Nukleinovye_Kisloty_33_33_33 (1).ppt
- Количество слайдов: 64
 Нуклеиновые Кислоты
Нуклеиновые Кислоты
 Нуклеиновые кислоты –") Нуклеиновые кислоты l l Открыты швейцарским биохимиком Фридрихом Мишером (1844 -1895) Нуклеиновые кислоты – высокомолекулярные органические соединения, обеспечивающие хранение и передачу наследственной ( генетической информации) в живых организмах.
Нуклеиновые кислоты l l Открыты швейцарским биохимиком Фридрихом Мишером (1844 -1895) Нуклеиновые кислоты – высокомолекулярные органические соединения, обеспечивающие хранение и передачу наследственной ( генетической информации) в живых организмах.
 СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ Химическая структура нуклеиновых кислот. Первичная структура. Под первичной структурой подразумевается последовательность расположения мономерных звеньев в линейных полимерах. Для нуклеиновых кислот, т. е. ДНК и РНК, такими мономерами являются нуклеотиды.
СТРУКТУРА НУКЛЕИНОВЫХ КИСЛОТ Химическая структура нуклеиновых кислот. Первичная структура. Под первичной структурой подразумевается последовательность расположения мономерных звеньев в линейных полимерах. Для нуклеиновых кислот, т. е. ДНК и РНК, такими мономерами являются нуклеотиды.
 Нуклеотиды состоят из остатков гетероциклического основания, пентозы и фосфата. Исходными основаниями, производные которых входят в природные нуклеотиды, являются пурин и пиримидин В состав природной ДНК входят 4 азотистых основания; 2 из них, аденин и гуанин, являются производными пурина, а 2 других цитозин и тимин – пиримидина.
Нуклеотиды состоят из остатков гетероциклического основания, пентозы и фосфата. Исходными основаниями, производные которых входят в природные нуклеотиды, являются пурин и пиримидин В состав природной ДНК входят 4 азотистых основания; 2 из них, аденин и гуанин, являются производными пурина, а 2 других цитозин и тимин – пиримидина.
") Структуры соединений Пурин Пиримидин Структура пурина и пиримидина (азотистые основания)
Структуры соединений Пурин Пиримидин Структура пурина и пиримидина (азотистые основания)
 Тимин Цитозин Структура азотистых оснований, входящих в состав ДНК.
Тимин Цитозин Структура азотистых оснований, входящих в состав ДНК.
 Общая структура нуклеотида
Общая структура нуклеотида
 50 е годы XX - века А. Тодд установил точную структуру связей, соединяющих нуклеотиды одной цепи – фосфодиэфирные связи -
50 е годы XX - века А. Тодд установил точную структуру связей, соединяющих нуклеотиды одной цепи – фосфодиэфирные связи -
 Структура 3' , 5' фосфодиэфирных связей в ДНК.
Структура 3' , 5' фосфодиэфирных связей в ДНК.
 История открытия ДНК l l l 1950 г. М. Уилкинс получил рентгенограмму кристаллических волокон ДНК Розалин Франклин на менее упорядоченных агрегатах, которые образуются при более повышенной влажности Увидела четкий крестообразный рисунок – опознавательный знак двойной спирали
История открытия ДНК l l l 1950 г. М. Уилкинс получил рентгенограмму кристаллических волокон ДНК Розалин Франклин на менее упорядоченных агрегатах, которые образуются при более повышенной влажности Увидела четкий крестообразный рисунок – опознавательный знак двойной спирали
.") Нуклеотидный состав l l Впервые количественно проанализировал нуклеотидный состав американский биохимик Эдвин Чаргафф (1951). Правило Чаргаффа – число пуриновых оснований в ДНК всегда равно числу пиримидиновых, количество А равно количеству Т, с Г – количеству Ц
Нуклеотидный состав l l Впервые количественно проанализировал нуклеотидный состав американский биохимик Эдвин Чаргафф (1951). Правило Чаргаффа – число пуриновых оснований в ДНК всегда равно числу пиримидиновых, количество А равно количеству Т, с Г – количеству Ц
 Строение молекулы 1953 г Американский биохимик Дж. Уотсон и английский физик Ф. Крик представили модель строения молекулы ДНК
Строение молекулы 1953 г Американский биохимик Дж. Уотсон и английский физик Ф. Крик представили модель строения молекулы ДНК
 Структура ДНК l Сахаро – фосфатный остов находится на периферии молекулы ДНК. Пуриновые и пиримидиновые основания в центре. Между ними образуются водородные связи
Структура ДНК l Сахаро – фосфатный остов находится на периферии молекулы ДНК. Пуриновые и пиримидиновые основания в центре. Между ними образуются водородные связи
 Структура ДНК l l Пурин в одной цепи связан с пиримидином в другой цепи Аденин может спариваться только с Тимином, а гуанин с цитозином
Структура ДНК l l Пурин в одной цепи связан с пиримидином в другой цепи Аденин может спариваться только с Тимином, а гуанин с цитозином
 Структура ДНК l l А Т (две водородные связи Г Ц (три водородные связи
Структура ДНК l l А Т (две водородные связи Г Ц (три водородные связи
 • Каждая из пар оснований обладает симметрией, позволяющей ей включаться в двойную спираль в двух ориентациях А=Т и Т=А. • Т. о, в каждой из цепей ДНК основания могут чередоваться всеми существующими способами • Противолежащие последовательности и соответствующие полинуклеотидные цепи называют комплиментарными
