лекция 1 мит.pptx
- Количество слайдов: 36



Нургалиева Альфия Хаматьяновна К. б. н. , асс.

Основная литература: 1. Васильев В. Б. Генетические основы митохондриальных болезней. СПб. : Издатель ство «Нестор История» , 2006. 146 с. 2. Иванов В. И. 2005. Геномика медицине [под ред. В. И. Иванова и Л. Л. Киселева]. М. : Академкнига; , 392 с. 3. Никитина Л. П. , Соловьева Н. В. и др. Митохондриальные болезни. Критерии и методы диагностики. // Забайкальсикий медицинский вестник, № 1, 2012 4. Сукерник Р. И. , Дербенека О. А. и др. Митохондриальный геном и митохондриальные болезни человека. // Генетика, Т. 38, № 2, 2002 Интернет ресурсы: 1. http: //www. mdausa. org/disease/mito. html 2. http: //neuromuscular. wustl. edu/mitosyn. html 3. www. ncbi. nih. gov

Митохондриа льные заболева ния — группа наследственных заболеваний, связанных с дефектами в функционировании митохондрий, приводящими к нарушениям энергетических функций в клетках эукариотов, в частности — человека.

• • Митохондриальные заболевания обусловлены генетическими, структурными, биохимическими дефектами митохондрий, приводящими к нарушениям тканевого дыхания. передаются только по женской линии к детям обоих полов, так как сперматозоиды передают зиготе половину ядерного генома, а яйцеклетка поставляет и вторую половину генома, и митохондрии Патологические нарушения клеточного энергетического обмена могут проявляться в виде дефектов различных звеньев в цикле Кребса, в дыхательной цепи, процессах бета-окисления и т. д. Не все ферменты и другие регуляторы, необходимые для эффективного функционирования митохондрий, кодируются митохондриальной ДНК. Большая часть митохондриальных функций контролируется ядерной ДНК.
 в виде")
Митохондрии — внутриклеточные органеллы, присутствующие в каждой клетке человека (кроме зрелых эритроцитов) в виде нескольких сотен копий. Их назначение — обеспечивать клетки энергией.

митохондрии, в отличие от других клеточных органелл, имеют свой собственный геном, представленный кольцевой хромосомой длиной в 16569 п. н.

Уникальные свойства митохондриального генома: - Цитоплазматическое наследование - Отсутствие рекомбинаций - Высокая скорость мутирования

ИСТОРИЯ ИССЛЕДОВАНИЯ МИТОХОНДРИЙ
![Первые наблюдения митохондрий в поперечно полосатых мышцах были сделаны Хенле [Henle, 1841] Чуть](https://present5.com/presentation/27845853_128955661/image-9.jpg "Первые наблюдения митохондрий в поперечно полосатых мышцах были сделаны Хенле [Henle, 1841] Чуть")
Первые наблюдения митохондрий в поперечно полосатых мышцах были сделаны Хенле [Henle, 1841] Чуть позже Кёлликер описал «интерстициальные гранулы» [Kolliker, 1856], а Альтманн — «бактериоподобных паразитов» [Altmann, 1890]. На сегодняшний день однозначно установлено, что эти органеллы эукариотических клеток, получившие свое название от греческих «mitos» — «нить» и «chondros» — «зерно» [Benda, 1898], ответственны за функцию тканевого дыхания и синтез аденозин-трифосфата (АТФ) [Lehninger and Kennedy, 1948; Cleland Slater, 1953], т. е. за феномен преобразования и запасания (консервации) клетками эукариотических организмов энергии, содержащейся в питательных субстратах.

Тканевое дыхание - последовательность окислительновосстановительных реакций в митохондриях, когда питательные субстраты в виде карбоксильных кислот окисляются так, что получаемые при этом электроны передаются по цепи из нескольких ферментных комплексов, расположенных во внутренней митохондриальной мембране В ходе исследований первой половины XX столетия было выяснено, что процесс консервации энергии происходит, благодаря сопряжению окисления и фосфорилирования [Engelhardt, 1930; Friedkin and Lehninger, 1948]. Эти исследования положили начало развитию очень важного направления в современной биологии — биоэнергетики.

