

Лекция для аспирантов 1.ppt
- Количество слайдов: 75
 Нуклеозиды, нуклеотиды, нуклеиновые кислоты n n Вся информация о строении и функционировании любого клеточного живого организма содержится в закодированном виде в его генетическом материале, основу которого составляет дезоксирибонуклеиновая кислота (ДНК). ДНК большинства организмов — это длинная двухцепочечная полимерная молекула. Главная функция ДНК связана с ее участием в синтезе белков, аминокислотная последовательность которых определяет все множество процессов, составляющих основу жизни в течение миллионов лет ее эволюции.
Нуклеозиды, нуклеотиды, нуклеиновые кислоты n n Вся информация о строении и функционировании любого клеточного живого организма содержится в закодированном виде в его генетическом материале, основу которого составляет дезоксирибонуклеиновая кислота (ДНК). ДНК большинства организмов — это длинная двухцепочечная полимерная молекула. Главная функция ДНК связана с ее участием в синтезе белков, аминокислотная последовательность которых определяет все множество процессов, составляющих основу жизни в течение миллионов лет ее эволюции.
 История вопроса n n Индивидуальными генетическими элементами со строго специфичной нуклеотидной последовательностью, кодирующими определенные продукты, являются гены. Одни из них кодируют белки, другие -только молекулы РНК. 1860 г. Грегор Мендель выдвинул концепцию гена как неделимой функциональной единицы наследственности 1910 г. В. Йогансен ввел термин ген для обозначения гипотетической единицы информации, регулирующей наследование индивидуальных признаков организма
История вопроса n n Индивидуальными генетическими элементами со строго специфичной нуклеотидной последовательностью, кодирующими определенные продукты, являются гены. Одни из них кодируют белки, другие -только молекулы РНК. 1860 г. Грегор Мендель выдвинул концепцию гена как неделимой функциональной единицы наследственности 1910 г. В. Йогансен ввел термин ген для обозначения гипотетической единицы информации, регулирующей наследование индивидуальных признаков организма
 1953 год. Открытие века n Джеймс Уотсон и Френсис Крик обобщили накопленные к тому времени данные о составе и структуре ДНК, построив ставшую теперь классической модель двойной спирали ДНК. n n Все клеточные ДНК состоят из 2 -х комплементарных и антипараллельных полинуклеотидных цепей, закрученных вокруг общей оси с образованием двойной спирали. Последовательность мономерных единиц (дезоксирибонуклеотидов) в одной ее цепи соответствует (комплементарна) последовательности дезоксирибонуклеотидов в другой. Принцип комплементарности обеспечивает идентичность новосинтезированных молекул ДНК, образующихся при их удвоении (репликации), исходным молекулам.
1953 год. Открытие века n Джеймс Уотсон и Френсис Крик обобщили накопленные к тому времени данные о составе и структуре ДНК, построив ставшую теперь классической модель двойной спирали ДНК. n n Все клеточные ДНК состоят из 2 -х комплементарных и антипараллельных полинуклеотидных цепей, закрученных вокруг общей оси с образованием двойной спирали. Последовательность мономерных единиц (дезоксирибонуклеотидов) в одной ее цепи соответствует (комплементарна) последовательности дезоксирибонуклеотидов в другой. Принцип комплементарности обеспечивает идентичность новосинтезированных молекул ДНК, образующихся при их удвоении (репликации), исходным молекулам.
 Функции ДНК в клетке: n n n хранение воспроизводство передача генетической информации Центральная догма молекулярной биологии (предложена Ф. Криком) гласит передача генетической информации в клетке осуществляется по следующему пути: ДНК РНК Белок
Функции ДНК в клетке: n n n хранение воспроизводство передача генетической информации Центральная догма молекулярной биологии (предложена Ф. Криком) гласит передача генетической информации в клетке осуществляется по следующему пути: ДНК РНК Белок
 Структурные компоненты нуклеиновых кислот
Структурные компоненты нуклеиновых кислот
 Нуклеозиды
Нуклеозиды
 Нуклеотиды и нуклеиновые кислоты
Нуклеотиды и нуклеиновые кислоты
 Нуклеотиды
Нуклеотиды
 Двойная спираль ДНК в В-форме
Двойная спираль ДНК в В-форме
 ВВЕДЕНИЕ В ТЕХНОЛОГИЮ РЕКОМБИНАНТНЫХ ДНК n Молекулярная генетика – область знаний, предметом изучения которой являются генетические механизмы клетки n Генетические механизмы клетки включают: q - перенос генетических признаков (репликация) q экспрессия генов (транскрипция и трансляция) q - перераспределение генов – обновление генома (генетическая рекомбинация, мутации)
ВВЕДЕНИЕ В ТЕХНОЛОГИЮ РЕКОМБИНАНТНЫХ ДНК n Молекулярная генетика – область знаний, предметом изучения которой являются генетические механизмы клетки n Генетические механизмы клетки включают: q - перенос генетических признаков (репликация) q экспрессия генов (транскрипция и трансляция) q - перераспределение генов – обновление генома (генетическая рекомбинация, мутации)
 Репликация
Репликация
 Биологическая роль нуклеиновых кислот n n Генетическая программа клеточных организмов записана в нуклеотидной последовательности ДНК, поэтому для сохранения уникальных свойств организма необходимо точное воспроизведение этой последовательности в каждом последующем поколении. Размер ДНК прокариот около 4 х106 пар нуклеотидов Размер ДНК эукариот около 4 х109 пар нуклеотидов Уникальной особенностью ДНК является то, что в ней закодирована информация о механизме ее собственного удвоения: одни гены кодируют ферменты, синтезирующие последовательности ДНК, другие – белки, осуществляющие сборку активированных нуклеотидов в полинуклеотидную последовательность
Биологическая роль нуклеиновых кислот n n Генетическая программа клеточных организмов записана в нуклеотидной последовательности ДНК, поэтому для сохранения уникальных свойств организма необходимо точное воспроизведение этой последовательности в каждом последующем поколении. Размер ДНК прокариот около 4 х106 пар нуклеотидов Размер ДНК эукариот около 4 х109 пар нуклеотидов Уникальной особенностью ДНК является то, что в ней закодирована информация о механизме ее собственного удвоения: одни гены кодируют ферменты, синтезирующие последовательности ДНК, другие – белки, осуществляющие сборку активированных нуклеотидов в полинуклеотидную последовательность
 Основные понятия n n Репликон – генетическая единица репликации, т. е. сегмент ДНК, который автономно воспроизводится (т. е. реплицируется) в процессе клеточного роста и деления. Напр. , хромосома E. coli, плазмиды, ДНК бактериофагов представляют собой репликоны разной сложности. Каждый репликон должен иметь «систему управления» собственной репликацией. Это - совокупность всех белковых участников процесса репликации. Репликон может содержать в себе гены, кодирующие синтез всех белков, необходимых для репликации (хромосома E. coli), части таких белков (некоторые сравнительно крупные бактериофаги) или использовать для своей репликации практически только чужие белки (мелкие фаги М 13 и G 4, содержащие однонитевые циклические ДНК).
