Презентация (нуклеиновые кислоты).ppt
- Количество слайдов: 58



НУКЛЕИНОВЫЕ КИСЛОТЫ Строение и биосинтез ДНК и РНК

Нуклеиновые кислоты выполняют в организме ряд важнейших функций: они обеспечивают хранение и передачу генетической информации и они же участвуют в механизмах, при помощи которых эта информация реализуется в процессе синтеза клеточных белков. Нуклеиновые кислоты являются составной частью сложных белков - нуклеопротеинов, содержащихся во всех клетках животных, растений, бактерий, вирусов. Нуклеиновые кислоты обладают сильно выраженными кислотными свойствами (обусловлены остатками фосфорной кислоты в их составе) и при физиологических значениях р. Н несут отрицательный заряд. Этим объясняется одно из важных свойств нуклеиновых кислот – способность к взаимодействия по типу ионной связи с основными белками (гистонами), ионами металлов (преимущественно с Mg 2+) и полиаминами.

Поэтому для выделения нуклеиновых кислот из комплексов с белками необходимо прежде всего разрушить эти сильные электростатические связи между положительно заряженными молекулами белков и отрицательно заряженными молекулами нуклеиновых кислот. Для этого измельченный биоматериал обрабатывают крепкими солевыми растворами (10% раствор хлорида натрия) с последующим осаждением нуклеиновых кислот этанолом. В настоящее время для выделения нуклеиновых кислот используют фенольный метод. Нуклеопротеины обрабатывают фенолом, при этом белок взаимодействует с фенолом и выпадает в осадок. Нуклеиновые кислоты остаются в водной среде, из которой их осаждают на холоде добавлением этанола.

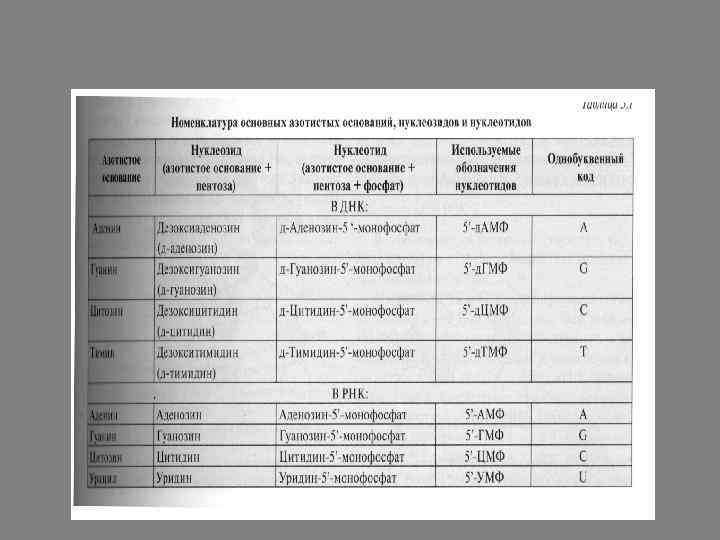
ДНК и РНК представляют собой линейные полимеры, построенные из нуклеотидов. Каждый нуклеотид в свою очередь состоит из трех компонентов: азотистого основания, являющегося производным пурина или пиримидина, пентозы (рибозы или дезоксирибозы) и остатка фосфорной кислоты. В состав нуклеиновый кислот входят два производных пурина – аденин и гуанин и три производных пиримидина – цитозин, урацил (в РНК) и тимин (в ДНК).

Производные пурина и пиримидина

В нуклеиновые кислоты входят два вида пентоз: β-D-рибоза в РНК и β-D-2 -дезоксирибоза в молекулу ДНК.

Углеродный атом в положении 1 пентозы связывается Nгликозидной связью с атомом азота в положении 1 пиримидина или в положении 9 пурина. Образующиеся соединения называют нуклеозидами. Атомы пентоз, в отличие от атомов азотистых оснований, обозначают номерами со штрихом (1’, 2’, 3’, 4’, 5’). Присоединение фосфата в положении 5’ пентоз приводит к образованию нуклеотидов.