В 1950 -е годы исследования митохондрий, в основном, сводились к изучению механизмов получения ими АТФ. 1961 год следует считать переломным моментом в истории биоэнергетики, благодаря появлению хемиосмотической гипотезы П. Митчелла [Mitchell, 1961]. Немедленного признания открытие Митчелла не получило, т. к. в начале 60 -х в биоэнергетике господствовала так называемая «химическая теория» консервации энергии [Racker, 1951; Slater, 1953; Chance and Williams, 1956]. Питер Митчелл

Хемиосмотическая гипотеза: Запасание высвобождаемой при окислении карбоксильных кислот энергии в митохондриях происходит в результате электрохимической активности протонов, создаваемой их перемещением из матрикса через внутреннюю мембрану в межмембранное пространство. Энергия для перемещения протонов через мембрану поступает от движения электронов от одного ферментного комплекса к другому, т. е. окислительно-восстановительного процесса

Схема дыхательной цепи митохондрий

Вопрос о происхождении митохондрий возник почти одновременно с интересом к их функциям. Идея об их самостоятельном воспроизводстве внутри клетки всегда была популярна среди исследователей хотя бы потому, что до начала 1920 -х годов многие естествоиспытатели рассматривали их как эндосимбиотические микроорганизмы. Эндосимбиотическая теория происхождения митохондрий, наиболее подробно разрабатывавшаяся Линн Маргулис [Margulis, 1970], имеет наибольшее число сторонников и сегодня.

В настоящее время эти органеллы считаются произошедшими от эндосимбионтных окисляющих бактерий, заселивших безмитохондриальные клетки, вероятно в то время, когда у эукариот формировалось ядро, или ранее [Gray et al, 1999; Lang etal. , 1999]. В связи с внутриклеточным существованием эндосимбионты претерпели серьезные изменения, однако и в нынешнем виде они сохраняют рудименты бактериальной структуры и функций — двойную мембрану, кольцевую ДНК и набор т. РНК и р. РНК, белки, позволяющие органеллам передвигаться, сливаться и расщепляться, множество реакций синтеза и распада — доставшихся сегодняшним митохондриям от предков.

ОРГАНИЗАЦИЯ МИТОХОНДРИАЛЬНОГО ГЕНОМА

- мт. ДНК человека представляет собой двойное кольцо, состоящее из 16569 нуклеотидных пар - в мт. ДНК млекопитающих отсутствуют интроны. гены примыкают вплотную один к другому или даже перекрываются - в геноме митохондрий нет гистонов, чем, в частности, объясняют более высокую частоту обнаружения мутаций в мт. ДНК по сравнению с ядерным геномом - В мт. ДНК человека обнаружено всего 37 генов

13 генов кодируют отдельные субъединицы комплексов дыхательной цепи митохондрий. Комплекс I в состав входят семь из 13 полипептидов (NADH -дегидрогеназы — коэнзим Q, оксидоредуктазы; по другой номенклатуре — NADH-убихинон оксидоредуктазы), Комплекс III входит один пп, представляет собой цитохром b(коэнзим Q — цитохром с оксидоредуктазы; по другой номенклатуре — убихинол цитохром с оксидоредуктазы), Комплекс IV – три пп (цитохром с оксидоредуктазы) Комплекс V – два пп (АТФ синтетазы) Следует отметить, что каждый из ферментных комплексов дыхательной цепи содержит и субъединицы, кодируемые в ядре. Сукцинатдегидрогеназа, образующая комплекс II, не имеет субъединиц, кодируемых в митохондриях.

Схема строения митохондриального генома

Остальные 24 гена мт. ДНК необходимы для трансляции белка на митохондриальных рибосомах (генетический код митохондрий несколько отличается от ядерного, поэтому синтез белка в них требует присутствия собственного вспомогательного аппарата). Из этих генов 22 кодируют т. РНК, а 2 — рибосомальные РНК митохондрий.

В мт. ДНК имеется некодирующий участок протяженностью в 1, 1 т. п. н. контрольная область или D-петля (displacement loop). Геном митохондрий в клетках млекопитающих организован очень компактно. Есть примеры неполных терминирующих кодонов (их достраивание достигается полиаденилированием м. РНК). D петля, единственный протяженный некодирующий участок, используется также весьма рационально — в ней расположены сайт инициации репликации тяжелой цепи мт. ДНК (H strand), точки начала транскрипции обеих цепей и последовательности, регулирующие репликацию и транскрипцию. Сайт инициации репликации легкой цепи мт. ДНК (L strand) находится в пределах кластера генов т. РНК, обозначаемого WANCY в соответствии с однобуквенными обозначениями аминокислот, связываемых этими т. РНК.