Основные понятия n n Репликон – генетическая единица репликации, т. е. сегмент ДНК, который автономно воспроизводится (т. е. реплицируется) в процессе клеточного роста и деления. Напр. , хромосома E. coli, плазмиды, ДНК бактериофагов представляют собой репликоны разной сложности. Каждый репликон должен иметь «систему управления» собственной репликацией. Это - совокупность всех белковых участников процесса репликации. Репликон может содержать в себе гены, кодирующие синтез всех белков, необходимых для репликации (хромосома E. coli), части таких белков (некоторые сравнительно крупные бактериофаги) или использовать для своей репликации практически только чужие белки (мелкие фаги М 13 и G 4, содержащие однонитевые циклические ДНК).
 Стадии процесса репликации n n n Инициация - в сайте ori C (origion replication inducing) (олигонуклеотидная последовательность, характерная для каждого конкретного вида живого мира) происходит узнавание и связывание инициаторного сегмента ДНК с белком хеликазой. Хеликаза расплетает цепи за счет энергии АТФ. Элонгация рост олигонуклеотидной цепи. Осуществляется путем отбора ДНК-зависимой-ДНКполимеразой нуклеотидтрифосфатного субстрата по принципу комплементарности нуклеотиду, ближайшему на матричной (родительской) цепи, и последующее создание фосфодиэфирных связей между последним и вновь введенным нуклеотидами. Терминация завершение синтеза в результате считывания всей последовательности ДНК.
Стадии процесса репликации n n n Инициация - в сайте ori C (origion replication inducing) (олигонуклеотидная последовательность, характерная для каждого конкретного вида живого мира) происходит узнавание и связывание инициаторного сегмента ДНК с белком хеликазой. Хеликаза расплетает цепи за счет энергии АТФ. Элонгация рост олигонуклеотидной цепи. Осуществляется путем отбора ДНК-зависимой-ДНКполимеразой нуклеотидтрифосфатного субстрата по принципу комплементарности нуклеотиду, ближайшему на матричной (родительской) цепи, и последующее создание фосфодиэфирных связей между последним и вновь введенным нуклеотидами. Терминация завершение синтеза в результате считывания всей последовательности ДНК.
 Синтез комплементарной цепи ДНК
Синтез комплементарной цепи ДНК
 Репликация ДНК в E. coli
Репликация ДНК в E. coli
 Схема репликации хроматина в эукариотической клетке
Схема репликации хроматина в эукариотической клетке
 Отличие репликации у прокариот и эукариот Прокариоты Инициация Скорость Размер фрагментов Оказаки Эукариоты В одной точке Множественная инициация Высокая: прибл. 1500 п. о. в сек. Низкая: прибл. 50 -100 п. о. в сек (но множественная инициация компенсирует ее и Р. заканчивается в разумное время) 1000 -2000 п. о. 100 -200 п. о.
Отличие репликации у прокариот и эукариот Прокариоты Инициация Скорость Размер фрагментов Оказаки Эукариоты В одной точке Множественная инициация Высокая: прибл. 1500 п. о. в сек. Низкая: прибл. 50 -100 п. о. в сек (но множественная инициация компенсирует ее и Р. заканчивается в разумное время) 1000 -2000 п. о. 100 -200 п. о.
 Репликация одноцепочечных вирусных ДНК. В –вирусный геном, К – его комплемент
Репликация одноцепочечных вирусных ДНК. В –вирусный геном, К – его комплемент
, расшифровывается") ЭКСПРЕССИЯ ГЕНА n n Информация, содержащаяся в генах, которые кодируют белки (структурных генах), расшифровывается в ходе экспрессии гена- двух последовательных процессов: транскрипции -( синтеза м. РНК) и трансляции (синтеза белка). Сначала на определенном участке ДНК как на матрице синтезируется матричная РНК (м. РНК). Затем в ходе согласованной работы многокомпонентной системы при участии транспортных РНК (т. РНК), м. РНК, ферментов и различных белковых факторов осуществляется синтез белковой молекулы. Все эти процессы обеспечивают правильный перевод зашифрованной в ДНК генетической информации с языка нуклеотидов на язык аминокислот. Аминокислотная последовательность белковой молекулы однозначно задает ее структуру и функции.
ЭКСПРЕССИЯ ГЕНА n n Информация, содержащаяся в генах, которые кодируют белки (структурных генах), расшифровывается в ходе экспрессии гена- двух последовательных процессов: транскрипции -( синтеза м. РНК) и трансляции (синтеза белка). Сначала на определенном участке ДНК как на матрице синтезируется матричная РНК (м. РНК). Затем в ходе согласованной работы многокомпонентной системы при участии транспортных РНК (т. РНК), м. РНК, ферментов и различных белковых факторов осуществляется синтез белковой молекулы. Все эти процессы обеспечивают правильный перевод зашифрованной в ДНК генетической информации с языка нуклеотидов на язык аминокислот. Аминокислотная последовательность белковой молекулы однозначно задает ее структуру и функции.