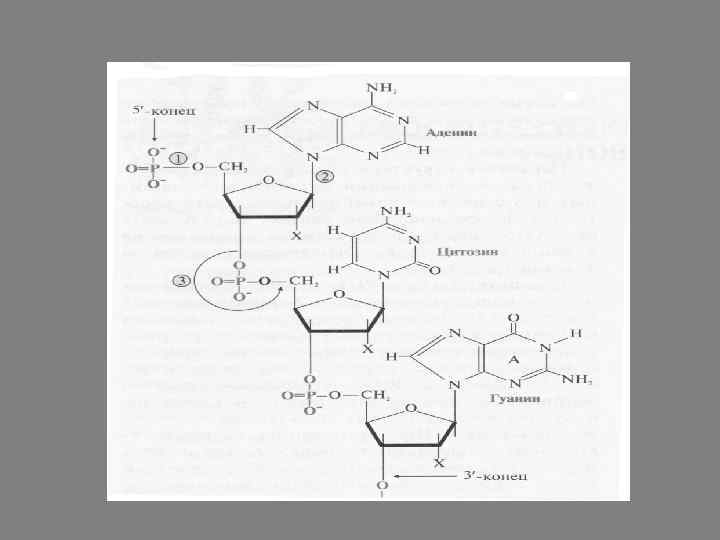
Молекулы полинуклеотидов имеют нитевидную структуру. В основе этих нитей лежит однообразно повторяющаяся последовательность из пентозы и остатка фосфорной кислоты, а азотистые основания, подобно радикалам аминокислот в полипептиде, находятся на внешней части цепей, где и выполняют свои основные функции.

Первичная структура нуклеиновых кислот – это порядок чередования нуклеотидов, связанных друг с другом в линейной последовательности 3’, 5’-фосфодиэфирной связью. В результате образуются полимеры с фосфатным остатком на 5’-конце и свободной – ОН- группой пентозы на 3’- конце. 1 - 5’-фосфоэфирная связь 2 - N-гликозидная связь 3 - 3’, 5’-фосфодиэфирная связь

В состав ДНК и РНК входит 4 основных нуклеотида: 2 пуриновых и 2 пиримидиновых. Однако если азотистые основания в пуриновых нуклеотидах – аденин и гуанин – у них одинаковы, то в пиримидиновых совпадает только цитозин, а второе основание различно: в ДНК – тимин, а в РНК – урацил. Для краткого изображения последовательности нуклеотидов в нуклеиновых кислотах пользуются однобуквенным кодом. При этом запись осуществляют слева направо таким образом, что первый нуклеотид имеет свободный 5’-фосфатный конец, а последний – ОН-групу в положении 3’ рибозы или дезоксирибозы.

Если полинуклеотидная цепь имеет противоположное направление, то в этом случае обязательно указывается направление цепей от 5’- к 3’ или от 3’- к 5’- концу. В каждой нуклеиновой кислоте присутствует остаток фосфорной кислоты. При р. Н 7 фосфатная группа полностью ионизирована и поэтому нуклеиновые кислоты имеют отрицательный заряд. Остатки пентоз тоже проявляют гидрофильные свойства. Азотистые основания нерастворимы в воде.

Вторичная структура ДНК В 1953 году Уотсоном и Криком была предложена модель пространственной структуры ДНК. Согласно этой модели, вторичная структура ДНК представлена двойной правозакрученной спиралью

В двойной спирали две полинуклеотидные цепи расположены антипараллельно (одна в направлении 3’к 5’, вторая в направлении 5’ к 3’) и удерживаются относительно друга за счет взаимодействия между комплементарными азотистыми основаниями.