• Каждая из пар оснований обладает симметрией, позволяющей ей включаться в двойную спираль в двух ориентациях А=Т и Т=А. • Т. о, в каждой из цепей ДНК основания могут чередоваться всеми существующими способами • Противолежащие последовательности и соответствующие полинуклеотидные цепи называют комплиментарными
 остатки в молекулах нуклеотидов присутствуют в циклической (фуранозной) (3 О форме; 2") Сахарные (пентозные) остатки в молекулах нуклеотидов присутствуют в циклической (фуранозной) (3 О форме; 2 дезоксирибоза в ДНК и рибоза в РНК В состав РНК входят основания аденин, гуанин, цитозин и урацил и в качестве углеводного (пентозного) компонента — рибоза (1 р В ри бофураноза) Полная структура нуклеотида (мономера нуклеиновых кислот) включает нуклеозидную группу (т. е. гетероциклическое основание и пентозу, в данном случае — рибозу) и остаток фосфата. Образование полинуклеотидной цепи, как ДНК, так и РНК, происходит за счет 3' , 5' фосфодиэфирных связей
Сахарные (пентозные) остатки в молекулах нуклеотидов присутствуют в циклической (фуранозной) (3 О форме; 2 дезоксирибоза в ДНК и рибоза в РНК В состав РНК входят основания аденин, гуанин, цитозин и урацил и в качестве углеводного (пентозного) компонента — рибоза (1 р В ри бофураноза) Полная структура нуклеотида (мономера нуклеиновых кислот) включает нуклеозидную группу (т. е. гетероциклическое основание и пентозу, в данном случае — рибозу) и остаток фосфата. Образование полинуклеотидной цепи, как ДНК, так и РНК, происходит за счет 3' , 5' фосфодиэфирных связей
. Урацил Рибоза Структура компонентов РНК — урацила") Структура пентозного компонента ДНК — дезоксирибозы (дезоксирибофуранозы). Урацил Рибоза Структура компонентов РНК — урацила и рибозы
Структура пентозного компонента ДНК — дезоксирибозы (дезоксирибофуранозы). Урацил Рибоза Структура компонентов РНК — урацила и рибозы
 Вторичная структура нуклеиновых кислот. Типы вторичных структур Основной тип вторичной структуры нуклеиновых кислот — двойная спираль. Формирование этой структуры определяется межнуклеотидными водородными связями, которые образуются между основаниями, входящими в так называемые канонические пары А-Т(U) и G-С. Каноническим парам (и только им) свойственно точное равенство геометрических параметров, необходимых для существования двойной спирали ДНК. Помимо водородных связей, структура двойной спирали дополнительно стабилизирована стэкингвзаимодействиями, электростатическими и гидрофобными связями.
Вторичная структура нуклеиновых кислот. Типы вторичных структур Основной тип вторичной структуры нуклеиновых кислот — двойная спираль. Формирование этой структуры определяется межнуклеотидными водородными связями, которые образуются между основаниями, входящими в так называемые канонические пары А-Т(U) и G-С. Каноническим парам (и только им) свойственно точное равенство геометрических параметров, необходимых для существования двойной спирали ДНК. Помимо водородных связей, структура двойной спирали дополнительно стабилизирована стэкингвзаимодействиями, электростатическими и гидрофобными связями.
 Третичная структура. Под третичной структурой подразумевается общая форма молекул биополимеров. На этом уровне структура как белков, так и нуклеиновых кислот не имеет определенных типов с жестко заданными параметрами. Так, для ДНК в качестве варианта третичной можно рассматривать сверхспиральную структуру, когда замкнутая двойная спираль образует не кольцо, а структуру с витками более высокого порядка. Такие структуры характеризуются (в качестве главного параметра) количеством супервитков; как правило, большинство природных ДНК сверхспирализованы.
Третичная структура. Под третичной структурой подразумевается общая форма молекул биополимеров. На этом уровне структура как белков, так и нуклеиновых кислот не имеет определенных типов с жестко заданными параметрами. Так, для ДНК в качестве варианта третичной можно рассматривать сверхспиральную структуру, когда замкнутая двойная спираль образует не кольцо, а структуру с витками более высокого порядка. Такие структуры характеризуются (в качестве главного параметра) количеством супервитков; как правило, большинство природных ДНК сверхспирализованы.
 Типы двойных спиралей ДНК Форма А В С 2 правая левая Число пар оснований на одном витке спирали 10, 7 10, 0 9, 3 12 Угол между соседними парами оснований (градусы) +33, 6 +36, 0 +38, 6 -30 Расстояние между соседними парами оснований (А) 2, 3 3, 4 3, 0 3, 8 Угол наклона оснований к перпендикуляру к оси спирали (градусы) + 19 -1, 2 -6 -9 23 20 19 18 Спираль Диаметр спирали (А)
Типы двойных спиралей ДНК Форма А В С 2 правая левая Число пар оснований на одном витке спирали 10, 7 10, 0 9, 3 12 Угол между соседними парами оснований (градусы) +33, 6 +36, 0 +38, 6 -30 Расстояние между соседними парами оснований (А) 2, 3 3, 4 3, 0 3, 8 Угол наклона оснований к перпендикуляру к оси спирали (градусы) + 19 -1, 2 -6 -9 23 20 19 18 Спираль Диаметр спирали (А)
 Четвертичная структура — укладка молекул в полимолекулярные ансамбли. Для нуклеиновых кислот это ансамбли, включающие также молекулы белков.
Четвертичная структура — укладка молекул в полимолекулярные ансамбли. Для нуклеиновых кислот это ансамбли, включающие также молекулы белков.