 Транскрипция
Транскрипция
 Этапы транскрипции n n n Инициация – узнавание ДНК-зависимой-РНК-полимеразой характерной олигонуклеотидной последовательности в транскрибируемой цепи ДНК– оператора, образование нуклеиново-белкового комплекса Элонгация - рост олигонуклеотидной цепи. Осуществляется по аналогии с репликацией путем отбора ДНК-зависимой-РНКполимеразой нуклеотидтрифосфатного субстрата по принципу комплементарности нуклеотиду, ближайшему на матричной (родительской) цепи, и последующее создание фосфодиэфирных связей между последним и вновь введенным нуклеотидами. Терминация - завершение синтеза м. РНК на терминаторном сайте в результате считывания всей последовательности гена и разрушения нуклеиново-белкового комплекса за счет образования терминаторной «шпильки» .
Этапы транскрипции n n n Инициация – узнавание ДНК-зависимой-РНК-полимеразой характерной олигонуклеотидной последовательности в транскрибируемой цепи ДНК– оператора, образование нуклеиново-белкового комплекса Элонгация - рост олигонуклеотидной цепи. Осуществляется по аналогии с репликацией путем отбора ДНК-зависимой-РНКполимеразой нуклеотидтрифосфатного субстрата по принципу комплементарности нуклеотиду, ближайшему на матричной (родительской) цепи, и последующее создание фосфодиэфирных связей между последним и вновь введенным нуклеотидами. Терминация - завершение синтеза м. РНК на терминаторном сайте в результате считывания всей последовательности гена и разрушения нуклеиново-белкового комплекса за счет образования терминаторной «шпильки» .
 Последовательное копирования одной из цепей ДНК
Последовательное копирования одной из цепей ДНК
 +
+
 Схема синтеза РНК с помощью ДНК-зависимой РНК-полимеразы
Схема синтеза РНК с помощью ДНК-зависимой РНК-полимеразы
 Формирование транскрипционных шпилек
Формирование транскрипционных шпилек
 ρ-зависимая терминация
ρ-зависимая терминация
 Процессинг
Процессинг
 Схема процессинга пре-м. РНК в ядре
Схема процессинга пре-м. РНК в ядре
 Участие отечественных ученых в познании механизмов транскрипции n Свердлов Евгений Давидович, академик РАН, зав. кафедрой молекулярной биологии МГУ им. М. В. Ломоносова – установление первичной структуры α, β, β‘ – субъединиц РНК-полимеразы E. coli n Георгиев Георгий Павлович, академик РАН, директор инта Биологии гена РАН – изучение информомер – комплексов м-РНК с белками в клеточном ядре n Спирин Александр Сергеевич, академик РАН, основатель ин-та Белка РАН в в Пущино - изучение информосом комплексов м-РНК с белками в цитоплазме
Участие отечественных ученых в познании механизмов транскрипции n Свердлов Евгений Давидович, академик РАН, зав. кафедрой молекулярной биологии МГУ им. М. В. Ломоносова – установление первичной структуры α, β, β‘ – субъединиц РНК-полимеразы E. coli n Георгиев Георгий Павлович, академик РАН, директор инта Биологии гена РАН – изучение информомер – комплексов м-РНК с белками в клеточном ядре n Спирин Александр Сергеевич, академик РАН, основатель ин-та Белка РАН в в Пущино - изучение информосом комплексов м-РНК с белками в цитоплазме
 Трансляция
Трансляция
 Генетический код
Генетический код
 Правила генетического кода n Последовательность нуклеотидов молекулы м-РНК –посредника при передаче информации от ДНК к белку – считывается по порядку группами из 3 -х нуклеотидов n Каждой аминокислоте соответствует триплет нуклеотидов (кодон) n Каждый кодон кодирует только одну аминокислоту n Последовательность кодонов в м-РНК определяет аминокислотную последовательность синтезируемого белка
Правила генетического кода n Последовательность нуклеотидов молекулы м-РНК –посредника при передаче информации от ДНК к белку – считывается по порядку группами из 3 -х нуклеотидов n Каждой аминокислоте соответствует триплет нуклеотидов (кодон) n Каждый кодон кодирует только одну аминокислоту n Последовательность кодонов в м-РНК определяет аминокислотную последовательность синтезируемого белка
 Строение т-РНК Слева представлен механизм спаривания оснований в соответствующих участках молекулы (структура «клеверного листа» ), а справа - общая трехмерная конформация молекулы, установленная с помощью дифракции рентгеновских лучей. Отметим, что молекула напоминает по форме букву L, один ее конец (акцепторный) предназначен для присоединения аминокислоты, а второй содержит антикодон, состоящий из трех нуклеотидов. Аминокислота присоединяется к остатку А последовательности ССА на 3'-конце молекулы
Строение т-РНК Слева представлен механизм спаривания оснований в соответствующих участках молекулы (структура «клеверного листа» ), а справа - общая трехмерная конформация молекулы, установленная с помощью дифракции рентгеновских лучей. Отметим, что молекула напоминает по форме букву L, один ее конец (акцепторный) предназначен для присоединения аминокислоты, а второй содержит антикодон, состоящий из трех нуклеотидов. Аминокислота присоединяется к остатку А последовательности ССА на 3'-конце молекулы
 Реакция аминоацилирования т-РНК
Реакция аминоацилирования т-РНК