Полинуклеотидные цепи молекулы ДНК комплементарны другу и соединяются между собой при помощи водородных связей. Следовательно, если известна первичная структура одной цепи, то последовательность нуклеотидов другой цепи задается правилом комплементарности оснований: Аденин одной цепи соответствует Тимину – другой, Гуанин соответствует - Цитозину. Поэтому в молекуле ДНК количество адениловых нуклеотидов равно количеству тимидиловых нуклеотидов (А=Т), а количество гуаниловых – количеству цитидиловых (С=G).
. Комплементарные")
Соотношение А+Т/ G+С – величина постоянная и является видоспецифической характеристикой организма (правило Чаргаффа). Комплементарные основания обращены внутрь молекулы, лежат в одной плоскости, которая практически перпендикулярна оси спирали. В результате образуется стопка оснований, между которыми возникают гидрофобные взаимодействия, обеспечивающие основной вклад в стабилизацию структуры спирали. Остатки фосфорной кислоты, несущие одинаковый заряд, отталкиваются друг от друга и также вносят определенный вклад в образование пространственной структуры.

Третичная структура ДНК Формируется при ее взаимодействии с белками. Каждая молекула ДНК упакована в отдельную хромосому, в составе которой разнообразные белки связываются с отдельными участками ДНК и обеспечивают суперспирализацию и компактизацию молекулы. Общая длина ДНК гаплоидного набора из 23 хромосом человека составляет 3, 5 х109 пар нуклеотидов. Хромосомы образуют компактные структуры только в фазу деления.

В период покоя комплексы ДНК с белками равномерно распределены по объему ядра, образуя хроматин. Белки хроматина делят на две группы: гистоны и негистоновые белки. Гистоны - это небольшие белки с высоким содержанием положительно заряженных аминокислот лизина и аргинина. Они взаимодействуют с отрицательно заряженными фосфатными группами ДНК длиной 146 нуклеотидных пар, образуя нуклеосомы. Между нуклеосомами находится участок ДНК, включающий около 30 нуклеотидных пар – линкерный участок, к которому также присоединяется молекула гистона.

Нуклеосома состоит из участка ДНК длиной около 146 нуклеотидных пар и 8 молекул гистонов 4 видов (Н 2 А, Н 2 В, Н 3, Н 4), которые в количестве по два каждого вида образуют комплекс – нуклеосомный кор. ДНК и белки удерживаются друг с другом за счет ионных связей. Линкерный участок ДНК связан с гистоном Н 1. Негистоновые белки представлены разными ферментами и белками (факторы транскрипции, полимеразы, гормональные рецепторы), участвующими в синтезе ДНК и РНК, в регуляции этих процессов, а также структурными белками, обеспечивающими компактизацию ДНК.

ПРОСТРАНСТВЕННАЯ СТРУКТУРА РНК Первичная структура РНК – порядок чередования рибонуклеотидов в полинуклеотидной цепи. В РНК также как и в ДНК, нуклеотиды связанны между собой 3’, 5’- фосфодиэфирными связями. Концы полинуклеотидных цепей РНК неодинаковы. На одном конце находится фосфорилированная группа 5’-углеродного атома, на другом конце - ОНгруппа 3’-углеродного атома рибозы, поэтому концы называют 5’- и 3’- концами цепи РНК. ОН-

Вторичная структура РНК формируется в результате спирализации отдельных участков одноцепочечной РНК. В спирализованных участках или шпильках комплементарные пары азотистых оснований аденин и урацил, гуанин и цитозин соединяются водородными связями. Спирализованные участки, состоящие от 20 до 30 нуклеотидных пар, чередуются с неспирализованными участками. Третичная структура РНК формируется за счет образования дополнительных водородных связей между нуклеотидами, полинуклеотидной цепью и белками. стабилизируется ионами Mg 2+ и обеспечивает дополнительную компактизацию и стабилизацию пространственной структуры молекулы.