 Рибонуклеиновая кислота РНК l l l Биополимер, мономером которого является нуклеотид. РНК представляет собой однонитевую молекулу Существует 4 типа нуклеотидов. Они состоят из азотистого основания, сахара пентозы, и фосфорной кислоты. Три азотистых основания такие же как в днк – А, Г, Ц. Т заменен на урацил -У
Рибонуклеиновая кислота РНК l l l Биополимер, мономером которого является нуклеотид. РНК представляет собой однонитевую молекулу Существует 4 типа нуклеотидов. Они состоят из азотистого основания, сахара пентозы, и фосфорной кислоты. Три азотистых основания такие же как в днк – А, Г, Ц. Т заменен на урацил -У
 АТФ –аденозин трифосфорная кислота Н 3 РО 4 аденин рибоза Н 3 РО 4 Реакция отщепления каждой молекулы фосфорной кислоты сопровождается освобождением 419 к. Дж/ моль
АТФ –аденозин трифосфорная кислота Н 3 РО 4 аденин рибоза Н 3 РО 4 Реакция отщепления каждой молекулы фосфорной кислоты сопровождается освобождением 419 к. Дж/ моль
 В состав РНК входят основания аденин, гуанин, цитозин и урацил и в качестве углеводного (пентозного) компонента — рибоза (1 р В ри бофураноза) Полная структура нуклеотида (мономера нуклеиновых кислот) включает нуклеозидную группу (т. е. гетероциклическое основание и пентозу, в данном случае — рибозу) и остаток фосфата. Образование полинуклеотидной цепи, как ДНК, так и РНК, происходит за счет 3' , 5' фосфодиэфирных связей
В состав РНК входят основания аденин, гуанин, цитозин и урацил и в качестве углеводного (пентозного) компонента — рибоза (1 р В ри бофураноза) Полная структура нуклеотида (мономера нуклеиновых кислот) включает нуклеозидную группу (т. е. гетероциклическое основание и пентозу, в данном случае — рибозу) и остаток фосфата. Образование полинуклеотидной цепи, как ДНК, так и РНК, происходит за счет 3' , 5' фосфодиэфирных связей
 БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ Биосинтез ДНК про- и эукариот. Принцип полуконсервативной репликации ДНК Биосинтез ДНК осуществляется специальными ферментами, так называемыми ДНК-полимеразами; в его основе лежит принцип копирования исходной цепи ДНК (путем образования 1. комплементарных пар нуклеотидов) с образованием двух идентичных дочерних молекул. Процесс биосинтеза ДНК имеет ряд принципиальных особенностей.
БИОСИНТЕЗ НУКЛЕИНОВЫХ КИСЛОТ Биосинтез ДНК про- и эукариот. Принцип полуконсервативной репликации ДНК Биосинтез ДНК осуществляется специальными ферментами, так называемыми ДНК-полимеразами; в его основе лежит принцип копирования исходной цепи ДНК (путем образования 1. комплементарных пар нуклеотидов) с образованием двух идентичных дочерних молекул. Процесс биосинтеза ДНК имеет ряд принципиальных особенностей.
 Поскольку синтез ДНК идет по 2. полуконсервативному механизму, т. е. новая двойная спираль содержит одну полинуклеотидную нить новую и одну старую, для начала процесса необходима эта старая нить, т. е. ДНК матрица; новая нить синтезируется как комплементарная нити матричной. 3. Антипараллельность 4. Прерывистость
Поскольку синтез ДНК идет по 2. полуконсервативному механизму, т. е. новая двойная спираль содержит одну полинуклеотидную нить новую и одну старую, для начала процесса необходима эта старая нить, т. е. ДНК матрица; новая нить синтезируется как комплементарная нити матричной. 3. Антипараллельность 4. Прерывистость
 Участок между двумя точками, в которых начинается синтез дочерних цепей – репликон. Это – единица репликации. В каждой молекуле ДНК имеется много репликонов – прерывистость процесса репликации. Ферменты: 1. ДНК – полимераза 2. Геликаза 3. Топоизомеразы 4. Лигаза 5. С каждой из формирующихся нитей связываются SSB – белки стабилизирующие одноцепочечное состояние
Участок между двумя точками, в которых начинается синтез дочерних цепей – репликон. Это – единица репликации. В каждой молекуле ДНК имеется много репликонов – прерывистость процесса репликации. Ферменты: 1. ДНК – полимераза 2. Геликаза 3. Топоизомеразы 4. Лигаза 5. С каждой из формирующихся нитей связываются SSB – белки стабилизирующие одноцепочечное состояние
 При репликации ДНК ферменты, осуществляющие этот процесс, должны выполнить: снятие топологических напряжений ДНК; расплетание ДНК; синтез РНК затравки; синтез дочерней цепи ДНК; вырезание последнего встроенного нуклеотида, если он некомплементарен матрице, т. е. встроен ошибочно (З' экзонуклеазная активность); вырезание РНК затравки (5' экзонуклеазная активность) и застройку брешей новой ДНК; сшивку фрагментов новой ДНК в одну ковалентно непрерывную цепь.
При репликации ДНК ферменты, осуществляющие этот процесс, должны выполнить: снятие топологических напряжений ДНК; расплетание ДНК; синтез РНК затравки; синтез дочерней цепи ДНК; вырезание последнего встроенного нуклеотида, если он некомплементарен матрице, т. е. встроен ошибочно (З' экзонуклеазная активность); вырезание РНК затравки (5' экзонуклеазная активность) и застройку брешей новой ДНК; сшивку фрагментов новой ДНК в одну ковалентно непрерывную цепь.
 новой полинуклеотидной цепи") Комплекс матрица-затравка Механизм действия ДНК полимераз таков, что синтез (на матрице) новой полинуклеотидной цепи не может начаться с включения в нее первого нуклеотида. Синтез идет только как удлинение уже существующего полинуклеотида, который комплементарен матрице и образует с ней двуспиральный комплекс матрицазатравка. Каждый нуклеотид включается в цепь лишь в случае его комплементарности нуклеотиду, занимающему данную позицию в составе матрицы. Природной затравкой для репликации ДНК является олигонуклеотид РНК -затравка.
