

4 11 2016 ДДД НУКЛЕОТИД АЛМАСУЫ.ppt
- Количество слайдов: 142
 Нуклеотидтердің алмасуы. .
Нуклеотидтердің алмасуы. .
 Жоспар: • • • Нуклеотидтер. Құрылысы. Қызметі Нуклеотидтердің жіктелуі Асқорту жолдары мен тіңнің нуклеазалары. Пиримидинді нуклеотид ыдырауы Пуринді нуклеотид ыдырауы Пиримидинді нуклеотид биосинтезі Пуринді нуклеотид биосинтезі Дезоксирибонуклеотидтердің биосинтезі Нуклеотидтердің алмасуын реттеу Нуклеидтер алмасуының бұзылыстары.
Жоспар: • • • Нуклеотидтер. Құрылысы. Қызметі Нуклеотидтердің жіктелуі Асқорту жолдары мен тіңнің нуклеазалары. Пиримидинді нуклеотид ыдырауы Пуринді нуклеотид ыдырауы Пиримидинді нуклеотид биосинтезі Пуринді нуклеотид биосинтезі Дезоксирибонуклеотидтердің биосинтезі Нуклеотидтердің алмасуын реттеу Нуклеидтер алмасуының бұзылыстары.
 Нуклеопротеидтер- күрделі белоктар НҚ + өзг. белок Нуклеопротеидтерге хромосомалар және рибосомалары жатады. Нуклеопротеидтердің негізгі қызметі -генетикалық ақпарат сақтау және іске асыру. Белок синтезін қамтамасыз ету .
Нуклеопротеидтер- күрделі белоктар НҚ + өзг. белок Нуклеопротеидтерге хромосомалар және рибосомалары жатады. Нуклеопротеидтердің негізгі қызметі -генетикалық ақпарат сақтау және іске асыру. Белок синтезін қамтамасыз ету .
 Нуклеопротеидтердің құрылымдық компонеттері - нуклеин қышқылдары Полинуклеотидті тізбектің құрылымдық бірлігімононуклеотидтер
Нуклеопротеидтердің құрылымдық компонеттері - нуклеин қышқылдары Полинуклеотидті тізбектің құрылымдық бірлігімононуклеотидтер

 , фосфор қышқылы. H-C-OH NH N") Нуклеотидтер- пуриндік немесе пиримидиндік негіздер, пентоза (рибоза, дезоксирибоза) , фосфор қышқылы. H-C-OH NH N пурин N пиримидин H-C-H H-C-OH N H-C-OH H-C-ОН CH 2 OH D-рибоза CH 2 OH D-дезоксирибоза
Нуклеотидтер- пуриндік немесе пиримидиндік негіздер, пентоза (рибоза, дезоксирибоза) , фосфор қышқылы. H-C-OH NH N пурин N пиримидин H-C-H H-C-OH N H-C-OH H-C-ОН CH 2 OH D-рибоза CH 2 OH D-дезоксирибоза

 Азотты негіздер ПУРИНДІ ПИРИМИДИНДІ N NH N N
Азотты негіздер ПУРИНДІ ПИРИМИДИНДІ N NH N N
 Пиримидин Цитозин Урацил Тимин
Пиримидин Цитозин Урацил Тимин
 Цитозин Урацил Тимин Аденин Гуанин
Цитозин Урацил Тимин Аденин Гуанин
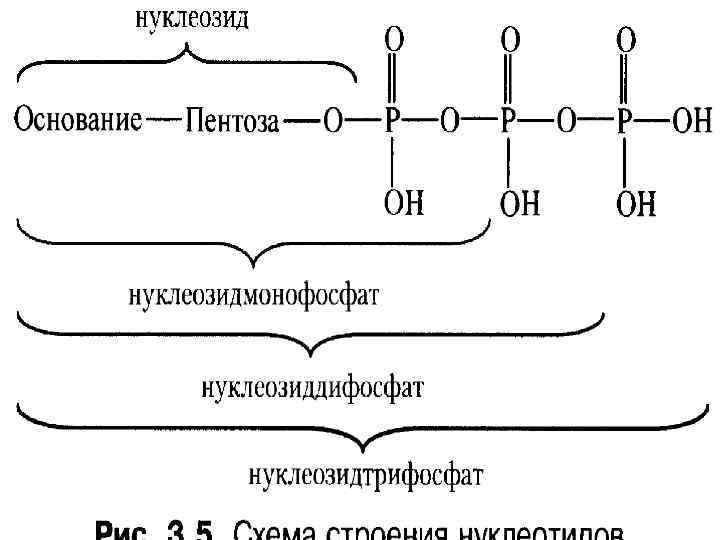
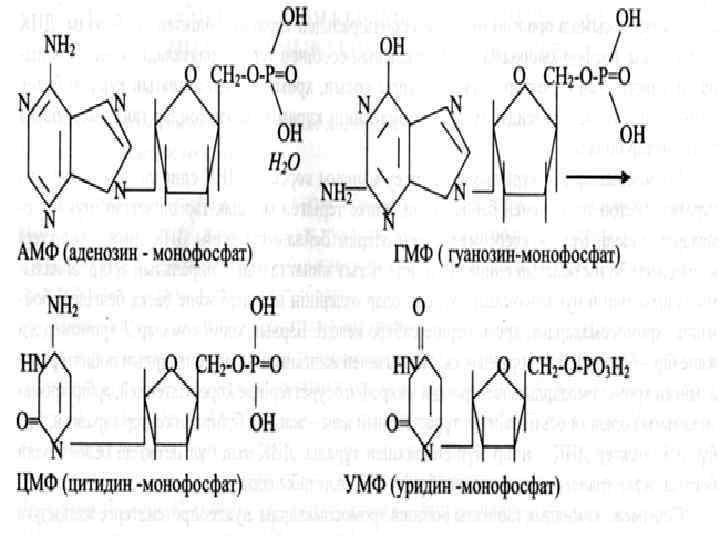
 Пуриниді негіздер: аденин, гуанин Пиринді негіздер: тимин, цитозин, урацил Нуклеозидтер: аденозин, гуанозин, тимидин, цитидин, уридин Нуклеотидтер: АМФ, ГМФ, ТМФ, ЦМФ, УМФ
Пуриниді негіздер: аденин, гуанин Пиринді негіздер: тимин, цитозин, урацил Нуклеозидтер: аденозин, гуанозин, тимидин, цитидин, уридин Нуклеотидтер: АМФ, ГМФ, ТМФ, ЦМФ, УМФ


 Дезокси-АТФ
Дезокси-АТФ
 Нуклеотидтердін биологиялық мәні: • Нуклеин қышқылының құрылымдық компоненттері болып табылады; • Клеткада энергия жинақтау және трансформациялау механизмдеріне қатысып, АТФ, ГТФ, ЦТФ және т. б макроэргтер түзеді; • Коферменттердің ( НАД, НАДФ, ФАД, ) құрамына кіреді. • 3´, 5´- АМФ және 3´, 5´- ГМФ циклды нуклеотидтерді түзіп , жасушаішілік эффекторлы жүйелерге гормональді және т. б. белгілерді жеткізуге қатысады
Нуклеотидтердін биологиялық мәні: • Нуклеин қышқылының құрылымдық компоненттері болып табылады; • Клеткада энергия жинақтау және трансформациялау механизмдеріне қатысып, АТФ, ГТФ, ЦТФ және т. б макроэргтер түзеді; • Коферменттердің ( НАД, НАДФ, ФАД, ) құрамына кіреді. • 3´, 5´- АМФ және 3´, 5´- ГМФ циклды нуклеотидтерді түзіп , жасушаішілік эффекторлы жүйелерге гормональді және т. б. белгілерді жеткізуге қатысады
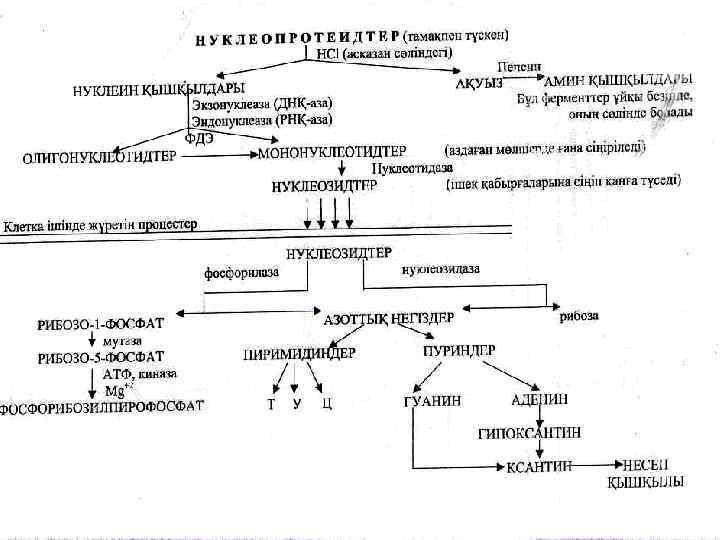
 Нуклеопротеидтердің қорытылуы Жануартектес тағам нуклеопротеидтерге өте бай. Асқазанда тұз қышқылының әсерінен нуклеопротеидтер нуклеин қышқылдарына және белокқа дейін ыдырайды. Нуклеопротеидтердің белокты бөлігі асқазан-ішек жолдарында аминқышқылдарына дейін қорытылады.
Нуклеопротеидтердің қорытылуы Жануартектес тағам нуклеопротеидтерге өте бай. Асқазанда тұз қышқылының әсерінен нуклеопротеидтер нуклеин қышқылдарына және белокқа дейін ыдырайды. Нуклеопротеидтердің белокты бөлігі асқазан-ішек жолдарында аминқышқылдарына дейін қорытылады.
 Нуклеин қышқылдары панкреатикалық сөлінің нуклеазалары әсерінен ыдырайды / гидролиз /. Нуклеазалар 5‘ 3‘фосфоэфирлік байланыстың гидролизін катализдейді.
Нуклеин қышқылдары панкреатикалық сөлінің нуклеазалары әсерінен ыдырайды / гидролиз /. Нуклеазалар 5‘ 3‘фосфоэфирлік байланыстың гидролизін катализдейді.