Основные виды РНК В зависимости от первичной структуры, размера молекул и их функций в клетках выделяют три основных вида РНК. Матричные РНК (м. РНК) или информационные РНК составляют 2 -4% всей РНК клетки. Они чрезвычайно разнообразны по первичной структуре, и их количество также велико, как и белков в организме. Каждая м. РНК является матрицей в синтезе соответствующего белка.
 являются молекулами адапторами, у которых к 3’-концу присоединяется аминокислота, а")
Транспортные РНК (т. РНК) являются молекулами адапторами, у которых к 3’-концу присоединяется аминокислота, а участок антикодона – к м. РНК. Семейство т. РНК включает более 30 различных по первичной структуре молекул, состоящих примерно из 80 нуклеотидов. Особенностью т. РНК является содержание 10 -20 % модифицированных нуклеотидов, в состав которых входят метилированные или восстановленные азотистые основания, нуклеотиды с С-С связью между азотистым основанием и рибозой.

Вторичная структура т. РНК описывается как структура клеверного листа. На долю т. РНК приходится 15% всей РНК клетки. 1 -шпильки (нити РНК в этих участках антипараллельны) 2 -петли образованы участками одноцепочечной РНК
 составляют около 80% всей РНК в клетке и входят в")
Рибосомные РНК (р. РНК) составляют около 80% всей РНК в клетке и входят в состав рибосом. В цитоплазматические рибосомы эукариот входит 4 типа р. РНК с разной константой седиментации – скоростью оседания в ультрацентрифуге. (5 S, 5, 8 S, 28 S и 18 S). Большая и малая субъединицы рибосомы образуют комплекс (80 S). Митохондриальные рибосомы значительно меньше цитоплазматических рибосом и их структура сходна со структурой рибосом прокариотов.
 состоит из 50 S и 30 S субъединиц. 50 S")
Рибосома прокариот (70 S) состоит из 50 S и 30 S субъединиц. 50 S субъединица содержит 5 S, 23 S и 34 молекулы белков 30 S субъединица содержит 16 S и 21 молекулу белка Рибосома эукариот (80 S) состоит из 60 S и 40 S субъединиц. 60 S субъединица содержит 5, 8 S, 28 S, 5 S р. РНК и 49 молекул белков 40 S субъединица содержит 18 S и 33 молекулы белков Рибосомы эукариот и прокариот различаются по молекулярной массе субъединиц, количеству и разнообразию белков
 Синтез ДНК протекает в ядре в S-фазу клеточного цикла и предшествует")
БИОСИНТЕЗ ДНК (РЕПЛИКАЦИЯ) Синтез ДНК протекает в ядре в S-фазу клеточного цикла и предшествует делению клеток. Первоначально клетка из состояния покоя Go вступает в G 1 -фазу, в ходе которой синтезируются ферменты и белки, необходимые для синтеза ДНК. Затем в S-фазу протекает репликация и диплоидная клетка превращается в тетраплоидную, а в ходе митоза делится, образуя 2 дочерние диплоидные клетки.

В эукариотических клетках репликация начинается одновременно во многих участках ДНК, которые имеют специфическую нуклеотидную последовательность и называются ориджинами репликации. От каждого ориджина синтез новых цепей ДНК идет в двух противоположных направлениях, образуя две репликативные вилки. Процесс является полуконсервативным, так как по завершении репликации каждая дочерняя молекула ДНК содержит одну родительскую нить и одну вновь синтезированную.

Матрицей служат обе нити ДНК. Субстратами являются 4 дезоксирибонуклеотидтрифосфата – д. АТФ, д. ТТФ, д. ЦТФ, д. ГТФ. Они служат субстратами синтеза и донорами энергии, так как содержат богатые энергией связи. При отщеплении от дезоксирибонуклеотидтрифосфата пирофосфата освобождается энергия, используемая на образование 3’, 5’-фосфодиэфирной связи между мономерами в процессе синтеза полимера.
 ДНК включает стадии инициации, элонгации и терминации. В ходе инициации образуются две")
Биосинтез (репликация) ДНК включает стадии инициации, элонгации и терминации. В ходе инициации образуются две репликативные вилки при участии ферментов ДНК-топоизомеразы, ДНК- хеликазы и белков, связывающихся с одноцепочечными участками ДНК (SSВ-белки).

