

12_Нуклеиновые кислоты.ppt
- Количество слайдов: 187
 НУКЛЕИНОВЫЕ КИСЛОТЫ
НУКЛЕИНОВЫЕ КИСЛОТЫ
 Нуклеиновые кислоты … • важнейшие биополимеры, осуществляющие хранение и передачу генетической информации в живой клетке дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты (РНК) ДНК – генетический материал большинства организмов В прокариотических клетках – хромосомная ДНК и внехромосомные ДНК – плазмиды В эукариотических клетках • • – – основная масса ДНК – в ядре, где связана с белками в хромосомах ДНК содержится в различных органеллах (митохондриях, хлоропластах)
Нуклеиновые кислоты … • важнейшие биополимеры, осуществляющие хранение и передачу генетической информации в живой клетке дезоксирибонуклеиновые кислоты (ДНК) и рибонуклеиновые кислоты (РНК) ДНК – генетический материал большинства организмов В прокариотических клетках – хромосомная ДНК и внехромосомные ДНК – плазмиды В эукариотических клетках • • – – основная масса ДНК – в ядре, где связана с белками в хромосомах ДНК содержится в различных органеллах (митохондриях, хлоропластах)
 РНК (м. РНК или и. РНК)") Нуклеиновые кислоты … • РНК – матричные (информационные) РНК (м. РНК или и. РНК) – рибосомные РНК (р. РНК) – транспортные РНК (т. РНК) – и ряд др. – входят в состав многих вирусов
Нуклеиновые кислоты … • РНК – матричные (информационные) РНК (м. РНК или и. РНК) – рибосомные РНК (р. РНК) – транспортные РНК (т. РНК) – и ряд др. – входят в состав многих вирусов
 Химический состав нуклеиновых кислот
Химический состав нуклеиновых кислот
 • При полном гидролизе нуклеиновых кислот образуются – пуриновые и пиримидиновые основания – углеводы (рибоза и дезоксирибоза) – фосфорная кислота
• При полном гидролизе нуклеиновых кислот образуются – пуриновые и пиримидиновые основания – углеводы (рибоза и дезоксирибоза) – фосфорная кислота
 Пиримидиновые основания • • • Производные пиримидина Сопряжение -электронов Бесцветное кристаллическое вещество, относящееся к классу диазинов Тпл 22 С, Ткип 124 С Хорошо растворим в воде Не дает щелочной реакции, но образует соли с сильными кислотами
Пиримидиновые основания • • • Производные пиримидина Сопряжение -электронов Бесцветное кристаллическое вещество, относящееся к классу диазинов Тпл 22 С, Ткип 124 С Хорошо растворим в воде Не дает щелочной реакции, но образует соли с сильными кислотами
 тимин (5 -метил-2, 4 диоксипиримидин)") Пиримидиновые основания • • • урацил (2, 4 -диоксипиримидин) тимин (5 -метил-2, 4 диоксипиримидин) цитозин (2 -окси-4 аминопиримидин)
Пиримидиновые основания • • • урацил (2, 4 -диоксипиримидин) тимин (5 -метил-2, 4 диоксипиримидин) цитозин (2 -окси-4 аминопиримидин)
 Пиримидиновые основания • • • Урацил входит в состав только РНК Тимин – в основном в ДНК и очень редко – в некоторые РНК Тимин и урацил имеют окисленный атом углерода в положении 4 В цитозине этот атом аминирован Все производные пиримидина имеют плоское строение