 Белковые факторы, участвующие в трансляции Стадии трансляции Инициация Прокариоты Факторы IF-3 IF-1 Элонгация Терминация Действие Мономерные белки IF-2 EF-Tu (unstable) EF-Ts (stable) EF-G RF-1 RF-2 RF-3 Эукариоты EIF 1 -8 Катализирует р-цию связывания т-РНК с рибосомой (30 S субъединица) Катализирует р-цию связывания м-РНК с рибосомой Стимулирует обе реакции, но играет подчиненную роль Состоят из неск. субъединиц, так EIF 2 состоит из 9 субъединиц Переносит аминоацил-т-РНК на рибосому EF 1 (аналог EF-Tu) Регенерирует активную форму Tu через комплекс Tu-Ts участвует в транслокации, т. е. в перемещении рибосом от кодона к кодону м-РНК EF 2 (аналог EF-G) Отщепляет пептидную цепь для терминирующих кодонов UAA и UAG Отщепляет пептидную цепь для терминирующих кодонов UAA и UGA Не имеет собственной каталитической активности, но стимулирует действие факторов RF-1 и RF-2 RF-фактор У млекопитающих один Узнает все три терминирующих кодона
Белковые факторы, участвующие в трансляции Стадии трансляции Инициация Прокариоты Факторы IF-3 IF-1 Элонгация Терминация Действие Мономерные белки IF-2 EF-Tu (unstable) EF-Ts (stable) EF-G RF-1 RF-2 RF-3 Эукариоты EIF 1 -8 Катализирует р-цию связывания т-РНК с рибосомой (30 S субъединица) Катализирует р-цию связывания м-РНК с рибосомой Стимулирует обе реакции, но играет подчиненную роль Состоят из неск. субъединиц, так EIF 2 состоит из 9 субъединиц Переносит аминоацил-т-РНК на рибосому EF 1 (аналог EF-Tu) Регенерирует активную форму Tu через комплекс Tu-Ts участвует в транслокации, т. е. в перемещении рибосом от кодона к кодону м-РНК EF 2 (аналог EF-G) Отщепляет пептидную цепь для терминирующих кодонов UAA и UAG Отщепляет пептидную цепь для терминирующих кодонов UAA и UGA Не имеет собственной каталитической активности, но стимулирует действие факторов RF-1 и RF-2 RF-фактор У млекопитающих один Узнает все три терминирующих кодона
 Стадия инициации. Образование инициаторного комплекса у прокариот
Стадия инициации. Образование инициаторного комплекса у прокариот
 Стадия элонгации. Синтез первой пептидной связи. Химизм процесса
Стадия элонгации. Синтез первой пептидной связи. Химизм процесса
 Стадия элонгации. Синтез первой пептидной связи
Стадия элонгации. Синтез первой пептидной связи
 Терминация белкового синтеза
Терминация белкового синтеза
 Полирибосома
Полирибосома
 Сравнительный анализ механизмов репликации, транскрипции и трансляции Механизм Репликация Транскрипция Трансляция матричный полуконсервативный матричный переход с нуклеотидного на аминокислотный язык прочтения информации Считывание информации от 3' - к 5' -концу от 5' - к 3' -концу Инициация осущ-ся в т. ori ос-ся на промоторе холо-фер-том осущ-ся на AUGкодоне Элонгация осущ-ся ДНКполимеразой осущ-ся корферментом осущ-ся мультиферментным комплексом Терминация по окончании считывания всей ДНК на терминаторе каждого гена (П, Э) или оперона (П) на стоп-кодонах: UAA, UAG, UGA Участие матрицы
Сравнительный анализ механизмов репликации, транскрипции и трансляции Механизм Репликация Транскрипция Трансляция матричный полуконсервативный матричный переход с нуклеотидного на аминокислотный язык прочтения информации Считывание информации от 3' - к 5' -концу от 5' - к 3' -концу Инициация осущ-ся в т. ori ос-ся на промоторе холо-фер-том осущ-ся на AUGкодоне Элонгация осущ-ся ДНКполимеразой осущ-ся корферментом осущ-ся мультиферментным комплексом Терминация по окончании считывания всей ДНК на терминаторе каждого гена (П, Э) или оперона (П) на стоп-кодонах: UAA, UAG, UGA Участие матрицы
 Проблемы нуклеиново-белкового узнавания n n n Узнавание нуклеиновой кислоты белком представляет собой процесс, каждая стадия которого осуществляется за счет нуклеиново-белковых взаимодействий. Под специфическим нуклеиново-белковым взаимодействием подразумевается взаимодействие определенных групп белка и нуклеиновой к-ты, возникающее за счет характерного для данного белка и данной нуклеиновой к-ты пространственного расположения этих групп. Примеры: репрессоры и операторы, РНК-полимеразы и промоторы. Неспецифические взаимодействия обусловлены электростатическими и гидрофобными связями. Примеры: РНК-полимераза образует неспецифические комплексы с любыми полианионами – гепарином, полиинозиновой кислотой, одноцепочечной и двухцепочечной ДНК.
Проблемы нуклеиново-белкового узнавания n n n Узнавание нуклеиновой кислоты белком представляет собой процесс, каждая стадия которого осуществляется за счет нуклеиново-белковых взаимодействий. Под специфическим нуклеиново-белковым взаимодействием подразумевается взаимодействие определенных групп белка и нуклеиновой к-ты, возникающее за счет характерного для данного белка и данной нуклеиновой к-ты пространственного расположения этих групп. Примеры: репрессоры и операторы, РНК-полимеразы и промоторы. Неспецифические взаимодействия обусловлены электростатическими и гидрофобными связями. Примеры: РНК-полимераза образует неспецифические комплексы с любыми полианионами – гепарином, полиинозиновой кислотой, одноцепочечной и двухцепочечной ДНК.