ДНК-топоизомераза 1 присоединяется к участку ориджина, расщепляет одну из цепей ДНК и связывается с фосфатным остатком в точке разрыва, происходит локальное раскручивание двухцепочечной нити ДНК. Две молекулы ДНК-хеликазы, используя энергию АТФ, разрывают водородные связи между комплементарными основаниями и разделяют цепи ДНК. Одновременно ДНК-топоизомераза восстанавливает фосфодиэфирную связь и освобождается из связи с ДНК. SSВ-белки присоединяются к одноцепочечным участкам и препятствуют их повторному скручиванию в двойную спираль.


Элонгация репликации ДНК у эукариот На стадии элонгации образуются дочерние цепи ДНК на материнской ДНК. Этот процесс катализирует ДНК-полимераза. Все ферменты синтезируют полинуклеотидные цепи из дезоксирибонуклеозидтрифосфатаов только в направлении от 5’- к 3’- концу на антипараллельной матрице (от 3’- к 5’- концу).

1 - ДНК-полимераза синтезирует РНК –праймер, которым начинается лидирующая цепь, и каждый фрагмент Оказаки в отстающей нити ДНК. Лидирующая нить растет непрерывно, а отстающая – в виде фрагментов Оказаки, каждый их которых включает РНКпраймер (10 нуклеотидов) и участок ДНК, примерно равный длине ДНК в составе нуклеосомы (примерно 150 нуклеотидов) 2 – когда следующий фрагмент Оказаки достигает праймера предыдущего фрагмента, ДНК-полимераза отделяется от синтезированной цепи, а праймер предыдущего фрагмента удаляют эндонуклеаза и РНКаза, образуется брешь 3 – ДНК-полимераза удлиняет последний фрагмент Оказаки, заполняя брешь 4 – ДНК-лигаза сшивает предыдущий и вновь синтезированный фрагменты между собой

Новые цепи синтезируются неодинаково. Одна цепь на матрице ДНК с направлением от 3’- к 5’- концу растет непрерывно по ходу движения репликативной вилки и называется лидирующей. Вторая на матрице с направлением от 5’- к 3’- концу синтезируется против движения репликативной вилки в виде коротких фрагментов – фрагментов Оказаки, ее называют запаздывающей или отстающей.
 – олигорибонуклеотид (РНК),")
В участке расхождения цепей по принципу комплементарности сначала синтезируется праймер (затравка) – олигорибонуклеотид (РНК), состоящий из 10 нуклеотидов. Образование праймера катализирует праймаза, входящая в состав ДНКполимеразы α.

В отстающей нити каждый фрагмент Оказаки содержит около 200 нуклеотидов, включающих РНК-праймер и участок ДНК. Праймер удаляется эндонуклеазой и РНК-азой, а ДНК-полимераза β заполняет образующуюся брешь по принципу комплементарности, используя дезоксирибонуклеотидтрифосфаты в качестве субстратов.

ДНК-лигаза объединяет фрагменты в полинуклеотидную цепь, затрачивая молекулу АТФ на образование каждой 3’, 5’- фосфодиэфирной связи. Кофактором всех стадий репликации являются ионы Mg 2+. В результате образуются дочерние цепи, комплементарные и антипараллельные нитям материнской ДНК. После деления каждая дочерняя клетка получает диплоидный набор хромосом, идентичный материнской клетке.