Пиримидиновые основания • • • Урацил входит в состав только РНК Тимин – в основном в ДНК и очень редко – в некоторые РНК Тимин и урацил имеют окисленный атом углерода в положении 4 В цитозине этот атом аминирован Все производные пиримидина имеют плоское строение
 Пиримидиновые основания • • • Минорные азотистые основания 5 -метилцитозин 5 -оксиметилцитозин
Пиримидиновые основания • • • Минорные азотистые основания 5 -метилцитозин 5 -оксиметилцитозин
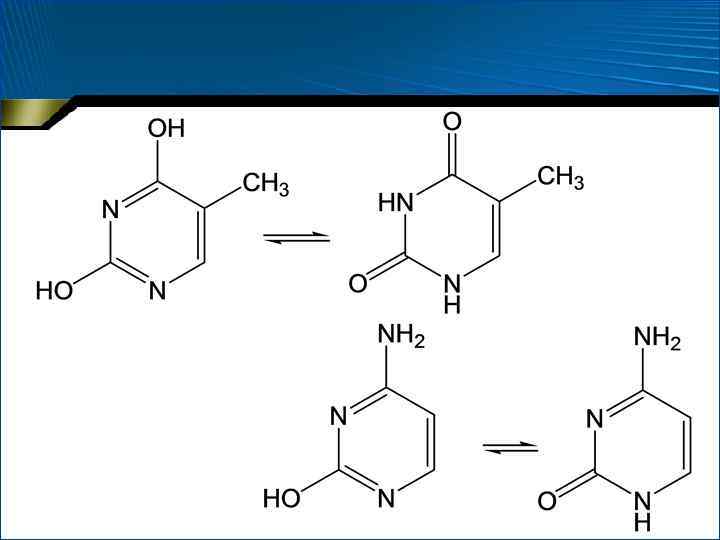
 Пиримидиновые основания • лактим-лактамная таутомерия
Пиримидиновые основания • лактим-лактамная таутомерия
 Пуриновые основания • • • Производные пурина Высокая степень сопряжения -электронов Бесцветное кристаллическое вещество, растворимое в воде
Пуриновые основания • • • Производные пурина Высокая степень сопряжения -электронов Бесцветное кристаллическое вещество, растворимое в воде
 Гуанин -амино-6 -оксипурин) (2") Пуриновые основания • • Аденин (6 -аминопурин) Гуанин -амино-6 -оксипурин) (2
Пуриновые основания • • Аденин (6 -аминопурин) Гуанин -амино-6 -оксипурин) (2
 Пуриновые основания • • Аденин и гуанин встречаются как в ДНК, так и в РНК Основное структурное отличие их – атом С 6 в гуанине окислен, а в аденине аминирован
Пуриновые основания • • Аденин и гуанин встречаются как в ДНК, так и в РНК Основное структурное отличие их – атом С 6 в гуанине окислен, а в аденине аминирован
 Пуриновые основания • • • Минорные азотистые основания 2 -метиладенин 1 -метилгуанин
Пуриновые основания • • • Минорные азотистые основания 2 -метиладенин 1 -метилгуанин
 Пуриновые основания • таутомерия
Пуриновые основания • таутомерия
 Углеводы
Углеводы
 Локализация в клетке ДНК и РНК Виды ДНК и РНК
Локализация в клетке ДНК и РНК Виды ДНК и РНК
") • Количество ДНК в клетке измеряется пикограммами (1 пг = 10– 12 г) и колеблется от 0, 01 пг у кишечной палочки до нескольких пг в клетках высших организмов
• Количество ДНК в клетке измеряется пикограммами (1 пг = 10– 12 г) и колеблется от 0, 01 пг у кишечной палочки до нескольких пг в клетках высших организмов
 Митохондриальная ДНК") Виды ДНК по месту локализации • • Ядерная ДНК (у эукариот преобладает) Митохондриальная ДНК (у животных), хлоропластная ДНК (у растений) • Центриольная ДНК • Эписомальная ДНК – Эписомы – внехромосомные единицы наследственности микроорганизмов • ДНК вирусов и фагов
Виды ДНК по месту локализации • • Ядерная ДНК (у эукариот преобладает) Митохондриальная ДНК (у животных), хлоропластная ДНК (у растений) • Центриольная ДНК • Эписомальная ДНК – Эписомы – внехромосомные единицы наследственности микроорганизмов • ДНК вирусов и фагов
 –") Виды РНК по функциональному значению, Мr и локализации • Транспортные РНК (т. РНК) – Мr от 17 до 35 тыс. – в гиалоплазме, ядерном соке, матриксе хлоропластов и митохондрий – Функция – перенос АК к месту синтеза белка • Рибосомальные РНК (р. РНК) – Мr • • – от 550 до 700 тыс. у РНК 30– 40 S субчастиц рибосом 1, 1– 1, 7 106 – у РНК 50– 60 S субчастиц рибосом 40 тыс. – у 5 S РНК ~50 тыс. – у 5, 8 S РНК Структурная основа рибосом