 Регуляция экспрессии гена
Регуляция экспрессии гена
 n n n В живых клетках действуют точно запрограммированные механизмы, обеспечивающие «правильные» соотношения между количествами различных синтезированных белков. Так, число молекул ферментов, катализирующих реакции основного метаболического пути, должны значительно превышать число молекул ферментов, ответственных за биосинтез коферментов, необходимых лишь в следовых количествах. Было показано, что клетки про- и эукариот обладают способностью к дифференциальной регуляции экспрессии генов. При определенных условиях многие гены вообще не экспрессируются, а степень экспрессии других различается на несколько порядков. Изменение внешних условий может привести к активации «молчавших» ранее генов и репрессии активно работавших. Подобная способность позволяет клеткам приспособить свои фенотипы к самым разнообразным условиям окружающей среды. Механизмы регуляции экспрессии генов весьма многообразны, многочисленны и очень сложны. И хотя многим из них присущи общие черты, тонкие механизмы регуляции всегда уникальны для - данного гена - определенного физиологического состояния организма и - условий окружающей среды
n n n В живых клетках действуют точно запрограммированные механизмы, обеспечивающие «правильные» соотношения между количествами различных синтезированных белков. Так, число молекул ферментов, катализирующих реакции основного метаболического пути, должны значительно превышать число молекул ферментов, ответственных за биосинтез коферментов, необходимых лишь в следовых количествах. Было показано, что клетки про- и эукариот обладают способностью к дифференциальной регуляции экспрессии генов. При определенных условиях многие гены вообще не экспрессируются, а степень экспрессии других различается на несколько порядков. Изменение внешних условий может привести к активации «молчавших» ранее генов и репрессии активно работавших. Подобная способность позволяет клеткам приспособить свои фенотипы к самым разнообразным условиям окружающей среды. Механизмы регуляции экспрессии генов весьма многообразны, многочисленны и очень сложны. И хотя многим из них присущи общие черты, тонкие механизмы регуляции всегда уникальны для - данного гена - определенного физиологического состояния организма и - условий окружающей среды
 Факторы регуляции экспрессии гена n n n Регуляция на уровне транскрипции - стадии инициации -- с помощью регуляторных белков --- активаторных, необходимых для ее начала - индуцибельный тип регуляции --- репрессорных, предотвращающих транскрипцию – репрессибельный тип регуляции -- с помощью «сильных» и «слабых» промоторов Эффективность инициации транскрипции (количество молекул синтезированной м. РНК) зависит от частоты актов инициации, которая определяется соответствием (аффинностью) пространственной структуры участников процесса: ДНК-зависимой-РНК-полимеразы и олигонуклеотидной последовательности оператора структурного гена. «Сильный» промотор, обладающий большей аффинностью к ферменту, обеспечивает и большую частоту инициации транскрипции. «Слабый» промотор менее продуктивен. - стадии элонгации -- контроль скорости элонгации - стадии терминации -- с помощью аттенуации ( «стоп-сигнал» в транскрибируемой последовательности)
Факторы регуляции экспрессии гена n n n Регуляция на уровне транскрипции - стадии инициации -- с помощью регуляторных белков --- активаторных, необходимых для ее начала - индуцибельный тип регуляции --- репрессорных, предотвращающих транскрипцию – репрессибельный тип регуляции -- с помощью «сильных» и «слабых» промоторов Эффективность инициации транскрипции (количество молекул синтезированной м. РНК) зависит от частоты актов инициации, которая определяется соответствием (аффинностью) пространственной структуры участников процесса: ДНК-зависимой-РНК-полимеразы и олигонуклеотидной последовательности оператора структурного гена. «Сильный» промотор, обладающий большей аффинностью к ферменту, обеспечивает и большую частоту инициации транскрипции. «Слабый» промотор менее продуктивен. - стадии элонгации -- контроль скорости элонгации - стадии терминации -- с помощью аттенуации ( «стоп-сигнал» в транскрибируемой последовательности)
 ГИПОТЕЗА ЖАКОБА И МОНО, поясняющая функционирование индуцибельных и репрессибельных систем n n n n n Французские ученые Жакоб и Моно (60 -е годы ХХ века) - репрессия ферментов – основной процесс - индукция ферментов достигается снятием репрессии - регуляторный ген (R-ген) кодирует аминокислотную последовательность белка -репрессора, который образует специфический комплекс с оператором структурного гена, контролируемый репрессором. Связывание репрессора с ДНК препятствует транскрипции соответствующего структурного гена. - молекула репрессора содержит сайт связывания, специфичный для участка ДНК, названного оператором, и расположенного перед структурным геном соответствующего фермента - для индуцибельных систем репрессия снимается при появлении индуктора, способного связаться с репрессором - поэтому молекула репрессора содержит два специфических центра связывания: один для оператора, другой для индуктора - для репрессибельных систем сам репрессор неактивен и репрессия осуществляется уже комплексом репрессора с корепрессором, которым является конечный продукт метаболического пути - гипотеза пригодна и для случая координированной индукции и репрессии, при которых один индуктор или репрессор могут регулировать синтез нескольких ферментов
ГИПОТЕЗА ЖАКОБА И МОНО, поясняющая функционирование индуцибельных и репрессибельных систем n n n n n Французские ученые Жакоб и Моно (60 -е годы ХХ века) - репрессия ферментов – основной процесс - индукция ферментов достигается снятием репрессии - регуляторный ген (R-ген) кодирует аминокислотную последовательность белка -репрессора, который образует специфический комплекс с оператором структурного гена, контролируемый репрессором. Связывание репрессора с ДНК препятствует транскрипции соответствующего структурного гена. - молекула репрессора содержит сайт связывания, специфичный для участка ДНК, названного оператором, и расположенного перед структурным геном соответствующего фермента - для индуцибельных систем репрессия снимается при появлении индуктора, способного связаться с репрессором - поэтому молекула репрессора содержит два специфических центра связывания: один для оператора, другой для индуктора - для репрессибельных систем сам репрессор неактивен и репрессия осуществляется уже комплексом репрессора с корепрессором, которым является конечный продукт метаболического пути - гипотеза пригодна и для случая координированной индукции и репрессии, при которых один индуктор или репрессор могут регулировать синтез нескольких ферментов
 Индуцибельный тип регуляции Структура lac-оперона E. сoli
Индуцибельный тип регуляции Структура lac-оперона E. сoli
 Репрессибельный тип регуляции Триптофановый оперон
Репрессибельный тип регуляции Триптофановый оперон
 Аттенуация
Аттенуация
 экспрессии триптофанового оперона n n n Аттенуатор представляет собой нуклеотидную последовательность, содержащую") Аттенуация (ослабление) экспрессии триптофанового оперона n n n Аттенуатор представляет собой нуклеотидную последовательность, содержащую команду, по которой происходит преждевременная терминация транскрипции. Аттенуатор содержит 3 сегмента: - 1 -й сегмент кодирует короткий полипептид из 14 аминокислот, две из кото-рых являются тандемными триптофановыми остатками - 2 -й и 3 -й сегменты включают инвертированные повторы, из-за чего проис-ходит внутримолекулярное спаривание оснований РНКтранскрипта и образование двух альтернативных шпилечных структур. Одна из них, в которой спарены 1+2 и 3+4 участки, является терминирующей, другая, в которой спарены 2+3 участки, является разрешающей. Какая именно шпилечная структура образуется при транскрипции зависит от уровня содержания триптофана в клетке. Терминация происходит при уровнях от среднего до высокого. При низком уровне терминация блокируется и транскрипция проходит через аттенуатор с образованием полноразмерной м. РНК триптофанового оперона. На аттенуацию влияет трансляция, идущая одновременно с транскрипцией.