 Основные функции нуклеотидов
Основные функции нуклеотидов
 Нуклеазалар ДНҚ-аза ДНҚ-ның полинуклеотидтік тізбегін гидролиздейді РНҚ-аза РНҚ-ның поли нуклеотидтік тізбегін гидролиздейді
Нуклеазалар ДНҚ-аза ДНҚ-ның полинуклеотидтік тізбегін гидролиздейді РНҚ-аза РНҚ-ның поли нуклеотидтік тізбегін гидролиздейді
 • Нуклеин қыш-ң гидролиздену нәтижесінде мононуклеотидтер • (АМФ, ГМФ, ЦМФ, УМФ, d-ТМФ, d-АМФ, d-ГМФ, d-ЦМФ) пайда болады. Арығарай • мононуклеотидтер ішектің 5`- нуклеотидазаларының әсерімен нуклеозидтер және фосфор қышқылға дейін гидролизденеді:
• Нуклеин қыш-ң гидролиздену нәтижесінде мононуклеотидтер • (АМФ, ГМФ, ЦМФ, УМФ, d-ТМФ, d-АМФ, d-ГМФ, d-ЦМФ) пайда болады. Арығарай • мононуклеотидтер ішектің 5`- нуклеотидазаларының әсерімен нуклеозидтер және фосфор қышқылға дейін гидролизденеді:
 • Нуклеин қыш-ң гидролиздену нәтижесінде мононуклеотидтер • (АМФ, ГМФ, ЦМФ, УМФ, d-ТМФ, d-АМФ, d-ГМФ, d-ЦМФ) пайда болады. • •
• Нуклеин қыш-ң гидролиздену нәтижесінде мононуклеотидтер • (АМФ, ГМФ, ЦМФ, УМФ, d-ТМФ, d-АМФ, d-ГМФ, d-ЦМФ) пайда болады. • •
 Арығарай мононуклеотидтер ішектің 5`нуклеотидазаларының әсерімен нуклеозидтер және фосфор қышқылға дейін гидролизденеді:
Арығарай мононуклеотидтер ішектің 5`нуклеотидазаларының әсерімен нуклеозидтер және фосфор қышқылға дейін гидролизденеді:
 Нуклеозидтер: аденозин, гуанозин, тимидин, цитидин, уридин
Нуклеозидтер: аденозин, гуанозин, тимидин, цитидин, уридин
 Переваривание • • • • Нуклеазы панкреатического сока катализируют гидролиз 5`3`-фосфоэфирные связи. Различают два типа нуклеаз: ДНК-азы и РНК-аза. В результате гидролиза полинуклеотидных цепей нуклеиновых кислот образуются мононуклеотиды (АМФ, ГМФ, ЦМФ, УМФ, d-ТМФ, d-АМФ, d-ГМФ, d-ЦМФ). Мононуклеотиды под воздействием 5`-нуклетидаз кишечника разрушаются на нуклеозиды и фосфорную кислоту: АМФ Н 2 О 5`-нуклетидаза ЦМФ Н 2 О 5`-нуклетидаза аденозин + фосфорная кислота цитидин + фосфорная кислота Нуклеозиды в свою очередь разрушаются нуклеозидазами кишечного сока до азотистых оснований и пентозы (рибоза или дезоксирибоза): d-тимидин Н 2 О нуклеозидаза тимин + дезоксирибоза
Переваривание • • • • Нуклеазы панкреатического сока катализируют гидролиз 5`3`-фосфоэфирные связи. Различают два типа нуклеаз: ДНК-азы и РНК-аза. В результате гидролиза полинуклеотидных цепей нуклеиновых кислот образуются мононуклеотиды (АМФ, ГМФ, ЦМФ, УМФ, d-ТМФ, d-АМФ, d-ГМФ, d-ЦМФ). Мононуклеотиды под воздействием 5`-нуклетидаз кишечника разрушаются на нуклеозиды и фосфорную кислоту: АМФ Н 2 О 5`-нуклетидаза ЦМФ Н 2 О 5`-нуклетидаза аденозин + фосфорная кислота цитидин + фосфорная кислота Нуклеозиды в свою очередь разрушаются нуклеозидазами кишечного сока до азотистых оснований и пентозы (рибоза или дезоксирибоза): d-тимидин Н 2 О нуклеозидаза тимин + дезоксирибоза
 ҚОРЫТЫЛУ Нуклеазалардың / ДНК аза, РНК-аза / әсерімен полинуклеотидты тіізбектер мононуклеотидтерге дейін гидролизденеді (АМФ, ГМФ, ЦМФ, УМФ, d ТМФ, d-АМФ, d-ГМФ, d-ЦМФ)
ҚОРЫТЫЛУ Нуклеазалардың / ДНК аза, РНК-аза / әсерімен полинуклеотидты тіізбектер мононуклеотидтерге дейін гидролизденеді (АМФ, ГМФ, ЦМФ, УМФ, d ТМФ, d-АМФ, d-ГМФ, d-ЦМФ)
 ҚОРЫТЫЛУ Арығарай олар ішектің 5`нуклеотидазалары ның әсерімен нуклеозидтер, фосфор қышқылы на ыдырайды
ҚОРЫТЫЛУ Арығарай олар ішектің 5`нуклеотидазалары ның әсерімен нуклеозидтер, фосфор қышқылы на ыдырайды
 АМФ • Н 2 О аденозин + Н 3 РО 4 5`-нуклеотидаза • • ЦМФ Н 2 О 5`-нуклеотидаза цитидин + Н 3 РО 4
АМФ • Н 2 О аденозин + Н 3 РО 4 5`-нуклеотидаза • • ЦМФ Н 2 О 5`-нуклеотидаза цитидин + Н 3 РО 4
 гидролизденеді.") Нуклеозидтер ішектің нуклеозидазаларының әсерімен азотты негіздер , пентозаларға дейін (рибоза , дезоксирибоза) гидролизденеді. Тимидин Н 2 О нуклеотидаза тимин +
Нуклеозидтер ішектің нуклеозидазаларының әсерімен азотты негіздер , пентозаларға дейін (рибоза , дезоксирибоза) гидролизденеді. Тимидин Н 2 О нуклеотидаза тимин +
 Пуриниді негіздер: аденин, гуанин Пиринді негіздер: тимин, цитозин, урацил Пентозалар рибоза дезоксирибоза
Пуриниді негіздер: аденин, гуанин Пиринді негіздер: тимин, цитозин, урацил Пентозалар рибоза дезоксирибоза
 H-C-OH H-C-OH H-C CH 2 OH D-рибоза CH 2 OH D-дезоксирибоза") D-рибоза, D-дезоксирибоза) H-C-OH H-C-OH H-C CH 2 OH D-рибоза CH 2 OH D-дезоксирибоза
D-рибоза, D-дезоксирибоза) H-C-OH H-C-OH H-C CH 2 OH D-рибоза CH 2 OH D-дезоксирибоза
 Ішекте нуклеозидтер, азотты негіздер, пентозалар , фосфор қышқылы, АМҚ сіңіріледі. Ішектен қан ағынымен нуклеопротеид тердің қорытылу өнімдері бауырға және басқа мүшелер мен тіндерге жеткізіледі.
Ішекте нуклеозидтер, азотты негіздер, пентозалар , фосфор қышқылы, АМҚ сіңіріледі. Ішектен қан ағынымен нуклеопротеид тердің қорытылу өнімдері бауырға және басқа мүшелер мен тіндерге жеткізіледі.
 Пурінді және пириминді нуклеотидтердін аралық алмасуы
Пурінді және пириминді нуклеотидтердін аралық алмасуы
 Пурин нуклеотидтердің ыдырауы
Пурин нуклеотидтердің ыдырауы

 Зәр қышқылы зәрғе
Зәр қышқылы зәрғе


 .
.
 Пуринді нуклеотидтердің ыдырауның соңғы өнімі-зәр қышқылы. Ағзада тәулігіне 0, 5 -1, 0 гр. зәр қышқылы түзіліп , зәрмен шығарылады. Қанда 0, 16 -0, 57 ммоль/л
Пуринді нуклеотидтердің ыдырауның соңғы өнімі-зәр қышқылы. Ағзада тәулігіне 0, 5 -1, 0 гр. зәр қышқылы түзіліп , зәрмен шығарылады. Қанда 0, 16 -0, 57 ммоль/л
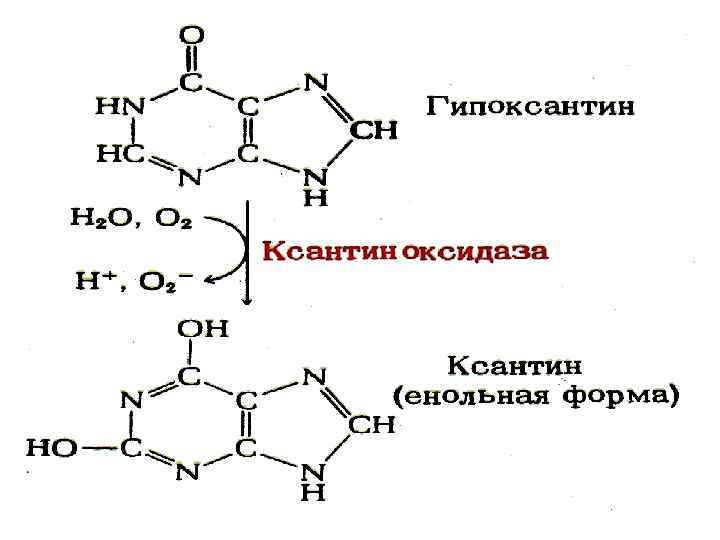
 Ксантинурия. Себебі – ксантиноксидаза ның туа біткен жетіспеушілігі
Ксантинурия. Себебі – ксантиноксидаза ның туа біткен жетіспеушілігі
 Зәр қышқылы
Зәр қышқылы
 Ксантинурияда –бүйрек каналшаларында ксан тиннің реабсорбциясы ның бұзылуымен ілеседі Нәтижесінде зәр өтетің жолдарында ксантинді тастар түзіледі.
Ксантинурияда –бүйрек каналшаларында ксан тиннің реабсорбциясы ның бұзылуымен ілеседі Нәтижесінде зәр өтетің жолдарында ксантинді тастар түзіледі.