Комплекс матрица-затравка Механизм действия ДНК полимераз таков, что синтез (на матрице) новой полинуклеотидной цепи не может начаться с включения в нее первого нуклеотида. Синтез идет только как удлинение уже существующего полинуклеотида, который комплементарен матрице и образует с ней двуспиральный комплекс матрицазатравка. Каждый нуклеотид включается в цепь лишь в случае его комплементарности нуклеотиду, занимающему данную позицию в составе матрицы. Природной затравкой для репликации ДНК является олигонуклеотид РНК -затравка.
 Запаздывающая цепь – короткие") Лидирующая цепь – длинный непрерывный участок (1600000 н. п. ) Запаздывающая цепь – короткие участки (1500 н. п) фрагменты Оказаки
Лидирующая цепь – длинный непрерывный участок (1600000 н. п. ) Запаздывающая цепь – короткие участки (1500 н. п) фрагменты Оказаки
 Старая ДНК Две двойные цепи новой ДНК Репликативная вилка. Короткие стрелки — фрагменты Оказаки
Старая ДНК Две двойные цепи новой ДНК Репликативная вилка. Короткие стрелки — фрагменты Оказаки
 Варианты роста новой цепи ДНК: рост происходит в одном направлении от точки начала репликации; рост происходит в обоих направлениях; рост происходит в одном направлении, вдоль одноцепочечной ДНК
Варианты роста новой цепи ДНК: рост происходит в одном направлении от точки начала репликации; рост происходит в обоих направлениях; рост происходит в одном направлении, вдоль одноцепочечной ДНК
 В пределах одной молекулы ДНК может быть одна или несколько точек начала репликации и соответствующих им репликонов Организм Число репликонов Длина, тыс. пар Скорость репликации, нукл. /мин Бактерии 1 4200 50000 Дрожжи 500 40 3600 Растения 35000 Дрозофила 3500 40 2600 Мышь 25000 150 2200 300
В пределах одной молекулы ДНК может быть одна или несколько точек начала репликации и соответствующих им репликонов Организм Число репликонов Длина, тыс. пар Скорость репликации, нукл. /мин Бактерии 1 4200 50000 Дрожжи 500 40 3600 Растения 35000 Дрозофила 3500 40 2600 Мышь 25000 150 2200 300
 Химический механизм присоединения мономерных звеньев к ДНК, направление синтеза ДНК, репликативная вилка, фрагменты Оказаки. Суммарное уравнение ДНК полимеразной реакции: (д. НМФ)n + д НТФ = (д. НМФ)пn+ 1 + ФФ. Таким образом, низкомолекулярным субстратом реакции являются нуклеозидтрифосфаты, а продуктом — неорганический пирофосфат Пирофосфат быстро расщепляется внутриклеточной пирофосфатазой с образованием ортофосфата: ФФ → 2 Ф.
Химический механизм присоединения мономерных звеньев к ДНК, направление синтеза ДНК, репликативная вилка, фрагменты Оказаки. Суммарное уравнение ДНК полимеразной реакции: (д. НМФ)n + д НТФ = (д. НМФ)пn+ 1 + ФФ. Таким образом, низкомолекулярным субстратом реакции являются нуклеозидтрифосфаты, а продуктом — неорганический пирофосфат Пирофосфат быстро расщепляется внутриклеточной пирофосфатазой с образованием ортофосфата: ФФ → 2 Ф.
 ДНК полимеразная реакция
ДНК полимеразная реакция
 ДНК-полимераза имеет два субстрата — комплекс матрица затравка и дезоксирибонуклеозид 5' трифосфат. Реакция осуществляется с участием концевой З' гидроксильной группы, атакующей а фосфатную группу дезоксинуклеозидтрифосфата. Таким образом, удлинение цепи ДНК (и РНК) идет только с З'-конца, матричная и дочерняя цепи расположены антипараллельно. Фермент имеет участки связывания для обоих субстратов, которые вступают в реакцию в виде комплексов с ионами магния, что обеспечивает их правильную взаимную ориентацию
ДНК-полимераза имеет два субстрата — комплекс матрица затравка и дезоксирибонуклеозид 5' трифосфат. Реакция осуществляется с участием концевой З' гидроксильной группы, атакующей а фосфатную группу дезоксинуклеозидтрифосфата. Таким образом, удлинение цепи ДНК (и РНК) идет только с З'-конца, матричная и дочерняя цепи расположены антипараллельно. Фермент имеет участки связывания для обоих субстратов, которые вступают в реакцию в виде комплексов с ионами магния, что обеспечивает их правильную взаимную ориентацию
 Важная особенность ДНК полимеразы — несущественность конкретной нуклеотидной последовательности матрицы, которая реплицируется с образованием точной копии независимо от нуклеотидного «текста» . При репликации природной двунитевой ДНК, состоящей из двух антипараллельных цепей, образуется так называемая репликативная вилка
Важная особенность ДНК полимеразы — несущественность конкретной нуклеотидной последовательности матрицы, которая реплицируется с образованием точной копии независимо от нуклеотидного «текста» . При репликации природной двунитевой ДНК, состоящей из двух антипараллельных цепей, образуется так называемая репликативная вилка
 ДНК-полимеразы эукариот ДНК-полимераза α Основной фермент ядерной репликации. Состоит из нескольких субъединиц разного размера (у дрозофилы 148, 58, 46 и 42 к. Д). Полимеразной активностью обладает субъединица 148 к. Д, одна из субъединиц является праймазой. Повидимому, в основном синтезирует отстающую цепь (т. е. фрагменты Оказаки).