Аттенуация (ослабление) экспрессии триптофанового оперона n n n Аттенуатор представляет собой нуклеотидную последовательность, содержащую команду, по которой происходит преждевременная терминация транскрипции. Аттенуатор содержит 3 сегмента: - 1 -й сегмент кодирует короткий полипептид из 14 аминокислот, две из кото-рых являются тандемными триптофановыми остатками - 2 -й и 3 -й сегменты включают инвертированные повторы, из-за чего проис-ходит внутримолекулярное спаривание оснований РНКтранскрипта и образование двух альтернативных шпилечных структур. Одна из них, в которой спарены 1+2 и 3+4 участки, является терминирующей, другая, в которой спарены 2+3 участки, является разрешающей. Какая именно шпилечная структура образуется при транскрипции зависит от уровня содержания триптофана в клетке. Терминация происходит при уровнях от среднего до высокого. При низком уровне терминация блокируется и транскрипция проходит через аттенуатор с образованием полноразмерной м. РНК триптофанового оперона. На аттенуацию влияет трансляция, идущая одновременно с транскрипцией.
 Лидерная trp-м. РНК
Лидерная trp-м. РНК
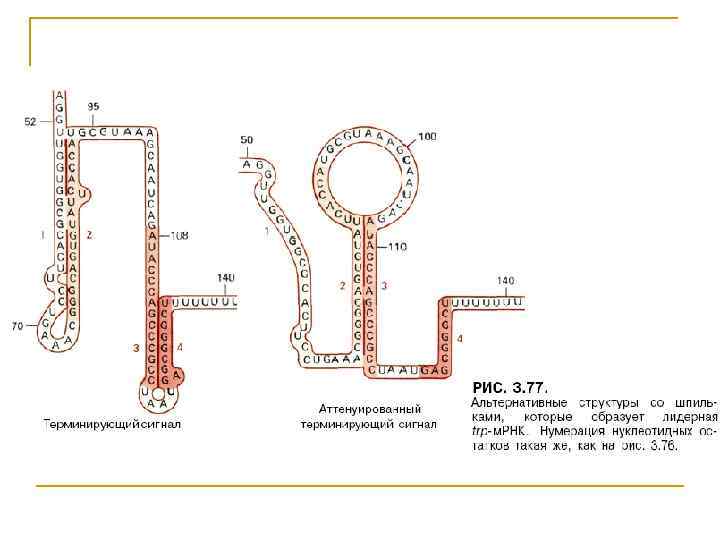
 На аттенуацию влияет трансляция, идущая параллельно с транскрипцией n n При недостатке триптофанил-т. РНК рибосомы останавливаются, когда доходят до тандемных триптофановых кодонов, расположенных в сегменте 1. Тогда последовательности сегментов 2 и 3 свободно спариваются и образуют пропускающую шпилечную структуру. При достаточном количестве триптофанилт. РНК рибосомы беспрепятственно движутся до терминаторного кодона лидерного пептида, ассоциируясь на данном участке с сегментом 2. Поэтому, предоставленные самим себе сегменты 3 и 4 спариваются, образуя терминирующую шпильку. Таким образом аттенуатор позволяет РНК-полимеразе опосредованно «почувствовать» концентрацию триптофанил-т. РНК через расположение рибосом.
На аттенуацию влияет трансляция, идущая параллельно с транскрипцией n n При недостатке триптофанил-т. РНК рибосомы останавливаются, когда доходят до тандемных триптофановых кодонов, расположенных в сегменте 1. Тогда последовательности сегментов 2 и 3 свободно спариваются и образуют пропускающую шпилечную структуру. При достаточном количестве триптофанилт. РНК рибосомы беспрепятственно движутся до терминаторного кодона лидерного пептида, ассоциируясь на данном участке с сегментом 2. Поэтому, предоставленные самим себе сегменты 3 и 4 спариваются, образуя терминирующую шпильку. Таким образом аттенуатор позволяет РНК-полимеразе опосредованно «почувствовать» концентрацию триптофанил-т. РНК через расположение рибосом.

 ИЗМЕНЧИВОСТЬ МИКРООРГАНИЗМОВ
ИЗМЕНЧИВОСТЬ МИКРООРГАНИЗМОВ
 Сосуществуют 2 геномных пространства. Одному отведена роль консервативная – сохранения и поддержания существующего генома, наработанного трудами предыдущих тысячелетий. Способствует кратковременному выживанию вида. Другому геномному пространству отведена экспериментирующая роль (эволюционная). Способствует долговременному выживанию вида. «Ген представляется мне крохотным архипелагом, в котором островки информации разбросаны среди обширного моря бессмыслицы…» Роберт Вайнберг. «The Scientist» , 1987
Сосуществуют 2 геномных пространства. Одному отведена роль консервативная – сохранения и поддержания существующего генома, наработанного трудами предыдущих тысячелетий. Способствует кратковременному выживанию вида. Другому геномному пространству отведена экспериментирующая роль (эволюционная). Способствует долговременному выживанию вида. «Ген представляется мне крохотным архипелагом, в котором островки информации разбросаны среди обширного моря бессмыслицы…» Роберт Вайнберг. «The Scientist» , 1987
 ГЕНЕТИЧЕСКИЕ МЕХАНИЗМЫ КЛЕТКИ ОБЕСПЕЧИВАЮТ: 1. Сохранение, воспроизведение и передачу наследственных признаков, что достигается с помощью механизмов: - репликации - экспрессии гена (транскрипция и трансляция) - репарации 2. Изменчивость наследственных признаков, что достигается с помощью : - мутаций -- спонтанные -- индуцированные - генетической рекомбинации -- общая гомологическая -- сайт-специфическая -- общая негомологическая
ГЕНЕТИЧЕСКИЕ МЕХАНИЗМЫ КЛЕТКИ ОБЕСПЕЧИВАЮТ: 1. Сохранение, воспроизведение и передачу наследственных признаков, что достигается с помощью механизмов: - репликации - экспрессии гена (транскрипция и трансляция) - репарации 2. Изменчивость наследственных признаков, что достигается с помощью : - мутаций -- спонтанные -- индуцированные - генетической рекомбинации -- общая гомологическая -- сайт-специфическая -- общая негомологическая
 Генетическая рекомбинация
Генетическая рекомбинация