 Пуринді нуклеотидтер дін биосинтезі
Пуринді нуклеотидтер дін биосинтезі
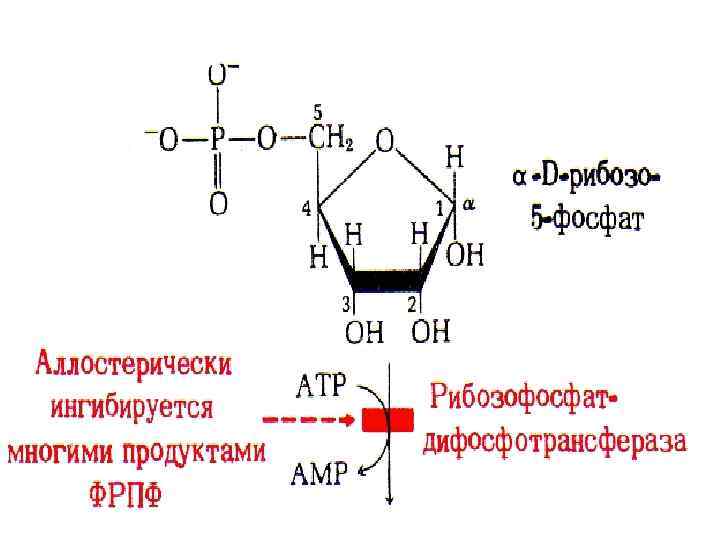
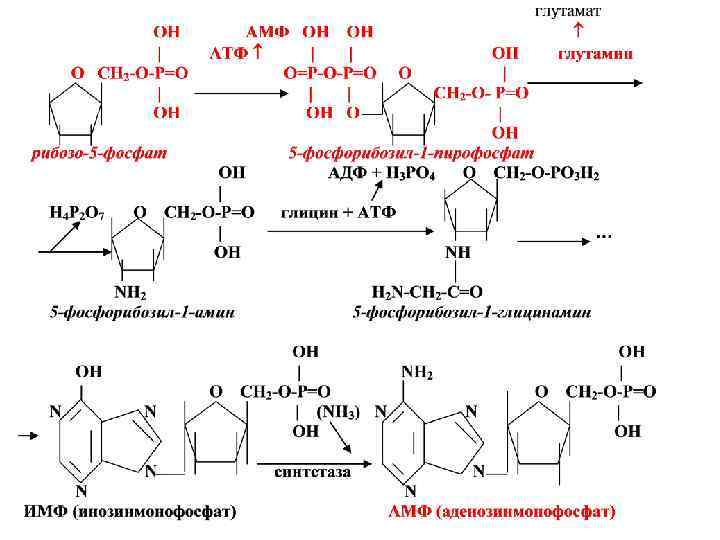
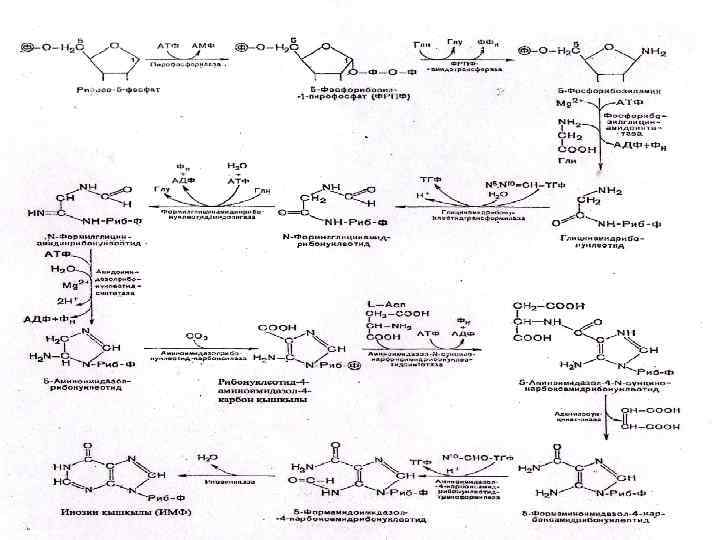
 Биосинтез пуриновых нуклеотидов • Для биосинтеза пуриновых нуклеотидов необходимо наличие глутамина, глицина, аспаратата, углекислого газа, формил. ТГФК и метенил-ТГФК - соединений, из которых формируется пуриновое ядро. • Формирование пуринового ядра происходит на рибозо-5 -фосфате, который активируется за счет АТФ превращаясь в 5 -фосфорибозил-1 -пирофосфат.
Биосинтез пуриновых нуклеотидов • Для биосинтеза пуриновых нуклеотидов необходимо наличие глутамина, глицина, аспаратата, углекислого газа, формил. ТГФК и метенил-ТГФК - соединений, из которых формируется пуриновое ядро. • Формирование пуринового ядра происходит на рибозо-5 -фосфате, который активируется за счет АТФ превращаясь в 5 -фосфорибозил-1 -пирофосфат.
 Биосинтез пуриновых нуклеотидов • Для биосинтеза пуриновых нуклеотидов необходимо наличие глутамина, глицина, аспаратата, углекислого газа, формил. ТГФК и метенил-ТГФК - соединений, из которых формируется пуриновое ядро. • Формирование пуринового ядра происходит на рибозо-5 -фосфате, который активируется за счет АТФ превращаясь в 5 -фосфорибозил-1 -пирофосфат.
Биосинтез пуриновых нуклеотидов • Для биосинтеза пуриновых нуклеотидов необходимо наличие глутамина, глицина, аспаратата, углекислого газа, формил. ТГФК и метенил-ТГФК - соединений, из которых формируется пуриновое ядро. • Формирование пуринового ядра происходит на рибозо-5 -фосфате, который активируется за счет АТФ превращаясь в 5 -фосфорибозил-1 -пирофосфат.
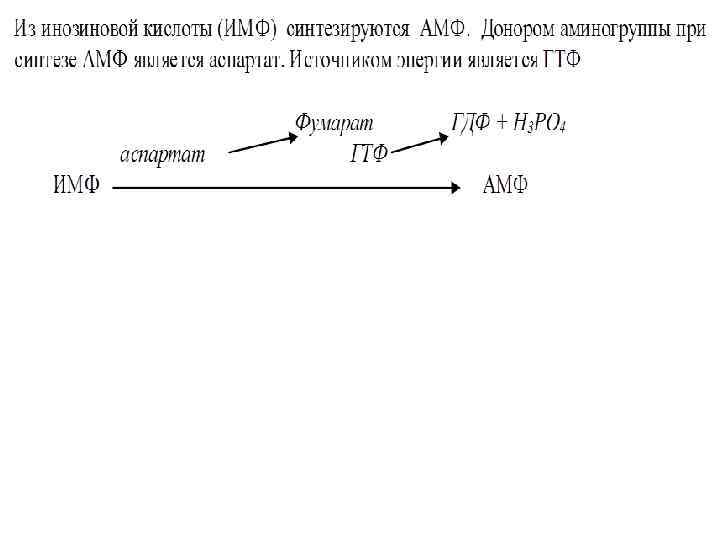
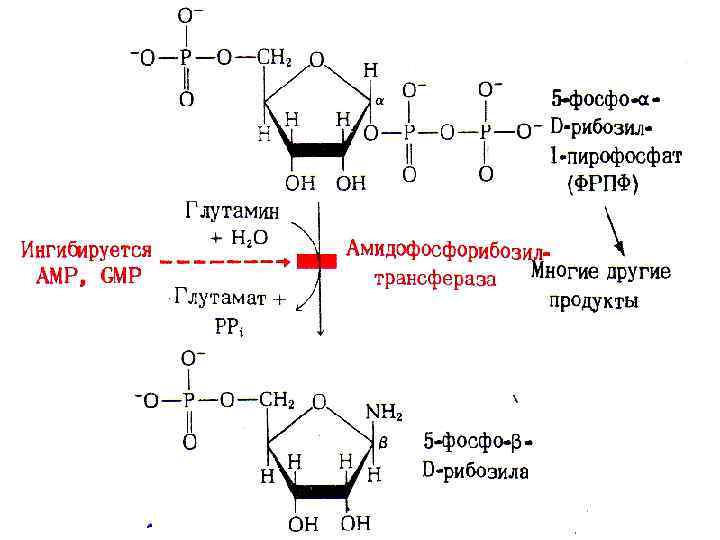
 • Затем под воздействием фермента амидинофосфо- рибозил трансферазы с использованием аминогруппы глутамина образуется 5 -фосфорибозиламин. • Далее в последовательных реакциях синтеза к 5 -фосфорибозиламину присоединяется остаток глицина, метенильный радикал т. д. • На заключительной стадии образуется инозинмонофосфат (ИМФ):
• Затем под воздействием фермента амидинофосфо- рибозил трансферазы с использованием аминогруппы глутамина образуется 5 -фосфорибозиламин. • Далее в последовательных реакциях синтеза к 5 -фосфорибозиламину присоединяется остаток глицина, метенильный радикал т. д. • На заключительной стадии образуется инозинмонофосфат (ИМФ):
 • Затем под воздействием фермента амидинофосфо- рибозил трансферазы с использованием аминогруппы глутамина образуется 5 -фосфорибозиламин. • Далее в последовательных реакциях синтеза к 5 -фосфорибозиламину присоединяется остаток глицина, метенильный радикал т. д. • На заключительной стадии образуется инозинмонофосфат (ИМФ):
• Затем под воздействием фермента амидинофосфо- рибозил трансферазы с использованием аминогруппы глутамина образуется 5 -фосфорибозиламин. • Далее в последовательных реакциях синтеза к 5 -фосфорибозиламину присоединяется остаток глицина, метенильный радикал т. д. • На заключительной стадии образуется инозинмонофосфат (ИМФ):
 Синтез ГМФ
Синтез ГМФ
 Синтез ГМФ
Синтез ГМФ
 Пуринді негіздердің пуринді сакинаның биосинтезі глицин, аспартат, глутамин қышқылының, тетрагидрофоль қышқылының , СО 2 қатысуымен жүреді. Пуринді сакинаның қалыптасуы рибозо-5 -фосфатан басталады. Ал ол 5 фосфорибозил-1 -пирофосфатка айналады.
Пуринді негіздердің пуринді сакинаның биосинтезі глицин, аспартат, глутамин қышқылының, тетрагидрофоль қышқылының , СО 2 қатысуымен жүреді. Пуринді сакинаның қалыптасуы рибозо-5 -фосфатан басталады. Ал ол 5 фосфорибозил-1 -пирофосфатка айналады.
 Пуринді негіздердің пуринді сакинаның биосинтезі глицин, аспартат, глутамин қышқылының, тетрагидрофоль қышқылының , СО 2 қатысуымен жүреді. Пуринді сакинаның қалыптасуы рибозо-5 -фосфатан басталады. Ал ол 5 фосфорибозил-1 -пирофосфатка айналады.
Пуринді негіздердің пуринді сакинаның биосинтезі глицин, аспартат, глутамин қышқылының, тетрагидрофоль қышқылының , СО 2 қатысуымен жүреді. Пуринді сакинаның қалыптасуы рибозо-5 -фосфатан басталады. Ал ол 5 фосфорибозил-1 -пирофосфатка айналады.
