемия, тип II Орнитинкарбамоилтрансфераза Сцепленный с Х -хромосомой Гипотония, снижение толерантности к белкам Глн Ала NH 3 Оротат Цитруллинемия Аргининосукцинатсинтетаза Аутосомнорецессивный Гипераммонием ия тяжёлая у новорождённых. У взрослых после белковой нагрузки Цитруллин NH 3 Цитрулли н Аргининосукцинатурия Аргининосукцинатлиаза Аутосомнорецессивный Гипераммоними я, атаксия, судороги, выпадение волос Аргининосукцинат NH 3 Аргининосукцинат, Глн, Ала, Лиз Гипераргининемия Аргиназа Аутосомнорецессивный Гипераргинине мия Apr NH 3 Apr Лиз Орнитин

При некоторых вирусных заболеваниях также наблюдается торможение орнитинового цикла (например, вирус гриппа ингибирует карбамоилфосфатсинтетазу I). Все нарушения орнитинового цикла приводят к значительному повышению в крови концентрации аммиака, глутамина и аланина. Гипераммониемия сопровождается появлением следующих симптомов: • тошнота, повторяющаяся рвота; • головокружение, судороги; • потеря сознания, отёк мозга (в тяжёлых случаях); • отставание умственного развития (при хронической врождённой форме). Все симптомы гипераммониемии - проявление действия аммиака на ЦНС.

Основной диагностический признак - повышение концентрации аммиака в крови. Содержание аммиака в крови может достигать 6000 мкмоль/л (в норме - 60 мкмоль/л). Однако в большинстве хронических случаев уровень аммиака может повышаться только после белковой нагрузки или в течение острых осложнённых заболеваний.

Лечение больных с различными дефектами орнитинового цикла в основном направлено на снижение концентрации аммиака в крови за счёт малобелковой диеты, введения кетоаналогов аминокислот в рацион и стимуляцию выведения аммиака. Вводимый больным с дефектом карбамоилфосфатсинтетазы I в качестве пищевой добавки фенилацетат в результате его конъюгации с глутамином образует фенилацетилглутамин, который экскретируется почками. Состояние больных при этом улучшается, так как происходит активация синтеза глутамина и снижение концентрации аммиака в крови.

Аналогичное действие оказывает введение бензоата, который связывает молекулу глицина. Образующаяся гиппуровая кислота выводится с мочой. В составе гиппуровой кислоты происходит выделение азота из организма. Недостаток глицина компенсируется либо путём синтеза его из серина, либо за счёт образования из NH 3 и СО 2 в реакции, катализируемой глицинсинтетазой. При этом образование глицина сопровождается связыванием одной молекулы аммиака.

Обмен аммиака и аминокислот между органами и тканями В катаболизме аминокислот и образовании аммиака участвуют многие ткани. В клетках происходит связывание аммиака. Из организма азот выводится почками в виде двух конечных продуктов азотистого обмена - аммонийных солей (~ 0, 5 г/сут), которые образуются в почках, и мочевины (~ 25 г/сут), которая содержит до 90% выводимого азота. Синтез мочевины происходит в печени в орнитиновом цикле, причём на образование 1 моля мочевины используется 1 моль аммиака и 1 моль аспарагиновой кислоты. Таким образом, для синтеза 25 г мочевины в сутки затрачивается 6, 3 г аммиака и 50 г аспартата. Для доставки азота в печень должны интенсивно функционировать специальные механизмы.

Транспорт азота из тканей в печень происходит, в основном, в составе 3 соединений: глутамина, аланина, аммиака. Кроме глутамина и аланина, в крови присутствуют и другие свободные аминокислоты, причём содержание их и направление транспорта зависят от приёма пищи и использования эндогенных белков. Наибольшее количество свободных аминокислот поступает из мышц и кишечника, причём до 50% составляют аланин и глутамин. Существует направленный поток аминокислот из этих тканей в печень, который усиливается в абсорбтивный период при белковом питании.

Основное количество глутамина поставляют в кровь мышцы и мозг. Из кровеносного русла его поглощают печень и почки, где он подвергается действию глутаминазы. Почки - основной источник серина и частично аланина, которые сорбируются из плазмы печенью. Головной мозг, в отличие от всех других тканей, способен поглощать и окислять большие количества аминокислот с разветвлённой боковой цепью (валин, лейцин, изолейцин).

После приёма пищи из кишечника в плазму крови поступает много аминокислот, причём преобладают аминокислоты с разветвлённой боковой цепью (до 20% от общего количества), которые затем поглощаются, в основном, печенью, мышцами и мозгом. В мышцах происходит усиленный катаболизм этих аминокислот, причём они выступают основными донорами аминогруппы в синтезе аланина из пирувата.