 ОСНОВНЫЕ ПОНЯТИЯ n n n n n Генотип – информация, заключенная в ДНК клетки или организма, т. е. совокупность всех наследственных задатков клетки или организма Геном – совокупность всех ДНК клетки или организма Фенотип – совокупность наблюдаемых признаков клетки или организма: -особые белки - функции - морфологические (форма, окраска по Граму, наличие и расположение жгутиков, наличие капсулы и способность к спорообразованию) и - поведенческие признаки Генотипическая изменчивость – изменения в последовательности нуклеотидов Фенотипическая изменчивость – изменение наблюдаемых свойств клетки или организма Мутация – скачкообразное изменение наследственного признака (впервые сформулировано для растений Де Фризом)
ОСНОВНЫЕ ПОНЯТИЯ n n n n n Генотип – информация, заключенная в ДНК клетки или организма, т. е. совокупность всех наследственных задатков клетки или организма Геном – совокупность всех ДНК клетки или организма Фенотип – совокупность наблюдаемых признаков клетки или организма: -особые белки - функции - морфологические (форма, окраска по Граму, наличие и расположение жгутиков, наличие капсулы и способность к спорообразованию) и - поведенческие признаки Генотипическая изменчивость – изменения в последовательности нуклеотидов Фенотипическая изменчивость – изменение наблюдаемых свойств клетки или организма Мутация – скачкообразное изменение наследственного признака (впервые сформулировано для растений Де Фризом)
 СРАВНЕНИЕ ГЕНОТИПИЧЕСКОЙ И ФЕНОТИПИЧЕСКОЙ ИЗМЕНЧИВОСТЕЙ ГЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ФЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ 1. ПРИЧИНА: проявляется в результате мутаций и генетических рекомбинаций 1. ПРИЧИНА: проявляется в результате изменения внешних условий 2. ПРИРОДА: наследственная 2. ПРИРОДА: адаптационная 3. Обеспечивает долговременное выживание вида 3. Обеспечивает кратковременное выживание вида 4. Затрагивает немногие клетки 4. Затрагивает большинство особей в популяции 5. Устойчива 5. Не устойчива (исчезает со снятием фактора, ее вызвавшего)
СРАВНЕНИЕ ГЕНОТИПИЧЕСКОЙ И ФЕНОТИПИЧЕСКОЙ ИЗМЕНЧИВОСТЕЙ ГЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ ФЕНОТИПИЧЕСКАЯ ИЗМЕНЧИВОСТЬ 1. ПРИЧИНА: проявляется в результате мутаций и генетических рекомбинаций 1. ПРИЧИНА: проявляется в результате изменения внешних условий 2. ПРИРОДА: наследственная 2. ПРИРОДА: адаптационная 3. Обеспечивает долговременное выживание вида 3. Обеспечивает кратковременное выживание вида 4. Затрагивает немногие клетки 4. Затрагивает большинство особей в популяции 5. Устойчива 5. Не устойчива (исчезает со снятием фактора, ее вызвавшего)
 ВАЖНО!!! Фенотипическое проявление одного и того же генотипа может быть различным в зависимости от условий окружающей среды
ВАЖНО!!! Фенотипическое проявление одного и того же генотипа может быть различным в зависимости от условий окружающей среды
 МУТАЦИИ n n n n n Сущность: нарушение нуклеотидной последовательности в ДНК клетки Спонтанные Причина: ошибки при репликации из-за возможности существования оснований в различных таутомерных формах Частота мутаций: 10 -6 – 10 -8 Индуцированные Причина: ошибки при репликации под воздействием мутагенов: - химические - физические - биологические Частота мутаций: 10 -4 – 10 -6
МУТАЦИИ n n n n n Сущность: нарушение нуклеотидной последовательности в ДНК клетки Спонтанные Причина: ошибки при репликации из-за возможности существования оснований в различных таутомерных формах Частота мутаций: 10 -6 – 10 -8 Индуцированные Причина: ошибки при репликации под воздействием мутагенов: - химические - физические - биологические Частота мутаций: 10 -4 – 10 -6
 ОБЩАЯ ГОМОЛОГИЧНАЯ ГЕНЕТИЧЕСКАЯ РЕКОМБИНАЦИЯ n n n Сущность: две молекулы ДНК рекомбинируются путем реципрокного (взаимодополняющего) обмена протяженными гомологичными участками. Гомологичная рекомбинация находится под контролем гена Rec A.
ОБЩАЯ ГОМОЛОГИЧНАЯ ГЕНЕТИЧЕСКАЯ РЕКОМБИНАЦИЯ n n n Сущность: две молекулы ДНК рекомбинируются путем реципрокного (взаимодополняющего) обмена протяженными гомологичными участками. Гомологичная рекомбинация находится под контролем гена Rec A.
 Схема, иллюстрирующая начальный одноцепочечный между двумя обмен гомологичными двойными спиралями ДНК в процессе общей рекомбинации. Разрыв в одной из цепей высвобождает эту цепь и она внедряется во ДНК вторую спираль, образуя здесь короткий спаренный участок. Спариваться таким путем и тем самым инициировать общую рекомбинацию могут только такие две молекулы ДНК, у которых нуклеотидные последовательности комплементарны.
Схема, иллюстрирующая начальный одноцепочечный между двумя обмен гомологичными двойными спиралями ДНК в процессе общей рекомбинации. Разрыв в одной из цепей высвобождает эту цепь и она внедряется во ДНК вторую спираль, образуя здесь короткий спаренный участок. Спариваться таким путем и тем самым инициировать общую рекомбинацию могут только такие две молекулы ДНК, у которых нуклеотидные последовательности комплементарны.