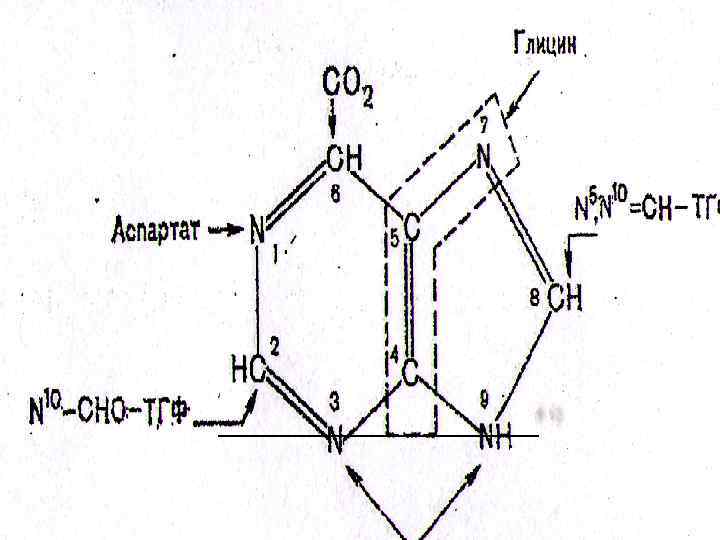
 СО 2 Глицин Аспартат Формил ТГФҚ Глутамин Пуринді сақинаның атомдарының қайнар көздері
СО 2 Глицин Аспартат Формил ТГФҚ Глутамин Пуринді сақинаның атомдарының қайнар көздері

 ГТФ асп ИМФ ----------- АМФ АМФ-синтетаза АМФ + АТФ ---------- АДФ аденилаткиназа бос энергия МХ-да: АДФ + Н 3 РО 4 --------- АТФ Тын. алу тіз. АТФ - синтетаза
ГТФ асп ИМФ ----------- АМФ АМФ-синтетаза АМФ + АТФ ---------- АДФ аденилаткиназа бос энергия МХ-да: АДФ + Н 3 РО 4 --------- АТФ Тын. алу тіз. АТФ - синтетаза
 НАДН 2 ИМФ ---------- > ксантинмонофосфат ----ГЛН АТФ ---------- ГМФ – синтетаза ГМФ + АТФ ------ ГДФ + АДФ киназа ГДФ + АТФ ------- ГТФ + АДФ киназа
НАДН 2 ИМФ ---------- > ксантинмонофосфат ----ГЛН АТФ ---------- ГМФ – синтетаза ГМФ + АТФ ------ ГДФ + АДФ киназа ГДФ + АТФ ------- ГТФ + АДФ киназа
 Нуклеотидтер алмасуын реттеу • Пуринді негіздердің биосинтезі. • Ең бірінші реттеуші фермент амидофосфорибозилтрансфе раза-ең бастапқы кезеніңде, яғни • 5 -фосфорибозил-1 пирофосфаттан 5 фосфорибозиламиннің синтезін реттейді.
Нуклеотидтер алмасуын реттеу • Пуринді негіздердің биосинтезі. • Ең бірінші реттеуші фермент амидофосфорибозилтрансфе раза-ең бастапқы кезеніңде, яғни • 5 -фосфорибозил-1 пирофосфаттан 5 фосфорибозиламиннің синтезін реттейді.
 Рибозанның дезоксирибозаға ауысуы Тотықсызданған Тотыққан
Рибозанның дезоксирибозаға ауысуы Тотықсызданған Тотыққан
 Пуринді негіздердің синтезделуінің қосымша жолдары: • Аденозин+АТФ АМФ + АДФ; • АМФ+АТФ АТФ+АДФ; • А+ФРПФ АМФ+H 4 P 2 O 7
Пуринді негіздердің синтезделуінің қосымша жолдары: • Аденозин+АТФ АМФ + АДФ; • АМФ+АТФ АТФ+АДФ; • А+ФРПФ АМФ+H 4 P 2 O 7
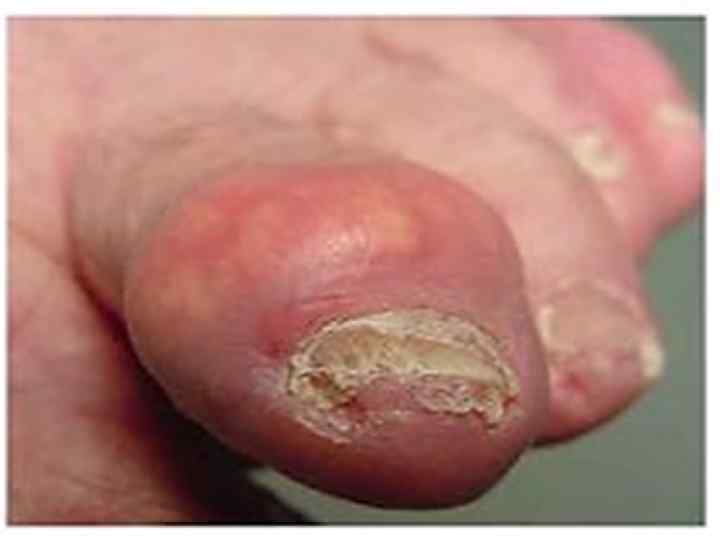
 1. Ағзадан зәр") Подаграның себептері 1. Пуриндер синтезінің жоғарылауы ( фосфорибозилпирофосфат синтетазаның белсенділігінің күшеюі) 1. Ағзадан зәр қышқылының шығарылуының бұзылуы ( зәр қышқылдың каналша реабсорбциясының жоғарылуы)
Подаграның себептері 1. Пуриндер синтезінің жоғарылауы ( фосфорибозилпирофосфат синтетазаның белсенділігінің күшеюі) 1. Ағзадан зәр қышқылының шығарылуының бұзылуы ( зәр қышқылдың каналша реабсорбциясының жоғарылуы)
 • Подаграда зәр қышқылынан, аздап аммоний немесе натрий тұздарынан тұратын тастар болады. Бұл тастар ураттар деп аталады.
• Подаграда зәр қышқылынан, аздап аммоний немесе натрий тұздарынан тұратын тастар болады. Бұл тастар ураттар деп аталады.
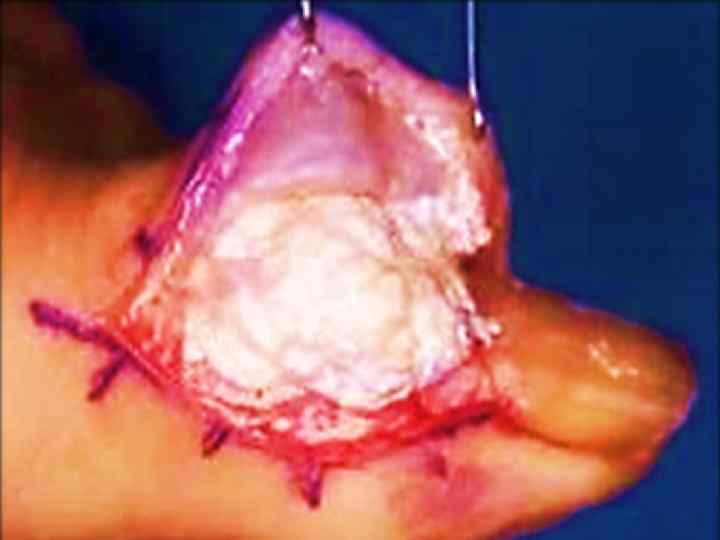


 Тіндерде зәр қышқылдың кристалдары: буынды шеміршектерде, синовиальды қабықшада, тері астындағы клетчаткада жинақталады.
Тіндерде зәр қышқылдың кристалдары: буынды шеміршектерде, синовиальды қабықшада, тері астындағы клетчаткада жинақталады.
 2. Пробенецид – зәр қышқылының шығуын") Емдеу: 1. Зәр қышқылының синтезін төмендету ( аллопуринол) 2. Пробенецид – зәр қышқылының шығуын күшейтеді
Емдеу: 1. Зәр қышқылының синтезін төмендету ( аллопуринол) 2. Пробенецид – зәр қышқылының шығуын күшейтеді


 Аллопуринол ксантиноксидазаның сыбайлас ингибиторы
Аллопуринол ксантиноксидазаның сыбайлас ингибиторы
 из гипоксантина и") Нарушения обмена пуриновых нуклетидов • Наследственной формой нарушения синтеза инозинмонофосфата (ИМФ) из гипоксантина и ГМФ из гуанина является болезнь Леша - Нихана • При этом заболевании, вследствие дефекта фермента гипоксантин-гуанинфосфорибозилтрансферазы, гипоксантин и гуанин не используются повторно для синтеза нуклеотидов и целиком превращаются в мочевую кислоту. • Наряду с гиперурекемией, у больных детей (мальчики) наблюдаются церебральные параличи, нарушения интеллекта, попытка наносить себе раны (укусы губ, пальцев). • Реабилитация этих больных может осуществляться использованием пиримидиновых нуклеотидов (АМФ, ГМФ, и ИМФ).
Нарушения обмена пуриновых нуклетидов • Наследственной формой нарушения синтеза инозинмонофосфата (ИМФ) из гипоксантина и ГМФ из гуанина является болезнь Леша - Нихана • При этом заболевании, вследствие дефекта фермента гипоксантин-гуанинфосфорибозилтрансферазы, гипоксантин и гуанин не используются повторно для синтеза нуклеотидов и целиком превращаются в мочевую кислоту. • Наряду с гиперурекемией, у больных детей (мальчики) наблюдаются церебральные параличи, нарушения интеллекта, попытка наносить себе раны (укусы губ, пальцев). • Реабилитация этих больных может осуществляться использованием пиримидиновых нуклеотидов (АМФ, ГМФ, и ИМФ).
 из гипоксантина и") Нарушения обмена пуриновых нуклетидов • Наследственной формой нарушения синтеза инозинмонофосфата (ИМФ) из гипоксантина и ГМФ из гуанина является болезнь Леша - Нихана • При этом заболевании, вследствие дефекта фермента гипоксантин-гуанинфосфорибозилтрансферазы, гипоксантин и гуанин не используются повторно для синтеза нуклеотидов и целиком превращаются в мочевую кислоту. • Наряду с гиперурекемией, у больных детей (мальчики) наблюдаются церебральные параличи, нарушения интеллекта, попытка наносить себе раны (укусы губ, пальцев). • Реабилитация этих больных может осуществляться использованием пиримидиновых нуклеотидов (АМФ, ГМФ, и ИМФ).