ДНК-полимеразы эукариот ДНК-полимераза α Основной фермент ядерной репликации. Состоит из нескольких субъединиц разного размера (у дрозофилы 148, 58, 46 и 42 к. Д). Полимеразной активностью обладает субъединица 148 к. Д, одна из субъединиц является праймазой. Повидимому, в основном синтезирует отстающую цепь (т. е. фрагменты Оказаки).
 ДНК-полимераза ƅ Состоит из одного полипептида около 40 к. Д, по видимому, является ферментом исправление ошибок репликации. репарации- ДНК-полимераза у Состоит из нескольких идентичных субъединиц около 50 к. Д, фермент синтеза ДНК митохондрий. ДНК-полимераза 8 Состоит из двух неидентичных субъединиц, общая молекулярная масса около 150 к. Д; обладает З' экзонуклеазной активностью. Функция окончательно не выяснена, по видимому, синтезирует ведущую цепь ДНК.
ДНК-полимераза ƅ Состоит из одного полипептида около 40 к. Д, по видимому, является ферментом исправление ошибок репликации. репарации- ДНК-полимераза у Состоит из нескольких идентичных субъединиц около 50 к. Д, фермент синтеза ДНК митохондрий. ДНК-полимераза 8 Состоит из двух неидентичных субъединиц, общая молекулярная масса около 150 к. Д; обладает З' экзонуклеазной активностью. Функция окончательно не выяснена, по видимому, синтезирует ведущую цепь ДНК.
 Обратная транскриптаза — РНК-зависимая ДНКполимераза Фермент вирусного или клеточного происхождения, синтезирующий ДНК на матрице РНК.
Обратная транскриптаза — РНК-зависимая ДНКполимераза Фермент вирусного или клеточного происхождения, синтезирующий ДНК на матрице РНК.
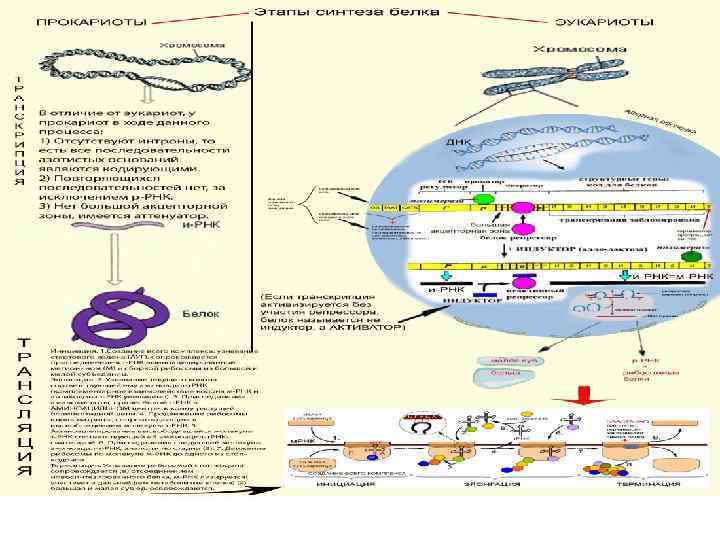
 ОРГАНИЗАЦИЯ ГЕНОВ ПРОКАРИОТ И РЕГУЛЯЦИЯ ИХ АКТИВНОСТИ Транскрипция проходит 4 стадии связывания ДНК матрицы, инициации, элонгации и терминации; в рамках каждой из этих стадий осуществляются определенные функции мультимолекулярного фермент ного комплекса — РНК полимеразы
ОРГАНИЗАЦИЯ ГЕНОВ ПРОКАРИОТ И РЕГУЛЯЦИЯ ИХ АКТИВНОСТИ Транскрипция проходит 4 стадии связывания ДНК матрицы, инициации, элонгации и терминации; в рамках каждой из этих стадий осуществляются определенные функции мультимолекулярного фермент ного комплекса — РНК полимеразы
 Промотор — последовательность ДНК, необходимая для образова ния комплекса РНК полимеразы с ДНК матрицей; расположена вблизи нуклеотида, с которого начинается синтез РНК. Стартовая точка транскрипции — нуклеотид, с которого начинается синтез РНК. Терминатор — последовательность ДНК, узнаваемая РНК полимеразой и являющаяся сигналом окончания транскрипции Матричной для каждой молекулы РНК является одна из двух нитей ДНК; в областях перекрывания участков синтеза различных молекул РНК (транскриптонов) матричными могут быть обе цепи
Промотор — последовательность ДНК, необходимая для образова ния комплекса РНК полимеразы с ДНК матрицей; расположена вблизи нуклеотида, с которого начинается синтез РНК. Стартовая точка транскрипции — нуклеотид, с которого начинается синтез РНК. Терминатор — последовательность ДНК, узнаваемая РНК полимеразой и являющаяся сигналом окончания транскрипции Матричной для каждой молекулы РНК является одна из двух нитей ДНК; в областях перекрывания участков синтеза различных молекул РНК (транскриптонов) матричными могут быть обе цепи
 Собственно РНК полимеразная активность, т. е. образование фосфодиэфирных связей с нуклеотидами, комплементарными матрице, свойственна так называемому минимальному ферменту; таким образом, холофермент РНК полимеразы может быть разделен на минимальный фермент α 2ββ’ и σ фактор. Инициацию синтеза РНК осуществляет только холофермент, затем σ фактор диссоциирует и минимальный фермент ведет элонгацию. Основная функция σ фактора — узнавание промоторов и обеспечение прочного связывания РНК полимеразы с промоторами. Существуют минорные разновидности σ фактора, узнающие определенные группы промоторов.