Завершение синтеза ДНК в процессе репликации происходит на стадии терминации. Несмотря на высокую точность репликации, в молекуле ДНК постоянно • происходят повреждения, вызванные ультрафиолетовым облучением, радиационным излучением. урацил) • гидролитическое отщепление пуриновых оснований (аденина и гуанина) • образование пиримидиновых димеров между расположенными рядом в цепи основаниями • разрыв нуклеотидных цепей появление ковалентных сшивок между цепями или между цепями и гистонами нарушения комплементарности цепей

За сутки в каждой клетке происходят тысячи повреждений ДНК. Для их устранения в организме существуют системы репарации. Универсальная система репарации • специфическая эндонуклеаза обнаруживает нарушение комплементарности и гидролизует 3’, 5’-фосфодиэфирную связь в поврежденной нити ДНК • экзонуклеаза удаляет от 20 до 30 нуклеотидных остатков в области разрыва к 3’- концу образовавшейся бреши присоединяется ДНК-полимераза и, используя нуклеотидтрифосфаты в качестве субстратов и доноров энергии, заполняет брешь • ДНК – лигаза, используя АТФ как источник энергии, соединяет 3’, 5’-фосфодиэфирной связью место разрыва между вновь синтезированной и основной нитями ДНК.
 Транскрипция – это синтез РНК на матрице ДНК. Процесс катализируют РНК-полимеразы,")
БИОСИНТЕЗ РНК (ТРАНСКРИПЦИЯ) Транскрипция – это синтез РНК на матрице ДНК. Процесс катализируют РНК-полимеразы, которые подобно ДНК-полимеразам, образуют фосфодиэфирные связи между рибонуклеотидами в соответствии с принципами комплементарности к одной из нитей ДНК, которую обозначают как матричную.

У эукариот синтез РНК происходит в ядре и митохондриях практически постоянно вне зависимости от фаз клеточного цикла. В ядре РНК синтезируют 3 фермента: РНК-полимераза I катализирует образование р. РНК, РНК-полимераза II –синтез м. РНК, РНК-полимераза III – образование т. РНК. Нуклеотидтрифосфаты (АТФ, ГТФ, ЦТФ, УТФ) выполняют функции субстратов синтеза и источников энергии. В основе процесса лежит принцип комплементарного спаривания оснований.

В ходе транскрипции матрицей является нить ДНК, имеющая направление от 3’ к 5’-концу, так как все РНК-полимеразы осуществляют рост новых цепей РНК в направлении от 5’ к 3’-концу антипараллельно матрице.

Процесс транскрипции включает стадии • инициации • элонгации • терминации

РНК-полимеразы узнают место начала транскрипции - промотер, имеющий специфическую последовательность нуклеотидов – ТАТА-. • На стадии инициации к –ТАТАпоследовательности матричной цепи ДНК присоединяется белок –ТАТА-фактор, который стимулирует присоединение к ДНК РНКполимеразы и белковых факторов инициации транскрипции. Образующийся комплекс вызывает расплетение двойной нити ДНК длиной в один виток спирали (около 10 нуклеотидных пар).

На этапе элонгации происходит удаление факторов инициации и присоединение фактора элонгации. Синтез РНК осуществляется на матричной нити ДНК по принципу комплементарности. При этом в активном центре РНК-полимеразы каждый последующий нуклеотид связывается с 3’-концом предыдущего нуклеотида. По мере движения РНК-полимеразы по нити ДНК к освободившемуся промотору присоединяются новые молекулы фермента, поэтому один ген может одновременно транскрибироваться несколькими молекулами РНК-полимеразы.

Стадия терминации начинается, когда РНКполимераза достигает специфической последовательности нуклеотидов – сайта терминации. При этом фактор элонгации отделяется от РНКполимеразы, а фактор терминации присоединяется. Он облегчает отделение синтезированной молекулы пре-РНК и фермента от матрицы ДНК.

Молекулы РНК, которые синтезируются РНКполимеразами, функционально неактивны и являются молекулами-предшественниками - пре- РНК. Они превращаются в зрелые молекулы только после соответствующих посттранскрипционных модификаций – созревания молекул РНК.