Нарушения обмена пуриновых нуклетидов • Наследственной формой нарушения синтеза инозинмонофосфата (ИМФ) из гипоксантина и ГМФ из гуанина является болезнь Леша - Нихана • При этом заболевании, вследствие дефекта фермента гипоксантин-гуанинфосфорибозилтрансферазы, гипоксантин и гуанин не используются повторно для синтеза нуклеотидов и целиком превращаются в мочевую кислоту. • Наряду с гиперурекемией, у больных детей (мальчики) наблюдаются церебральные параличи, нарушения интеллекта, попытка наносить себе раны (укусы губ, пальцев). • Реабилитация этих больных может осуществляться использованием пиримидиновых нуклеотидов (АМФ, ГМФ, и ИМФ).
 Пиримидинді нуклеотидтердің биосинтезі Пуриндік нуклеотидтерге қарағанда, пиримидиндік нуклеотидтердің биосинтезінің ерекшелігі - алдымен пиримидиндік сақинаны синтезделеді, сонан кейін пиримидиндік азоттық негіз рибозофосфатпен әрекеттесіп нуклеотид түзеді.
Пиримидинді нуклеотидтердің биосинтезі Пуриндік нуклеотидтерге қарағанда, пиримидиндік нуклеотидтердің биосинтезінің ерекшелігі - алдымен пиримидиндік сақинаны синтезделеді, сонан кейін пиримидиндік азоттық негіз рибозофосфатпен әрекеттесіп нуклеотид түзеді.
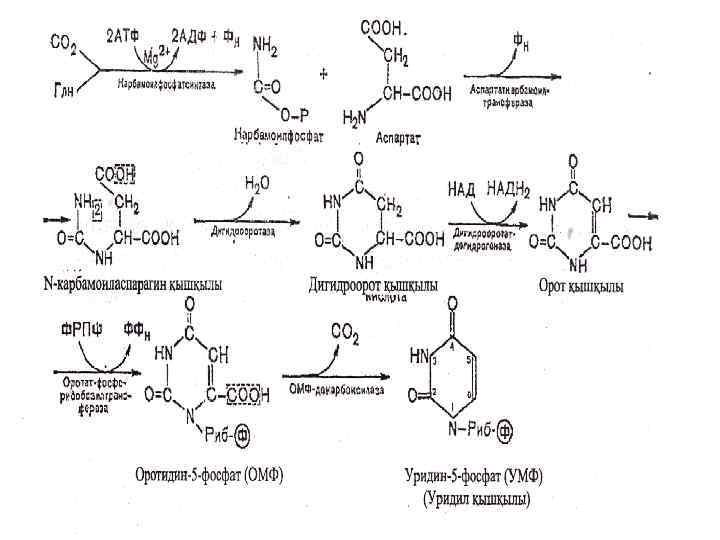
 Пиримидин нуклеотидтердің биосинтезі Пиримидин нуклеотидтердің пиридин сакинасы биосинтезі үшін глутаминның амидты тобы, СО 2 , асп пайдаланады. Синтез карбомоилфосфаттың түзулуінен басталады карбомоилфосфатсинтетаза II , цитозольде . Арығарай ---- уреидосукцинат асп, карбамоилтрансфераза ----…. оротат.
Пиримидин нуклеотидтердің биосинтезі Пиримидин нуклеотидтердің пиридин сакинасы биосинтезі үшін глутаминның амидты тобы, СО 2 , асп пайдаланады. Синтез карбомоилфосфаттың түзулуінен басталады карбомоилфосфатсинтетаза II , цитозольде . Арығарай ---- уреидосукцинат асп, карбамоилтрансфераза ----…. оротат.
 УМФ УТФ айналады УМФ + АТФ - УДФ + АДФ УДФ +АТФ УТФ +АДФ
УМФ УТФ айналады УМФ + АТФ - УДФ + АДФ УДФ +АТФ УТФ +АДФ
 УТФ-тан ЦТФ-тың синтезі жүреді:
УТФ-тан ЦТФ-тың синтезі жүреді:
 УТФ-тан ТТФ-тың синтезі жүреді: УТФ ТТФ
УТФ-тан ТТФ-тың синтезі жүреді: УТФ ТТФ
 Нуклеотидтер алмасуын реттеу * Пиримидиндік негіздер синтезі аспартаткарбомоил трансфераза арқылы реттелінеді *
Нуклеотидтер алмасуын реттеу * Пиримидиндік негіздер синтезі аспартаткарбомоил трансфераза арқылы реттелінеді *
 • Недостаток пиримидиновых нуклеотидов приводит к необратимому отставанию умственного и физического развития. • Для лечения этой болезни применяют уридин в дозах 0, 5 -1, 0 грамм в сутки. • Это обеспечивает образование УМФ и других пиримидиновых нуклеотидов в обход нарушенных реакций.
• Недостаток пиримидиновых нуклеотидов приводит к необратимому отставанию умственного и физического развития. • Для лечения этой болезни применяют уридин в дозах 0, 5 -1, 0 грамм в сутки. • Это обеспечивает образование УМФ и других пиримидиновых нуклеотидов в обход нарушенных реакций.
 • Недостаток пиримидиновых нуклеотидов приводит к необратимому отставанию умственного и физического развития. • Для лечения этой болезни применяют уридин в дозах 0, 5 -1, 0 грамм в сутки. • Это обеспечивает образование УМФ и других пиримидиновых нуклеотидов в обход нарушенных реакций.
• Недостаток пиримидиновых нуклеотидов приводит к необратимому отставанию умственного и физического развития. • Для лечения этой болезни применяют уридин в дозах 0, 5 -1, 0 грамм в сутки. • Это обеспечивает образование УМФ и других пиримидиновых нуклеотидов в обход нарушенных реакций.
 из гипоксантина и") Нарушения обмена пуриновых нуклетидов • Наследственной формой нарушения синтеза инозинмонофосфата (ИМФ) из гипоксантина и ГМФ из гуанина является болезнь Леша - Нихана • При этом заболевании, вследствие дефекта фермента гипоксантин-гуанинфосфорибозилтрансферазы, гипоксантин и гуанин не используются повторно для синтеза нуклеотидов и целиком превращаются в мочевую кислоту. • Наряду с гиперурекемией, у больных детей (мальчики) наблюдаются церебральные параличи, нарушения интеллекта, попытка наносить себе раны (укусы губ, пальцев). • Реабилитация этих больных может осуществляться использованием пиримидиновых нуклеотидов (АМФ, ГМФ, и ИМФ).
Нарушения обмена пуриновых нуклетидов • Наследственной формой нарушения синтеза инозинмонофосфата (ИМФ) из гипоксантина и ГМФ из гуанина является болезнь Леша - Нихана • При этом заболевании, вследствие дефекта фермента гипоксантин-гуанинфосфорибозилтрансферазы, гипоксантин и гуанин не используются повторно для синтеза нуклеотидов и целиком превращаются в мочевую кислоту. • Наряду с гиперурекемией, у больных детей (мальчики) наблюдаются церебральные параличи, нарушения интеллекта, попытка наносить себе раны (укусы губ, пальцев). • Реабилитация этих больных может осуществляться использованием пиримидиновых нуклеотидов (АМФ, ГМФ, и ИМФ).
 из гипоксантина и") Нарушения обмена пуриновых нуклетидов • Наследственной формой нарушения синтеза инозинмонофосфата (ИМФ) из гипоксантина и ГМФ из гуанина является болезнь Леша - Нихана • При этом заболевании, вследствие дефекта фермента гипоксантин-гуанинфосфорибозилтрансферазы, гипоксантин и гуанин не используются повторно для синтеза нуклеотидов и целиком превращаются в мочевую кислоту. • Наряду с гиперурекемией, у больных детей (мальчики) наблюдаются церебральные параличи, нарушения интеллекта, попытка наносить себе раны (укусы губ, пальцев). • Реабилитация этих больных может осуществляться использованием пиримидиновых нуклеотидов (АМФ, ГМФ, и ИМФ).
Нарушения обмена пуриновых нуклетидов • Наследственной формой нарушения синтеза инозинмонофосфата (ИМФ) из гипоксантина и ГМФ из гуанина является болезнь Леша - Нихана • При этом заболевании, вследствие дефекта фермента гипоксантин-гуанинфосфорибозилтрансферазы, гипоксантин и гуанин не используются повторно для синтеза нуклеотидов и целиком превращаются в мочевую кислоту. • Наряду с гиперурекемией, у больных детей (мальчики) наблюдаются церебральные параличи, нарушения интеллекта, попытка наносить себе раны (укусы губ, пальцев). • Реабилитация этих больных может осуществляться использованием пиримидиновых нуклеотидов (АМФ, ГМФ, и ИМФ).
 Нарушения синтеза пиримидиновых нуклеотидов • Оротацидурия. При этом заболевании вследствие недостаточности ферментов, катализирующих образование и декарбоксилирование оротидиловой кислоты накапливается оротовая кислота. • Вследствие недостатка УТФ - аллостерического ингибитора карбомоилфосфат синтетазы, еще больше усиливается синтез оротовой кислоты, которая выводится в больших количествах с мочой. • При охлаждении мочи таких больных оротовая кислота образует осадки в виде игольчатых кристаллов.
Нарушения синтеза пиримидиновых нуклеотидов • Оротацидурия. При этом заболевании вследствие недостаточности ферментов, катализирующих образование и декарбоксилирование оротидиловой кислоты накапливается оротовая кислота. • Вследствие недостатка УТФ - аллостерического ингибитора карбомоилфосфат синтетазы, еще больше усиливается синтез оротовой кислоты, которая выводится в больших количествах с мочой. • При охлаждении мочи таких больных оротовая кислота образует осадки в виде игольчатых кристаллов.
 Нарушения синтеза пиримидиновых нуклеотидов • Оротацидурия. При этом заболевании вследствие недостаточности ферментов, катализирующих образование и декарбоксилирование оротидиловой кислоты накапливается оротовая кислота. • Вследствие недостатка УТФ - аллостерического ингибитора карбомоилфосфат синтетазы, еще больше усиливается синтез оротовой кислоты, которая выводится в больших количествах с мочой. • При охлаждении мочи таких больных оротовая кислота образует осадки в виде игольчатых кристаллов.
Нарушения синтеза пиримидиновых нуклеотидов • Оротацидурия. При этом заболевании вследствие недостаточности ферментов, катализирующих образование и декарбоксилирование оротидиловой кислоты накапливается оротовая кислота. • Вследствие недостатка УТФ - аллостерического ингибитора карбомоилфосфат синтетазы, еще больше усиливается синтез оротовой кислоты, которая выводится в больших количествах с мочой. • При охлаждении мочи таких больных оротовая кислота образует осадки в виде игольчатых кристаллов.
 Пиримидиндер алмасуынің бұзылыстары Оротацидурия– оро татфосфорибозилт рансфераза, декар боксилазаның туа біткен жетіспеушілігі.