Собственно РНК полимеразная активность, т. е. образование фосфодиэфирных связей с нуклеотидами, комплементарными матрице, свойственна так называемому минимальному ферменту; таким образом, холофермент РНК полимеразы может быть разделен на минимальный фермент α 2ββ’ и σ фактор. Инициацию синтеза РНК осуществляет только холофермент, затем σ фактор диссоциирует и минимальный фермент ведет элонгацию. Основная функция σ фактора — узнавание промоторов и обеспечение прочного связывания РНК полимеразы с промоторами. Существуют минорные разновидности σ фактора, узнающие определенные группы промоторов.
 Субъединицы холофермента РНК-полимеразы Е. сoli Число Наимено Молекулярная субъедини вание масса, цилодальтон ц 2 α 40 1 β 155 1 β’ 160 1 σ 85
Субъединицы холофермента РНК-полимеразы Е. сoli Число Наимено Молекулярная субъедини вание масса, цилодальтон ц 2 α 40 1 β 155 1 β’ 160 1 σ 85
 Стадии цикла σ-фактора Образование холофермента, т. е. ассоциация минимального ком плекса и σ фактора. Связывание холофермента с промотором, образование закрыто го комплекса (при связывании с неспецифической последовательно стью процесс дальше не идет). Переход к открытому двойному комплексу, т. е. расплавление уча стка связанной ДНК (около 15 пар нуклеотидов из примерно 60, экранируемых ферментом), возникновение прочного комплекса холо фермент — ДНК (t 1/2 =0, 2 с, превращение комплекса идет по пути не диссоциации, а перехода в тройной комплекс). Образование прочно го комплекса обеспечивает строгую процессивность синтеза РНК. Образование тройного комплекса, т. е. синтез первого динуклео тидного звена дочерней РНК. Диссоциация σ фактора.
Стадии цикла σ-фактора Образование холофермента, т. е. ассоциация минимального ком плекса и σ фактора. Связывание холофермента с промотором, образование закрыто го комплекса (при связывании с неспецифической последовательно стью процесс дальше не идет). Переход к открытому двойному комплексу, т. е. расплавление уча стка связанной ДНК (около 15 пар нуклеотидов из примерно 60, экранируемых ферментом), возникновение прочного комплекса холо фермент — ДНК (t 1/2 =0, 2 с, превращение комплекса идет по пути не диссоциации, а перехода в тройной комплекс). Образование прочно го комплекса обеспечивает строгую процессивность синтеза РНК. Образование тройного комплекса, т. е. синтез первого динуклео тидного звена дочерней РНК. Диссоциация σ фактора.
 Нуклеотидные последовательности промоторов прокариот. 60 5' 35 ТТСАСА 10 ТАТААТ +1 САТ Такой набор последовательностей достаточен для связывания РНК полимеразы и начала транскрипции. Гены, транскрипция которых постоянна, т. е. не требует специальных регуляторных воздействий, называются конститутивными; гены, требующие таких воздействий (индукции) для начала транскрипции, называются индуцибельными. Как позитивная (усиление), так и негативная (подавление) регуляция активности транскрипции осуществляется с помощью особых белков регуляторов.
Нуклеотидные последовательности промоторов прокариот. 60 5' 35 ТТСАСА 10 ТАТААТ +1 САТ Такой набор последовательностей достаточен для связывания РНК полимеразы и начала транскрипции. Гены, транскрипция которых постоянна, т. е. не требует специальных регуляторных воздействий, называются конститутивными; гены, требующие таких воздействий (индукции) для начала транскрипции, называются индуцибельными. Как позитивная (усиление), так и негативная (подавление) регуляция активности транскрипции осуществляется с помощью особых белков регуляторов.
 Механизмы репрессии и активации транскрипции Белки, подавляющие транскрипцию, называются репрессорами. Если белок усиливает транскрипцию путем уменьшения активности репрессора, он называется индуктором; если транскрипция активируется непосредственно, без участия репрессора, белок называется активатором. Участки связывания репрессора или активатора на ДНК называются операторами, а единица транскрипции, находящаяся под контролем данного оператора, — опероном (у прокариот это, как правило, несколько генов, регулируемых параллельно).
Механизмы репрессии и активации транскрипции Белки, подавляющие транскрипцию, называются репрессорами. Если белок усиливает транскрипцию путем уменьшения активности репрессора, он называется индуктором; если транскрипция активируется непосредственно, без участия репрессора, белок называется активатором. Участки связывания репрессора или активатора на ДНК называются операторами, а единица транскрипции, находящаяся под контролем данного оператора, — опероном (у прокариот это, как правило, несколько генов, регулируемых параллельно).
 Механизмы репрессии и активации транскрипции Простейший механизм репрессии — связывание репрессора с промоторной областью, т. е. перекрывание промотора и оператора. В этом случае инициации транскрипции не происходит, поскольку участок связывания РНК полимеразы уже занят репрессором. Простейший механизм активации — образование комплекса активатора и РНК полимеразы, что облегчает формирование открытого комплекса на матрице ДНК.
Механизмы репрессии и активации транскрипции Простейший механизм репрессии — связывание репрессора с промоторной областью, т. е. перекрывание промотора и оператора. В этом случае инициации транскрипции не происходит, поскольку участок связывания РНК полимеразы уже занят репрессором. Простейший механизм активации — образование комплекса активатора и РНК полимеразы, что облегчает формирование открытого комплекса на матрице ДНК.