В абсорбтивный период основным источником свободных аминокислот служит кишечник. Большую часть поступивших аминокислот составляют гидрофобные аминокислоты с разветвлённой цепью. Экзогенные полярные аминокислоты из воротной вены сорбируются и используются в основном печенью. Разветвлённые аминокислоты поглощаются из кровотока клетками мозга или мышц.

В постабсорбтивном периоде основными источниками свободных аминокислот служат мышцы. Они поставляют в основном аланин и глутамин.

В постабсорбтивный период свободные аминокислоты поступают преимущественно из мышц, в которых усиливается катаболизм белков. Аминокислоты используются в глюконеогенезе в печени. В крови повышен уровень аланина, серина и глутамина. Аланин поглощается печенью, глутамин - кишечником и почками. В кишечнике азот глутамина переносится в аланин или серии и в их составе транспортируется в печень, где активируется процесс глюконеогенеза. Интенсивность глюконеогенеза из этих аминокислот намного выше, чем из всех других. Таким образом, аланин и серии - основные гликогенные аминокислоты. Аминокислоты с разветвлённой боковой цепью (валин, лейцин, изолейцин и др. ), которые освобождаются из мышц, направляются в мозг, где окисляются и служат важным

ПУТИ ОБМЕНА БЕЗАЗОТИСТОГО ОСТАТКА АМИНОКИСЛОТ

В процессе катаболизма аминокислоты превращаются в 6 метаболитов, вступающих в ОПК: пируват, ацетил-Ко. А, α-кетоглутарат, сукцинил-Ко. А, фумарат, оксалоацетат.
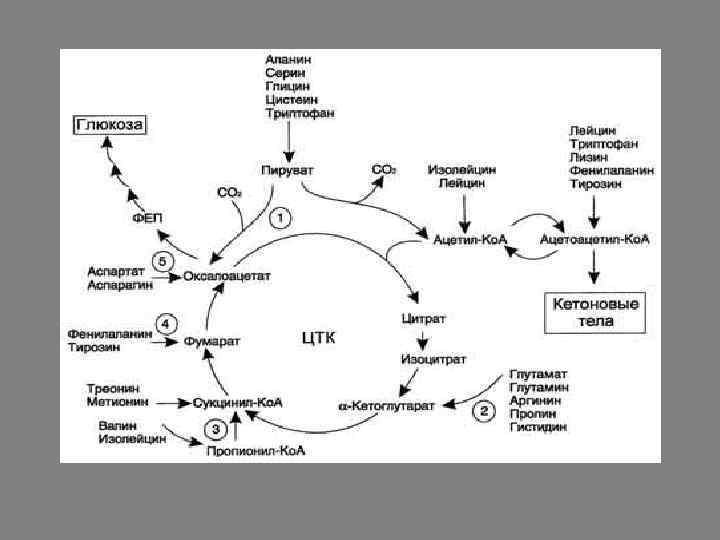

В дальнейшем эти метаболиты превращаются в глюкозу или образуют ацетоацетат. В зависимости от процесса, в который они включаются, аминокислоты можно разделить на 3 группы. 1. Гликогенные и кетонные аминокислоты 2. Кетогенные аминокислоты 3. Гликокетогенные аминокислоты

• Гликогенные аминокислоты образуют пируват и другие промежуточные продукты ЦТК (α-кетоглутарат, сукцинил-Ко. А, фумарат) превращаются в оксалоацетат и используются в процессе глюконеогенеза (ала, аспарагин, гли, глу, глутамин, про, сер, цис, арг, гис, вал, мет, тре). • Кетогенные аминокислоты (лиз, лей) превращаются в ацетоацетат или ацетил-Ко. А и могут использоваться в синтезе кетоновых тел. • Гликокетогенные аминокислоты распадаются на 2 продукта - определённый метаболит цитратного цикла и ацетоацетат (три, фен, тир) или ацетил-Ко. А (иле) и могут использоваться для синтеза как глюкозы, так и кетоновых тел.

Анаплеротические реакции Безазотистые остатки аминокислот используются для восполнения того количества метаболитов общего пути катаболизма, которое затрачивается на синтез биологически активных веществ. Такие реакции называют анаплеротическими. 1. Фермент пируваткарбоксилаза (кофермент - биотин), катализирующий эту реакцию, обнаружен в печени и мышцах. 2. Аминокислоты → Глутамат → α-Кетоглутарат Превращение происходит во многих тканях под действием глутаматдегидрогеназы или аминотрансфераз. 3. Пропионил-Ко. А, а затем и сукцинил-Ко. А могут образоваться также при распаде высших жирных кислот с нечётным числом атомов углерода.

4. Аминокислоты → Фумарат 5. Аминокислоты → Оксалоацетат Реакции 2, 3 происходят во всех тканях (кроме печени и мышц), где отсутствует пируваткарбоксилаза, а реакции 4 и 5 - в основном в печени. Реакции 1 и 3 - основные анаплеротические реакции.
ОБМЕН АММИАКА.ppt