Пиримидиндер алмасуынің бұзылыстары Оротацидурия– оро татфосфорибозилт рансфераза, декар боксилазаның туа біткен жетіспеушілігі.
 Орот қ-лы ------ > оротидин -5 –фосфат----- уридил қ-лы оротат- декар- фосфо- бокси рибозил лаза трансфераза
Орот қ-лы ------ > оротидин -5 –фосфат----- уридил қ-лы оротат- декар- фосфо- бокси рибозил лаза трансфераза
 Пиримидиндер метаболизмінің бұзылыстары Нәтижесінде ағзада орот қышқылы артық мөлшерде жиналып, зәрмен шығады – ұсақ кристалдар ретінде
Пиримидиндер метаболизмінің бұзылыстары Нәтижесінде ағзада орот қышқылы артық мөлшерде жиналып, зәрмен шығады – ұсақ кристалдар ретінде
 Нәтижесінде пиридинді нуклеотидтердің жетіспеушілігі ақыл - ойдың кемістігіне және физикалық дамудын төмендеуне әкеледі.
Нәтижесінде пиридинді нуклеотидтердің жетіспеушілігі ақыл - ойдың кемістігіне және физикалық дамудын төмендеуне әкеледі.
 Пиримидинді нуклеотидтердің ыдырауы
Пиримидинді нуклеотидтердің ыдырауы
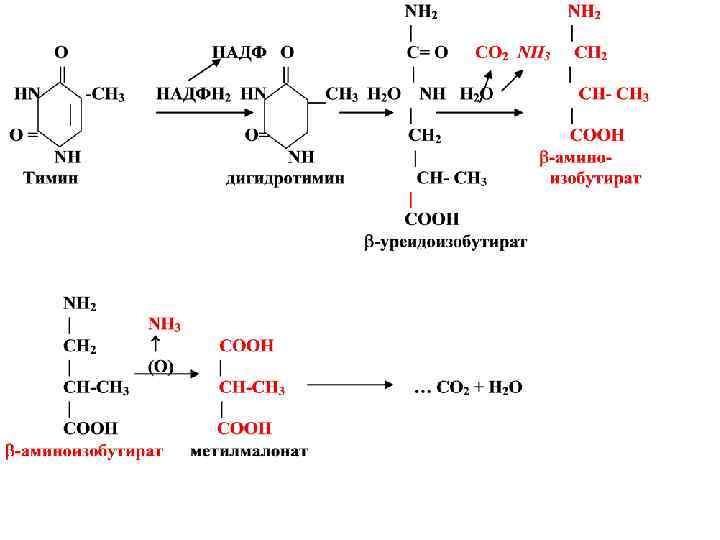
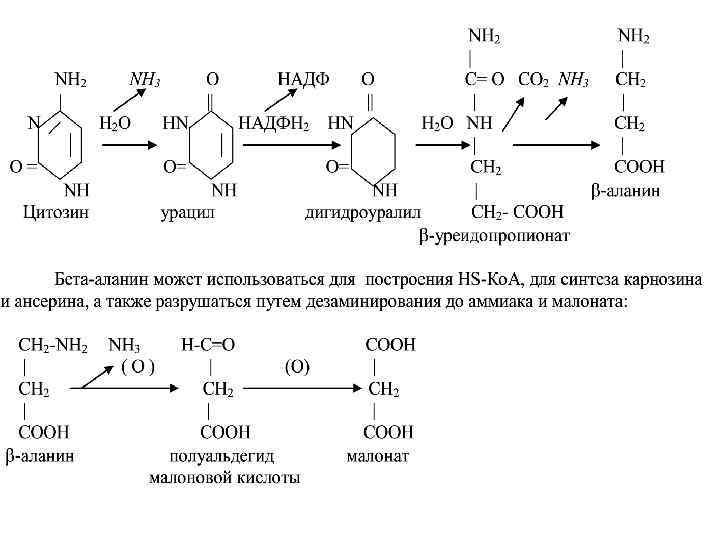
 • Катаболизм пиримидиновых нуклеотидов Пиримидиновые нуклеотиды соответствующими клеточными ферментами, гидролитически разрушаются до нуклеозидов, фосфорной кислоты, затем до азотистых оснований и пентоз (рибоза или дезоксирибоза). • Далее идет специфический путь катаболизма пиримидиновых оснований. • Цитозин дезаминируется с образованием урацила и аммиака. Урацил восстанавливается до дигидроурацила. • Затем происходит гидролитическая дециклизация дигидроурацила и образуется бетауридопропионат, который гидролизуется с образованием СО 2, аммиака и бета-аланина
• Катаболизм пиримидиновых нуклеотидов Пиримидиновые нуклеотиды соответствующими клеточными ферментами, гидролитически разрушаются до нуклеозидов, фосфорной кислоты, затем до азотистых оснований и пентоз (рибоза или дезоксирибоза). • Далее идет специфический путь катаболизма пиримидиновых оснований. • Цитозин дезаминируется с образованием урацила и аммиака. Урацил восстанавливается до дигидроурацила. • Затем происходит гидролитическая дециклизация дигидроурацила и образуется бетауридопропионат, который гидролизуется с образованием СО 2, аммиака и бета-аланина
 • Катаболизм пиримидиновых нуклеотидов Пиримидиновые нуклеотиды соответствующими клеточными ферментами, гидролитически разрушаются до нуклеозидов, фосфорной кислоты, затем до азотистых оснований и пентоз (рибоза или дезоксирибоза). • Далее идет специфический путь катаболизма пиримидиновых оснований. • Цитозин дезаминируется с образованием урацила и аммиака. Урацил восстанавливается до дигидроурацила. • Затем происходит гидролитическая дециклизация дигидроурацила и образуется бетауридопропионат, который гидролизуется с образованием СО 2, аммиака и бета-аланина
• Катаболизм пиримидиновых нуклеотидов Пиримидиновые нуклеотиды соответствующими клеточными ферментами, гидролитически разрушаются до нуклеозидов, фосфорной кислоты, затем до азотистых оснований и пентоз (рибоза или дезоксирибоза). • Далее идет специфический путь катаболизма пиримидиновых оснований. • Цитозин дезаминируется с образованием урацила и аммиака. Урацил восстанавливается до дигидроурацила. • Затем происходит гидролитическая дециклизация дигидроурацила и образуется бетауридопропионат, который гидролизуется с образованием СО 2, аммиака и бета-аланина



 β-аланин ансерин және карнозин дипептидтер бұлшық еттің құрамына кіреді.
β-аланин ансерин және карнозин дипептидтер бұлшық еттің құрамына кіреді.


 Пиримидиндер метаболизмінің бұзылыстары Туа біткен оротацидурия – оротатфосфорибозилтрансферазан ың және декарбоксилазаның белсенділігінің төмендеуі нормамен салыстырғанда 1 -5 % . Нәтижесінде ағзада орот қышқылы артық мөлшерде жиналып, зәрмен шығады – ұсақ кристалдар ретінде
Пиримидиндер метаболизмінің бұзылыстары Туа біткен оротацидурия – оротатфосфорибозилтрансферазан ың және декарбоксилазаның белсенділігінің төмендеуі нормамен салыстырғанда 1 -5 % . Нәтижесінде ағзада орот қышқылы артық мөлшерде жиналып, зәрмен шығады – ұсақ кристалдар ретінде
 Нуклеотидтер алмасуын реттеу * Пиримидиндік негіздер синтезі аспартаткарбомоил трансфераза арқылы реттелінеді *
Нуклеотидтер алмасуын реттеу * Пиримидиндік негіздер синтезі аспартаткарбомоил трансфераза арқылы реттелінеді *
 РЕФЛЕКСИВТЫ ТЕСТ • Рефлексия - субъектін өз өзіне, өз тұлғасына • /құндылықтар, қізіушіліқтар, се бептер, эмоциялар, қылықтары , • өз біліміне немесе өзінін меншікті жағдайына назар аудару
РЕФЛЕКСИВТЫ ТЕСТ • Рефлексия - субъектін өз өзіне, өз тұлғасына • /құндылықтар, қізіушіліқтар, се бептер, эмоциялар, қылықтары , • өз біліміне немесе өзінін меншікті жағдайына назар аудару
 • Мысалы; • Көп жаналықтарды білдім • Бұл маған өмірде керек болады • Туындаған сұрақтарға жауап алдым. Адал ниетпен жұмыс істеп, мақсатыма жеттім
• Мысалы; • Көп жаналықтарды білдім • Бұл маған өмірде керек болады • Туындаған сұрақтарға жауап алдым. Адал ниетпен жұмыс істеп, мақсатыма жеттім
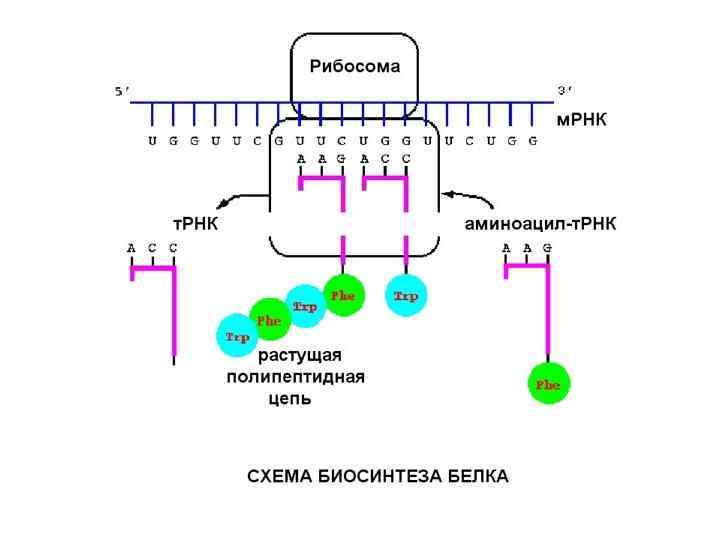
 Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул м. РНК и т. РНК
Биосинтез белка — сложный многостадийный процесс синтеза полипептидной цепи из аминокислотных остатков, происходящий на рибосомах клеток живых организмов с участием молекул м. РНК и т. РНК
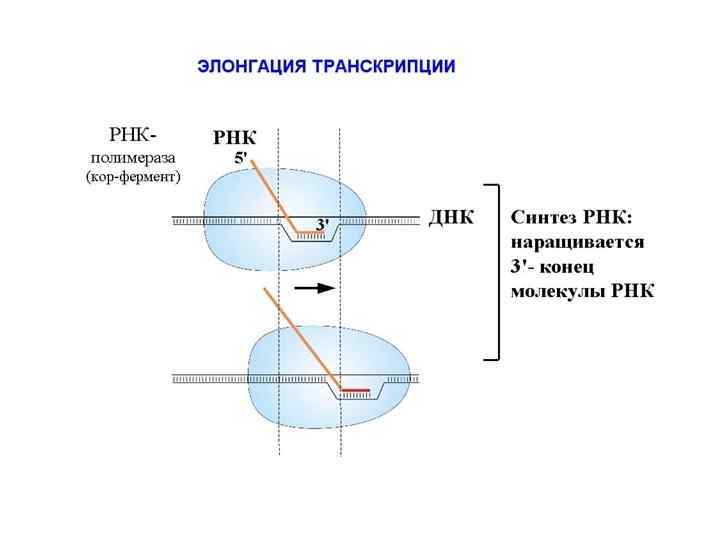
, синтез РНК – транскрипцией (переписывание с ДНК),") Синтез ДНК называется репликацией или редупликацией (удвоением), синтез РНК – транскрипцией (переписывание с ДНК), синтез белка, проводимый рибосомой на матричной РНК называется трансляцией, то есть переводим с языка нуклеотидов на язык аминокислот.