 ОРГАНИЗАЦИЯ ГЕНОВ ЭУКАРИОТ И РЕГУЛЯЦИЯ ИХ АКТИВНОСТИ Регуляция синтеза РНК-полимеразой II. Универсальные регуляторные последовательности Значимость таких регуляторных последовательностей заключается, во первых, в контроле правильного начала транскрипции и, во вторых, в передаче сигнала, регулирующего транскрипцию, к РНК синтезирующему аппарату. Реализация обеих этих функций осуществляется через связывание с регуляторными последовательностями специфических белков, которые взаимодействуют с РНК полимеразой, связывают другие белки эффекторы и др.
ОРГАНИЗАЦИЯ ГЕНОВ ЭУКАРИОТ И РЕГУЛЯЦИЯ ИХ АКТИВНОСТИ Регуляция синтеза РНК-полимеразой II. Универсальные регуляторные последовательности Значимость таких регуляторных последовательностей заключается, во первых, в контроле правильного начала транскрипции и, во вторых, в передаче сигнала, регулирующего транскрипцию, к РНК синтезирующему аппарату. Реализация обеих этих функций осуществляется через связывание с регуляторными последовательностями специфических белков, которые взаимодействуют с РНК полимеразой, связывают другие белки эффекторы и др.
, расположенные в позиции 30 / 20 относительно точки инициации синтеза РНК.") ТАТА-боксы последовательности ТАТАА(А), расположенные в позиции 30 / 20 относительно точки инициации синтеза РНК. Они обнаружены в большинстве эукариотических генов; считается, что ТАТА боксы определяют, что потенциально активная точка начала синтеза РНК оказывается реальным стартом транскрипции. СААТ-боксы — последовательности С(О)СААТ, находящиеся в положении 50 / 150. В отсутствие ТАТА бокса могут «активировать» потенциальный старт транскрипции
ТАТА-боксы последовательности ТАТАА(А), расположенные в позиции 30 / 20 относительно точки инициации синтеза РНК. Они обнаружены в большинстве эукариотических генов; считается, что ТАТА боксы определяют, что потенциально активная точка начала синтеза РНК оказывается реальным стартом транскрипции. СААТ-боксы — последовательности С(О)СААТ, находящиеся в положении 50 / 150. В отсутствие ТАТА бокса могут «активировать» потенциальный старт транскрипции
 GС-мотивы — участки длиной 5— 15 нуклеотидных пар, состоящие из О и С; не имеют строгого консенсуса, но обычно содержат последовательность (О)ООСОО(О). В отсутствие и ТАТА , и САА'Г боксов ОС последовательность определяет правильную инициацию транскрипции. Примечания а) Последовательности, непосредственно прилегающие к точкам инициации синтеза РНК, не объединены строгим консенсусом. б) Как правило, каждая промоторная область содержит и ТАТА бокс, и СААТ или ОС, но каждая из приведенных последовательностей может находиться в промоторе и без двух других и самостоятельно регулировать транскрипцию.
GС-мотивы — участки длиной 5— 15 нуклеотидных пар, состоящие из О и С; не имеют строгого консенсуса, но обычно содержат последовательность (О)ООСОО(О). В отсутствие и ТАТА , и САА'Г боксов ОС последовательность определяет правильную инициацию транскрипции. Примечания а) Последовательности, непосредственно прилегающие к точкам инициации синтеза РНК, не объединены строгим консенсусом. б) Как правило, каждая промоторная область содержит и ТАТА бокс, и СААТ или ОС, но каждая из приведенных последовательностей может находиться в промоторе и без двух других и самостоятельно регулировать транскрипцию.
 С перечисленными выше универсальными последовательностями взаимодействуют определенные белковые факторы, которые в силу универсальности последовательностей являются общими регуляторами транскрипции для большинства эукариотических генов
С перечисленными выше универсальными последовательностями взаимодействуют определенные белковые факторы, которые в силу универсальности последовательностей являются общими регуляторами транскрипции для большинства эукариотических генов
 ТАТА-боксы. С ними взаимодействует сложный белковый комплекс ТР 1 Ш, состоящий из 15— 20 субъединиц; непосредственно узнает данную последовательность ТВР, субъединица с молекулярной массой около 100— 120 к. Д. Компоненты фактора ТР 1 Ш образуют комплексы с многочисленными белками регуляторами и модулируют активность РНК полимеразы II. Действие минимального ТАТА связывающего комплекса осуществляется по схеме: ТВР связывается с ТАТА боксом; с образовавшимся комплексом связывается фактор ТРПВ; к ТРИ В в составе комплекса присоединяются фактор ТРПР и РНК полимераза II; образовавшийся минимальный комплекс присоединяет факторы ТРИ Е/Н, т. е. образуется полный инициаторный комплекс, который способен осуществить АТФ зависимую инициацию синтеза РНК.
ТАТА-боксы. С ними взаимодействует сложный белковый комплекс ТР 1 Ш, состоящий из 15— 20 субъединиц; непосредственно узнает данную последовательность ТВР, субъединица с молекулярной массой около 100— 120 к. Д. Компоненты фактора ТР 1 Ш образуют комплексы с многочисленными белками регуляторами и модулируют активность РНК полимеразы II. Действие минимального ТАТА связывающего комплекса осуществляется по схеме: ТВР связывается с ТАТА боксом; с образовавшимся комплексом связывается фактор ТРПВ; к ТРИ В в составе комплекса присоединяются фактор ТРПР и РНК полимераза II; образовавшийся минимальный комплекс присоединяет факторы ТРИ Е/Н, т. е. образуется полный инициаторный комплекс, который способен осуществить АТФ зависимую инициацию синтеза РНК.
 СААТ-боксы. Эти последовательности узнаются фактором транскрипции С/ЕВР. GС-мотивы. Данная группа регуляторных участков ДНК взаимодействует с фактором 8 Р 1.