 Следующим этапом является разрыв в одной из цепей второй ДНК обмен с перекрещиванием цепей и. Завершающий этап - разрыв точки обмена и возникновение двух новых ДНК. Много возможных путей ведет от структуры, представленной на слайде с одноцепочечным обменом, к структуре с перекрещенными цепями. Здесь показан один из таких путей, который дает два альтернативных результата: - обмен состоялся только между двумя цепями; - в обмене участвовали все четыре цепи
Следующим этапом является разрыв в одной из цепей второй ДНК обмен с перекрещиванием цепей и. Завершающий этап - разрыв точки обмена и возникновение двух новых ДНК. Много возможных путей ведет от структуры, представленной на слайде с одноцепочечным обменом, к структуре с перекрещенными цепями. Здесь показан один из таких путей, который дает два альтернативных результата: - обмен состоялся только между двумя цепями; - в обмене участвовали все четыре цепи
 Обмен гомологичными участками ДНК без кроссинговера
Обмен гомологичными участками ДНК без кроссинговера
 Обмен гомологичными участками ДНК с кроссинговером
Обмен гомологичными участками ДНК с кроссинговером
 При гибридизации ДНК in vitro двойные спирали ДНК образуются заново из ранее разделившихся цепей. Восстановление спиралей зависит от случайного cоударения двух комплементарных целей. Большинство таких соударений безрезультатно (как это видно из левой части рисунка), но некоторые из них приводят к спариванию на коротком участке комплементарных оснований (т. е. к нуклеации спирали). За этим следует быстрое «застегивание молнии» и двойная спираль готова. Посредством такого процесса - методом проб и ошибок каждая цепь ДНК может найти себе комплементарного партнера среди миллионов «неподходящих» цепей.
При гибридизации ДНК in vitro двойные спирали ДНК образуются заново из ранее разделившихся цепей. Восстановление спиралей зависит от случайного cоударения двух комплементарных целей. Большинство таких соударений безрезультатно (как это видно из левой части рисунка), но некоторые из них приводят к спариванию на коротком участке комплементарных оснований (т. е. к нуклеации спирали). За этим следует быстрое «застегивание молнии» и двойная спираль готова. Посредством такого процесса - методом проб и ошибок каждая цепь ДНК может найти себе комплементарного партнера среди миллионов «неподходящих» цепей.
 в хромосому клеткихозяина по строго определенному сайту") Сайт-специфическая рекомбинация Сущность: чужеродная ДНК внедряется (интегрируется) в хромосому клеткихозяина по строго определенному сайту (между генами gal и bio) Пример: внедрение ДНК бактериофага λ в хромосому E. coli
Сайт-специфическая рекомбинация Сущность: чужеродная ДНК внедряется (интегрируется) в хромосому клеткихозяина по строго определенному сайту (между генами gal и bio) Пример: внедрение ДНК бактериофага λ в хромосому E. coli
 в хромосому клетки-хозяина в") ОБЩАЯ НЕГОМОЛОГИЧНАЯ ГЕНЕТИЧЕСКАЯ РЕКОМБИНАЦИЯ n Сущность: чужеродная ДНК внедряется (интегрируется) в хромосому клетки-хозяина в любом месте n Примеры: гигантский транспозон (бактериофаг мю), мобильные генетические элементы, инсерционные последовательности
ОБЩАЯ НЕГОМОЛОГИЧНАЯ ГЕНЕТИЧЕСКАЯ РЕКОМБИНАЦИЯ n Сущность: чужеродная ДНК внедряется (интегрируется) в хромосому клетки-хозяина в любом месте n Примеры: гигантский транспозон (бактериофаг мю), мобильные генетические элементы, инсерционные последовательности
 СПОСОБЫ ДОСТАВКИ ЧУЖЕРОДНОЙ ДНК В КЛЕТКУ n n n Конъюгация – при непосредственном контакте двух клеток Например: наиболее характерна для энтеробактерий Трансдукция – с помощью переносчика Например: обнаружена у Bacillus, Pseudomonas, Staphylococcus и др. Трансформация – самостоятельное попадание свободной компетентную клетку Например: E. coli ДНК в
СПОСОБЫ ДОСТАВКИ ЧУЖЕРОДНОЙ ДНК В КЛЕТКУ n n n Конъюгация – при непосредственном контакте двух клеток Например: наиболее характерна для энтеробактерий Трансдукция – с помощью переносчика Например: обнаружена у Bacillus, Pseudomonas, Staphylococcus и др. Трансформация – самостоятельное попадание свободной компетентную клетку Например: E. coli ДНК в
 КЛЕТОЧНЫЙ ЦИКЛ БАКТЕРИОФАГА Т 2
КЛЕТОЧНЫЙ ЦИКЛ БАКТЕРИОФАГА Т 2
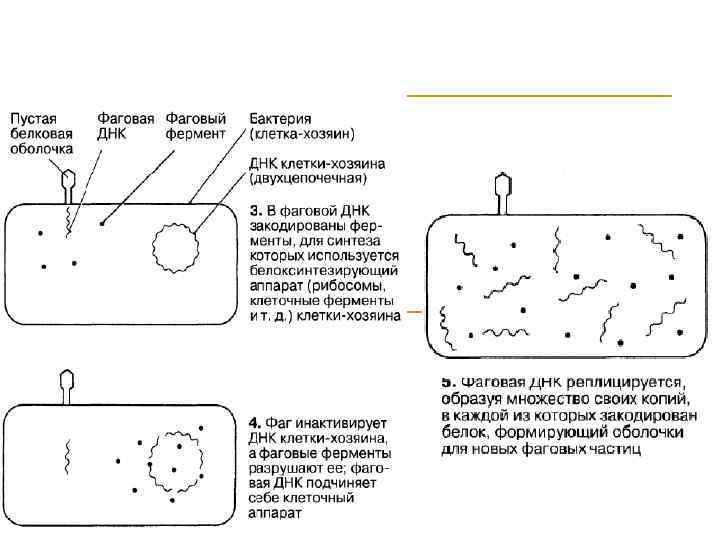
") КЛЕТОЧНЫЙ ЦИКЛ БАКТЕРИОФАГА Т 2 (ОКОНЧАНИЕ)
КЛЕТОЧНЫЙ ЦИКЛ БАКТЕРИОФАГА Т 2 (ОКОНЧАНИЕ)