Синтез ДНК называется репликацией или редупликацией (удвоением), синтез РНК – транскрипцией (переписывание с ДНК), синтез белка, проводимый рибосомой на матричной РНК называется трансляцией, то есть переводим с языка нуклеотидов на язык аминокислот.
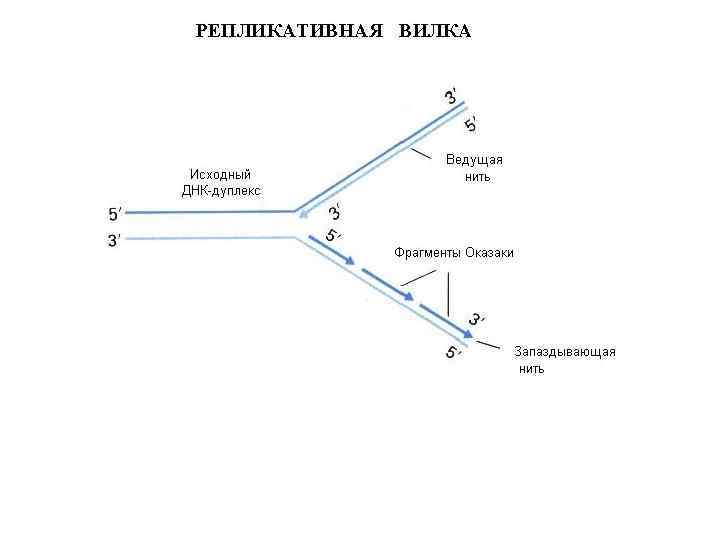
 Репликация ДНК Молекула ДНК, состоящая из двух спиралей, удваивается при делении клетки. Удвоение ДНК основано на том, что при расплетении нитей к каждой нити можно достроить комплементарную копию, таким образом получая две нити молекулы ДНК, копирующие исходную.
Репликация ДНК Молекула ДНК, состоящая из двух спиралей, удваивается при делении клетки. Удвоение ДНК основано на том, что при расплетении нитей к каждой нити можно достроить комплементарную копию, таким образом получая две нити молекулы ДНК, копирующие исходную.

 Трансляция Перейдем к трансляции – синтезу белков. Она проводится рибосомами. Рибосома состоит из двух субчастиц: большой и малой.
Трансляция Перейдем к трансляции – синтезу белков. Она проводится рибосомами. Рибосома состоит из двух субчастиц: большой и малой.
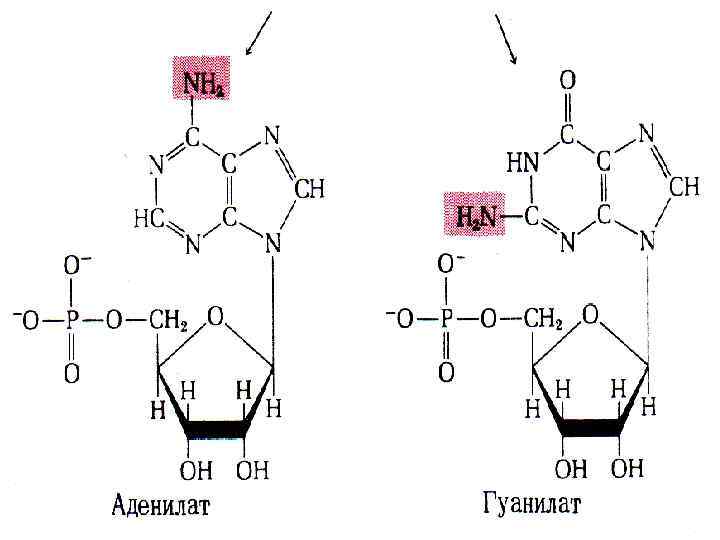
 2 -гипоксантин-гуанин-фосфорибозилтрасфераза (инозин мен гуанил қышқылын түзуге") 1 -аденинфосфорибозилтрансфераза (аденил қышқылының түзілуін қамтамасыз етеді) 2 -гипоксантин-гуанин-фосфорибозилтрасфераза (инозин мен гуанил қышқылын түзуге қатысады) Гипоксантин+ФРПФ инозинат + H 4 P 2 O 7 Гуанин+ ФРПФ гуанилат+ H 4 P 2 O 7
1 -аденинфосфорибозилтрансфераза (аденил қышқылының түзілуін қамтамасыз етеді) 2 -гипоксантин-гуанин-фосфорибозилтрасфераза (инозин мен гуанил қышқылын түзуге қатысады) Гипоксантин+ФРПФ инозинат + H 4 P 2 O 7 Гуанин+ ФРПФ гуанилат+ H 4 P 2 O 7
 Сызбанұсқа түрінде: Глюкоза глюкоза-6 -фосфат рибоза-5 -фосфат АТФ Mg 2+ ФРПФ Глн Глу амидотрансфераза 5 -фосфорибозил-1 -амин Осы жолмен түзілген 5 -фосфорибозиламин глицин, аспартат, глу-, СО 2, формил-ТГФҚ қышқылдарының қатысуымен нуклеотид түзілуіне қатынасады:
Сызбанұсқа түрінде: Глюкоза глюкоза-6 -фосфат рибоза-5 -фосфат АТФ Mg 2+ ФРПФ Глн Глу амидотрансфераза 5 -фосфорибозил-1 -амин Осы жолмен түзілген 5 -фосфорибозиламин глицин, аспартат, глу-, СО 2, формил-ТГФҚ қышқылдарының қатысуымен нуклеотид түзілуіне қатынасады:
 Пуринді негіздердің синтезделуінің қосымша жолдары: • Аденозин+АТФ АМФ + АДФ; • АМФ+АТФ АТФ+АДФ; • А+ФРПФ АМФ+H 4 P 2 O 7
Пуринді негіздердің синтезделуінің қосымша жолдары: • Аденозин+АТФ АМФ + АДФ; • АМФ+АТФ АТФ+АДФ; • А+ФРПФ АМФ+H 4 P 2 O 7
 Пиримидинді нуклеотид биосинтезі • Ерекшеліктері: Алдымен пиримидиндік сақинаның синтезі жүріп, сонан кейін пиримидиндік азоттық негіз рибозофосфатпен әрекеттесіп нуклеотид түзеді Синтез-карбомоилфосфаттың түзілуінен басталады. Карбомоилфосфат мочевинаның да синтезі үшін реакцияаралық зат, бірақ та пиримидин синтезіне жұмсалатын карбомоилфосфат клетка цитозолінде синтезделсе, мочевина үшін митохондрияде синтезделеді. Амин қышқылының доноры глутамин қышқылының амиді болып табылады:
Пиримидинді нуклеотид биосинтезі • Ерекшеліктері: Алдымен пиримидиндік сақинаның синтезі жүріп, сонан кейін пиримидиндік азоттық негіз рибозофосфатпен әрекеттесіп нуклеотид түзеді Синтез-карбомоилфосфаттың түзілуінен басталады. Карбомоилфосфат мочевинаның да синтезі үшін реакцияаралық зат, бірақ та пиримидин синтезіне жұмсалатын карбомоилфосфат клетка цитозолінде синтезделсе, мочевина үшін митохондрияде синтезделеді. Амин қышқылының доноры глутамин қышқылының амиді болып табылады:
 ФРПФ-трансфераза, оротидилатдекарбоксилаза белсенділігі төмен болса оротацидурия дамиды-орот қышқылының кристалдарымен несеп шығару жолдарының бітелуі, несеп жүруінің нашарлауы УМФ УТФ қа АТФ қатысуымен айналады УМФ+АТФ- УДФ+АДФ УДФ+АТФ- УТФ+АДФ УТФ-тан ЦТФ-тың және ТТФ-тың синтезі жүреді: УТФ ТТФ
ФРПФ-трансфераза, оротидилатдекарбоксилаза белсенділігі төмен болса оротацидурия дамиды-орот қышқылының кристалдарымен несеп шығару жолдарының бітелуі, несеп жүруінің нашарлауы УМФ УТФ қа АТФ қатысуымен айналады УМФ+АТФ- УДФ+АДФ УДФ+АТФ- УТФ+АДФ УТФ-тан ЦТФ-тың және ТТФ-тың синтезі жүреді: УТФ ТТФ
 Пиримидинді нуклеотидтердің биосинтезі Пуриндік нуклеотидтерге қарағанда, пиримидиндік нуклеотидтердің биосинтезінің ерекшелігі - алдымен пиримидиндік сақинаны синтезделеді, сонан кейін пиримидиндік азоттық негіз рибозофосфатпен әрекеттесіп нуклеотид түзеді.
Пиримидинді нуклеотидтердің биосинтезі Пуриндік нуклеотидтерге қарағанда, пиримидиндік нуклеотидтердің биосинтезінің ерекшелігі - алдымен пиримидиндік сақинаны синтезделеді, сонан кейін пиримидиндік азоттық негіз рибозофосфатпен әрекеттесіп нуклеотид түзеді.