СААТ-боксы. Эти последовательности узнаются фактором транскрипции С/ЕВР. GС-мотивы. Данная группа регуляторных участков ДНК взаимодействует с фактором 8 Р 1.
 Специфические регуляторные последовательности Цис-элементы являются участками связывания специфических белковых транскрипционных факторов; они расположены, как правило, на расстоянии до 300— 400 нуклеотидных пар перед точкой начала транскрипции. Их наборы индивидуальны для каждого гена, что обес печивает специфичность регуляции транскрипции. Связывание транскрипционных факторов может приводить как к усилению, так и к подавлению транскрипции.
Специфические регуляторные последовательности Цис-элементы являются участками связывания специфических белковых транскрипционных факторов; они расположены, как правило, на расстоянии до 300— 400 нуклеотидных пар перед точкой начала транскрипции. Их наборы индивидуальны для каждого гена, что обес печивает специфичность регуляции транскрипции. Связывание транскрипционных факторов может приводить как к усилению, так и к подавлению транскрипции.
 могут быть расположены на расстоянии до") Энхансеры. Особенности этих регуляторных последовательностей: (Всегда активизируют транскрипцию) могут быть расположены на расстоянии до нескольких тысяч нуклеотидных пар от кодирующей области гена; сохраняют свою активность при изменении ориентации; располагаются как перед, так и после кодирующей области. Последовательности, обладающие аналогичными свойствами, но ослабляющие транскрипцию, получили название «сайленсеры» . Действие энхансеров осуществляется, как и в предыдущем случае, благодаря связыванию
Энхансеры. Особенности этих регуляторных последовательностей: (Всегда активизируют транскрипцию) могут быть расположены на расстоянии до нескольких тысяч нуклеотидных пар от кодирующей области гена; сохраняют свою активность при изменении ориентации; располагаются как перед, так и после кодирующей области. Последовательности, обладающие аналогичными свойствами, но ослабляющие транскрипцию, получили название «сайленсеры» . Действие энхансеров осуществляется, как и в предыдущем случае, благодаря связыванию
 Типичный эукариотический транскрипционный фактор имеет два функциональных") Основные элементы структуры белковых транскрипционных факторов (транс-факторов) Типичный эукариотический транскрипционный фактор имеет два функциональных домена - ДНК-связывающий и трансактивирующий (или активирующий транскрипцию).
Основные элементы структуры белковых транскрипционных факторов (транс-факторов) Типичный эукариотический транскрипционный фактор имеет два функциональных домена - ДНК-связывающий и трансактивирующий (или активирующий транскрипцию).
 ДНК-связывающие домены обеспечивают две важнейшие функции факторов транскрипции — узнавание специфической последовательности и связывание с этой последовательностью. Транс-активирующие домены обеспечивают активацию (если данный фактор является активирующим) транскрипции; с этими же доменами связываются белкилиганды (активаторы, адаптеры, вспомогательные белки и т. д. ).
ДНК-связывающие домены обеспечивают две важнейшие функции факторов транскрипции — узнавание специфической последовательности и связывание с этой последовательностью. Транс-активирующие домены обеспечивают активацию (если данный фактор является активирующим) транскрипции; с этими же доменами связываются белкилиганды (активаторы, адаптеры, вспомогательные белки и т. д. ).
 расположены") Регуляция синтеза РНК-полимеразой I В каждом спейсере (длиной около 4 тыс. пар нуклеотидов) расположены многократно повторенные активаторы транскрипции (40— 60 пар нуклеотидов) и участки терминации / инициации Т. 188 5, 88 288 РНК Схема повторяющегося блока рибосомных генов.
Регуляция синтеза РНК-полимеразой I В каждом спейсере (длиной около 4 тыс. пар нуклеотидов) расположены многократно повторенные активаторы транскрипции (40— 60 пар нуклеотидов) и участки терминации / инициации Т. 188 5, 88 288 РНК Схема повторяющегося блока рибосомных генов.
 Последовательность событий при транскрипции рибосомных генов РНК полимераза I связывается с ДНК в области спейсера (этот процесс активируется в десятки раз регуляторными повторами в спейсере, причем данная активация не зависит от расстояния до этих по второв и от их ориентации); РНК полимераза I движется вдоль спейсерной ДНК к точке Т; по видимому, при этом происходит синтез РНК, которая очень быстро гидролизуется клеточными нуклеазами. в точке Т происходит терминация РНК, транскрибируемой с участка перед этой точкой и инициация новой молекулы РНК
Последовательность событий при транскрипции рибосомных генов РНК полимераза I связывается с ДНК в области спейсера (этот процесс активируется в десятки раз регуляторными повторами в спейсере, причем данная активация не зависит от расстояния до этих по второв и от их ориентации); РНК полимераза I движется вдоль спейсерной ДНК к точке Т; по видимому, при этом происходит синтез РНК, которая очень быстро гидролизуется клеточными нуклеазами. в точке Т происходит терминация РНК, транскрибируемой с участка перед этой точкой и инициация новой молекулы РНК
 Регуляция синтеза РНК-полимеразой III На данной схеме регуляции регуляторные участки расположены внутри кодирующих областей генов ▄ — Участки генов, регулирующие транскрипцию
Регуляция синтеза РНК-полимеразой III На данной схеме регуляции регуляторные участки расположены внутри кодирующих областей генов ▄ — Участки генов, регулирующие транскрипцию
 и") С этими участками связывается фактор транскрипции TFIIIA, затем другие факторы (2 3 молекулы) и РНК полимераза III, осуществляющая синтез РНК.
С этими участками связывается фактор транскрипции TFIIIA, затем другие факторы (2 3 молекулы) и РНК полимераза III, осуществляющая синтез РНК.