Образование зрелых молекул м. РНК начинается еще в процессе синтеза молекулы РНК-полимеразой II на стадии элонгации. К 5’концу растущей нити РНК присоединяется 5’концом молекула ГТФ и отщепляется ортофосфат. Затем основание – гуанин в составе ГТФ – метилируется с образованием 7 -метил-ГТФ. Эту группу в составе м. РНК называют «кэп» (колпачок или шапочка). Кэп защищает 5’-конец м. РНК от действия нуклеаз и обеспечивает инициацию трансляции.
-полимераза последовательно удлиняет 3’-конец молекулы,")
После того как пре-м. РНК освобождается от РНКполимеразы II поли(А)-полимераза последовательно удлиняет 3’-конец молекулы, присоединяя от 150 до 200 остатков АМФ. Субстратом является АТФ. В результате на 3’конце пре-м. РНК образуется поли (А)- «хвост» , который также защищает м. РНК от расщепления РНКазами.

Установлено, что эукариотические ДНК состоят из участков, кодирующих последовательность аминокислот в отдельных доменах молекулы белка – экзонов и участков, не содержащих информацию о строении белка – интронов. В ходе транскрипции получаются пре-РНК, содержащие участки, комплеменарные экзонам и интронам. В процессе созревания м. РНК интроны удаляются, а экзоны соединяются между собой с высокой точностью при помощи комплексов из малых ядерных рибонуклеопротеинов (мя. РНП) – сплайсосом. Этот процесс получил название сплайсинга.

СОЗРЕВАНИЕ м. РНК Сплайсосомы гидролизуют 3’, 5’фосфодиэфирные связи на границе интрон-экзон и связывают экзоны между собой. Ферментативной активностью обладают РНК в составе мя. РНП.

Посттранскрипционные модификации т. РНК. В клетках синтезируется около 20 семейств т. РНК, молекулы которых содержат примерно 100 нуклеотидов. Представители каждого семейства способны связываться только с одной из 20 аминокислот, входящих в состав белков.

Посттранскрипционные модификации т. РНК. 1 -удаляются участки полинуклеотидной цепи на 5’-и 3’концах молекулы пре-т. РНК и интрон в центральной области молекулы 2 -модифицируются азотистые основания и к 3’концу присоединяется триплет ССА 3 -в цитоплазму выходят зрелые т. РНК

В ядре при формировании пространственной конформации т. РНК молекулы укорачиваются с 5’-и 3’концов с помощью специфических РНКаз и удаляется интрон. 10 -15% азотистых оснований в молекулах модифицируется к 3’концу всех т. РНК с помощью нуклеотидилтрансферазы последовательно один за другим присоединяется триплет нуклеотидов ССА, который необходим для связывания аминокислот, участвующих в синтезе белков

Посттранскрипционные модификации прер. РНК. Пре-р. РНК освобождается из комплекса с ДНК в виде крупного транскрипта 45 S. 1 -2% нуклеотидов этой молекулы метилируется по 2’-гидроксильной группе рибозы. Метильные группы служат маркерами для последующего расщепления пре-р. РНК на молекулы 18 S, 28 S и 5, 8 S. Самая короткая, 5 S р. РНК кодируется отдельным геном и включается в рибонуклеопротеиновые частицы, содержащие 28 S и 5, 8 S РНК, образуя большую субъединицу рибосомы. 18 S РНК формирует малую 40 S субъединицу рибосомы.

1 -транскрипция пре-р. РНК 2 -связывание 45 S р. РНК с белками и 5 S р. РНК 3 -метилирование пре-р. РНК и расщепление на отдельные фрагменты 4 -дальнейшее укорочение р. РНК и формирование 40 S и 60 S субъединиц рибосом Субъединицы рибосомы и все зрелые м. РНК и т. РНК поступают в цитоплазму клетки и используются в синтезе белков.
Презентация (нуклеиновые кислоты).ppt