 Пиримидиндік азоттық негіздің синтезі карбомоилфосфаттың түзілуінен басталады. Карбомоилфосфат мочевинаның да синтезі үшін реакция аралық зат, бірақта пиримидин синтезіне жұмсалатын карбомоилфосфат клетка цитозолінде синтезделсе, мочевина үшін митохондриде синтезделеді. Амин тобының доноры глутамин қышқылының амиді болып табылады. О Глн+СО 2 +2 АТФ + Н 2 О NН 2 С~О-РО 3 Н 2 + 2 АДФ + Н 3 РО 4
Пиримидиндік азоттық негіздің синтезі карбомоилфосфаттың түзілуінен басталады. Карбомоилфосфат мочевинаның да синтезі үшін реакция аралық зат, бірақта пиримидин синтезіне жұмсалатын карбомоилфосфат клетка цитозолінде синтезделсе, мочевина үшін митохондриде синтезделеді. Амин тобының доноры глутамин қышқылының амиді болып табылады. О Глн+СО 2 +2 АТФ + Н 2 О NН 2 С~О-РО 3 Н 2 + 2 АДФ + Н 3 РО 4
 Пиримидиндік азоттық негіздердің синтезінде шешуші кезең карбомоилфосфат пен аспартаттың қатысуымен N-карбомоиласпартаттың түзілуі:
Пиримидиндік азоттық негіздердің синтезінде шешуші кезең карбомоилфосфат пен аспартаттың қатысуымен N-карбомоиласпартаттың түзілуі:
 Осы кезеңдегі ФРПФ-трансфераза және оротидилатдекарбоксилаза ферметтерінің белсенділігі төмен болса оротацидурия ауруы дамиды. Бұл кезде орот қышқылының кристалдарымен несеп шығару жолдарының бітелуі, несеп жүруінің нашарлауы байқалады. Жас нәрестелердің өсуі тоқтап, шаштары сирек, тырнақтары нашар өседі, көкбауырдың үлкеюі байқалады. Мұндай ауруларды уридинді дәрімен емдейді. Бұл жолмен емдегенде шаштың өсуі, көкбауыр қайта қалпына келгенімен ақыл-ойдың кемістігі қалпына келмейді. Уридилмонофосфар қыщқылы уридилтрифосфатқа АТФ-тың қатысуымен айналады: УМФ + АТФ УДФ + АДФ УДФ + АТФ УТФ + АДФ
Осы кезеңдегі ФРПФ-трансфераза және оротидилатдекарбоксилаза ферметтерінің белсенділігі төмен болса оротацидурия ауруы дамиды. Бұл кезде орот қышқылының кристалдарымен несеп шығару жолдарының бітелуі, несеп жүруінің нашарлауы байқалады. Жас нәрестелердің өсуі тоқтап, шаштары сирек, тырнақтары нашар өседі, көкбауырдың үлкеюі байқалады. Мұндай ауруларды уридинді дәрімен емдейді. Бұл жолмен емдегенде шаштың өсуі, көкбауыр қайта қалпына келгенімен ақыл-ойдың кемістігі қалпына келмейді. Уридилмонофосфар қыщқылы уридилтрифосфатқа АТФ-тың қатысуымен айналады: УМФ + АТФ УДФ + АДФ УДФ + АТФ УТФ + АДФ
 Сонан соң ғана, УТФ-тан ЦТФ-тің және ТТФ-тің синтезі жүреді:
Сонан соң ғана, УТФ-тан ЦТФ-тің және ТТФ-тің синтезі жүреді:
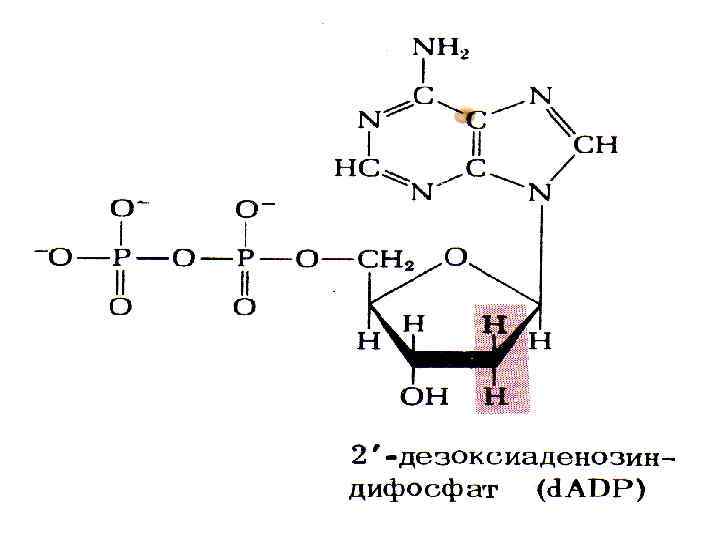
 Дезоксирибонуклеотидтердің биосинтезі Дезоксирибонуклеотидтердің құрамындағы 2‘-орналасқан. ОН тобының тотықсыздануы арқылы синтезделеді. Бұл реакция үшін негізінен рибонуклеозиддифосфат жұмсалады. Реакция сызбасы: Рибонуклеозиддифосфат+НАДФНН+ дезоксирибонуклеозид+НАДФ+Н 2 О Бұл реакция механизмі көрсетілген сызбанұсқаға қарағанда күрделірек. Себебі, электрондардың тасымалдануы кейбір ақуыз құрамында болатын сульфгидрилдік топ арқасында жүреді. Осы ақуыздардың бірі-тиоредоксин.
Дезоксирибонуклеотидтердің биосинтезі Дезоксирибонуклеотидтердің құрамындағы 2‘-орналасқан. ОН тобының тотықсыздануы арқылы синтезделеді. Бұл реакция үшін негізінен рибонуклеозиддифосфат жұмсалады. Реакция сызбасы: Рибонуклеозиддифосфат+НАДФНН+ дезоксирибонуклеозид+НАДФ+Н 2 О Бұл реакция механизмі көрсетілген сызбанұсқаға қарағанда күрделірек. Себебі, электрондардың тасымалдануы кейбір ақуыз құрамында болатын сульфгидрилдік топ арқасында жүреді. Осы ақуыздардың бірі-тиоредоксин.
 Цистеиндегі сульфгидрилдік топтар редуктаза ферментінің әсерінен тотығып дисульфидтік байланыс түзеді. Осы тотыққан тиоредоксин НАДФНН+ әрекеттесіп, қайтадан өзінің тотықсызданған қалпына келеді. Реакция сызбанұсқасы: Осы түзілген дезоксирибонуклеозиддифосфаттан дезоксирибонуклеозидтрифосфат түзіледі. d-ГДФ+d-АТФ d-ГТФ+d-АДФ
Цистеиндегі сульфгидрилдік топтар редуктаза ферментінің әсерінен тотығып дисульфидтік байланыс түзеді. Осы тотыққан тиоредоксин НАДФНН+ әрекеттесіп, қайтадан өзінің тотықсызданған қалпына келеді. Реакция сызбанұсқасы: Осы түзілген дезоксирибонуклеозиддифосфаттан дезоксирибонуклеозидтрифосфат түзіледі. d-ГДФ+d-АТФ d-ГТФ+d-АДФ
 Нуклеотидтер алмасуын реттеу *Пуриндік негіздер синтезі қайта айналып реттеу механизмі арқылы реттеледі. Ең бірінші реттеуші фермент амидофосфорибозилтрансфераза, бұл синтезді ең бастапқы кезеніңде, яғни 5 -фосфорибозил-1 -пирофосфаттан 5 фосфорибозиламиннің синтезі кезінде реттеліп отырады. *Пиримидиндік негіздер синтезі аспартаткарбомоилтрансфераза ферменті қызметі арқылы реттеледі. Бұл ферменттің аллостерлік реттеушісі ЦТФ. *Нуклеотидтердің алмасуының бұзылуы көптеген ауруға әкеліп соғады. Мысалы-фосфорибозилрансфераза жетіспеуі Леше. Нихан ауруына әкеп соғады.
Нуклеотидтер алмасуын реттеу *Пуриндік негіздер синтезі қайта айналып реттеу механизмі арқылы реттеледі. Ең бірінші реттеуші фермент амидофосфорибозилтрансфераза, бұл синтезді ең бастапқы кезеніңде, яғни 5 -фосфорибозил-1 -пирофосфаттан 5 фосфорибозиламиннің синтезі кезінде реттеліп отырады. *Пиримидиндік негіздер синтезі аспартаткарбомоилтрансфераза ферменті қызметі арқылы реттеледі. Бұл ферменттің аллостерлік реттеушісі ЦТФ. *Нуклеотидтердің алмасуының бұзылуы көптеген ауруға әкеліп соғады. Мысалы-фосфорибозилрансфераза жетіспеуі Леше. Нихан ауруына әкеп соғады.
 Дезоксиметил қышқылы дезоксиуридил қышқылынан N 5, N 10 -ТГФҚ арқасында метилдену арқылы түзіледі. Дезоксицитидил қышқылы да дезоксиуридил қышқылынан –NH 2 тобын қосып алу арқылы түзіледі. d-УДФ d-ц. ДФ Глн Глу
Дезоксиметил қышқылы дезоксиуридил қышқылынан N 5, N 10 -ТГФҚ арқасында метилдену арқылы түзіледі. Дезоксицитидил қышқылы да дезоксиуридил қышқылынан –NH 2 тобын қосып алу арқылы түзіледі. d-УДФ d-ц. ДФ Глн Глу
 .
.

 Аденофосфор қышқылы мен гуанизофосфор қышқылының биосинтезі:
Аденофосфор қышқылы мен гуанизофосфор қышқылының биосинтезі:
 Основные функции нуклеотидов
Основные функции нуклеотидов
 Нуклеотидтер алмасуын реттеу *Пуриндік негіздер синтезі қайта айналып реттеу механизмі арқылы реттеледі. Ең бірінші реттеуші фермент амидофосфорибозилтрансфераза, бұл синтезді ең бастапқы кезеніңде, яғни 5 -фосфорибозил-1 -пирофосфаттан 5 -фосфорибозиламиннің синтезі кезінде реттеп отырады. *Пиримидиндік негіздер синтезі аспартаткарбомоилтрансфераза арқылы реттеледі. Бұл ферменттің аллостерлік реттеушісі ЦТФ. *Нуклеотидтердің алмасуының бұзылуы көптеген ауруға әкеліп соғады. Мысалы-фосфорибозилрансфераза жетіспеуі Леше-Нихан ауруына әкеп соғады.
Нуклеотидтер алмасуын реттеу *Пуриндік негіздер синтезі қайта айналып реттеу механизмі арқылы реттеледі. Ең бірінші реттеуші фермент амидофосфорибозилтрансфераза, бұл синтезді ең бастапқы кезеніңде, яғни 5 -фосфорибозил-1 -пирофосфаттан 5 -фосфорибозиламиннің синтезі кезінде реттеп отырады. *Пиримидиндік негіздер синтезі аспартаткарбомоилтрансфераза арқылы реттеледі. Бұл ферменттің аллостерлік реттеушісі ЦТФ. *Нуклеотидтердің алмасуының бұзылуы көптеген ауруға әкеліп соғады. Мысалы-фосфорибозилрансфераза жетіспеуі Леше-Нихан ауруына әкеп соғады.