Карта некоторых наиболее распространенных точечных мутаций и гигантских делеций в мт. ДНК

В клетках млекопитающих присутствует много копий митохондриального генома — от нескольких сот в фибробластах мыши до примерно 105 в зрелой яйцеклетке, где мт. ДНК составляет до 1/з от общего количества ДНК. При этом каждая митохондрия содержит не одну, а несколько копий мт. ДНК. Большим числом копий мт. ДНК в значительной мере определяется сложный характер сегрегации и передачи митохондриального генома в поколениях клеток.

Молекулы мт. ДНК не распределены свободно и равномерно в пределах органеллы. Напротив, они объединены в небольшие кластеры по 2 -10 молекул. Такие кластеры получили название нуклеоидов. Было показано, что нуклеоиды прикреплены к внутренней митохондриальной мембране. О белках, обеспечивающих кластеризацию молекул мт. ДНК известно мало. Неясно и то, насколько стабильны нуклеоиды, и в какой мере постоянным является в них набор молекул мт. ДНК. Иными словами, нет ответа на вопрос, могут ли нуклеоиды обмениваться молекулами мт. ДНК, и, если да, то как часто происходят такие обмены.

Митохондриальный геном человека и других млекопитающих имеет ряд черт, необычных для генома ядерного. 1. В нем отсутствуют интроны, 2. генетический код отличается от ядерного 3. при транскрипции РНК гены, кодирующие т. РНК, используются в качестве «знаков пунктуации» 4. цепи мт. ДНК реплицируются асимметрично

Сравнение характеристик ядерного и митохондриального генома человека 9
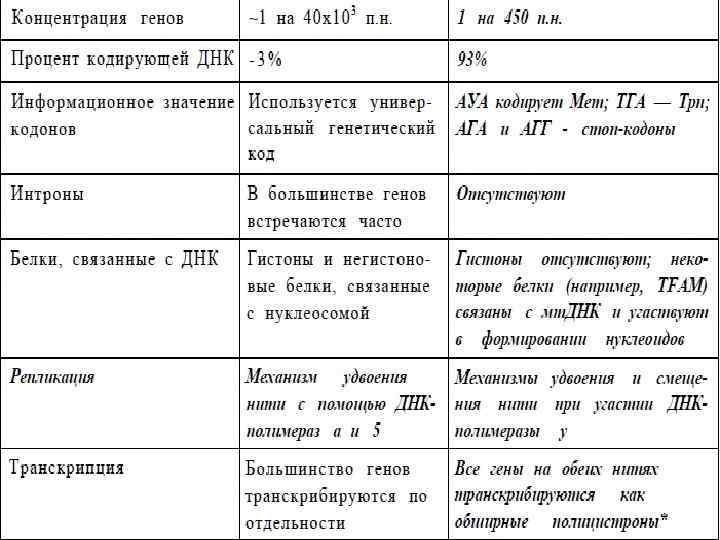


По сравнению с жестким контролем над репликацией и сегрегацией ядерной ДНК при клеточном делении, геном митохондрий находится в гораздо более свободных условиях. Впрочем, свобода репликации и сегрегации мт. ДНК — не абсолютная. Общее количество копий мт. ДНК на клетку остается постоянным для данной ткани после каждого цикла клеточного деления. Какие механизмы обеспечивают это постоянство, пока не вполне понятно. В репликации мт. ДНК, ее транскрипции и формировании нуклеоидов участвуют белки, кодируемые ядерной ДНК, и количество копий митохондриального генома в клетке может регулироваться в ходе разных процессов

Известно, что репликация мт. ДНК в клетках млекопитающих осуществляется преимущественно в митохондриях, расположенных близко к ядру. Скорее всего, это происходит из-за того, что факторы репликации, синтезируемые в ядре, в большей мере доступны для таких митохондрий. Вновь синтезированные молекулы мт. ДНК быстро распределяются по всей клетке, мигрируя по митохондриальной сети. Это наблюдение еще раз подчеркивает весьма относительную независимость митохондрий — будучи связанными с цитоскелетом, они не дрейфуют по цитоплазме; в итоге передача генетической информации дочерним органеллам также находится под влиянием клеточного ядра.

Генетические функции мт. ДНК сводятся к участию генома в 5 -6 процессах, протекающих в митохондриях. От генома зависят транскрипция РНК и ее созревание, трансляция кодируемых полипептидов и, наконец, импорт белков в митохондрии. Однако все эти функции мт. ДНК так или иначе сводятся к обеспечению тканевого дыхания и окислительного фосфорилирования. Последовательность нуклеотидов в мт. ДНК человека, впервые определенная группой Сэнгера [Anderson et al. , 1981], обозначается как Cambridge reference sequence (Кембриджская справочная последовательность) или CRS.





лекция 1 мит.pptx