Виды РНК по функциональному значению, Мr и локализации • Транспортные РНК (т. РНК) – Мr от 17 до 35 тыс. – в гиалоплазме, ядерном соке, матриксе хлоропластов и митохондрий – Функция – перенос АК к месту синтеза белка • Рибосомальные РНК (р. РНК) – Мr • • – от 550 до 700 тыс. у РНК 30– 40 S субчастиц рибосом 1, 1– 1, 7 106 – у РНК 50– 60 S субчастиц рибосом 40 тыс. – у 5 S РНК ~50 тыс. – у 5, 8 S РНК Структурная основа рибосом
 Виды РНК по функциональному значению, Мr и локализации • Информационные, или матричные РНК (и. РНК, м. РНК) – Мr от 300 тыс. до 4 106 – Матричная функция в синтезе белка • Вирусные РНК – составные части вирусных и фаговых рибонуклеопротеидов, несут информацию, необходимую для размножения вируса в клетках хозяина • Ядерные РНК – предшественники всех видов РНК эукариотической клетки
Виды РНК по функциональному значению, Мr и локализации • Информационные, или матричные РНК (и. РНК, м. РНК) – Мr от 300 тыс. до 4 106 – Матричная функция в синтезе белка • Вирусные РНК – составные части вирусных и фаговых рибонуклеопротеидов, несут информацию, необходимую для размножения вируса в клетках хозяина • Ядерные РНК – предшественники всех видов РНК эукариотической клетки
 Строение структурных элементов нуклеиновых кислот
Строение структурных элементов нуклеиновых кислот
 • Мономерные звенья ДНК и РНК – остатки нуклеотидов, состоящих из остатков азотистого основания, рибозы или дезоксирибозы и фосфорной кислоты • Образуются при гидролизе нуклеиновых кислот водным раствором щелочи или ферментами • При гидролизе РНК – рибонуклеотиды • При гидролизе ДНК – дезоксирибонуклеотиды
• Мономерные звенья ДНК и РНК – остатки нуклеотидов, состоящих из остатков азотистого основания, рибозы или дезоксирибозы и фосфорной кислоты • Образуются при гидролизе нуклеиновых кислот водным раствором щелочи или ферментами • При гидролизе РНК – рибонуклеотиды • При гидролизе ДНК – дезоксирибонуклеотиды
") Рибонуклеотиды • Цитидин-3 -монофосфат (ЦМФ, цитидиновая кислота)
Рибонуклеотиды • Цитидин-3 -монофосфат (ЦМФ, цитидиновая кислота)
") Рибонуклеотиды • Уридин-3 -монофосфат (УМФ, уридиновая кислота)
Рибонуклеотиды • Уридин-3 -монофосфат (УМФ, уридиновая кислота)
") Рибонуклеотиды • Аденозин-3 -монофосфат (АМФ, адениловая кислота)
Рибонуклеотиды • Аденозин-3 -монофосфат (АМФ, адениловая кислота)
") Рибонуклеотиды • Гуанозин-3 -монофосфат (ГМФ, гуаниловая кислота)
Рибонуклеотиды • Гуанозин-3 -монофосфат (ГМФ, гуаниловая кислота)
") Дезоксирибонуклеотиды • Дезокситимидин-3 монофосфат (д. ТМФ, дезокситимидиновая кислота)
Дезоксирибонуклеотиды • Дезокситимидин-3 монофосфат (д. ТМФ, дезокситимидиновая кислота)
") Дезоксирибонуклеотиды • Дезоксицитидин-3 монофосфат (д. ЦМФ, дезоксицитидиновая кислота)
Дезоксирибонуклеотиды • Дезоксицитидин-3 монофосфат (д. ЦМФ, дезоксицитидиновая кислота)
") Дезоксирибонуклеотиды • Дезоксиаденозин-3 монофосфат (д. АМФ, дезоксиадениловая кислота)
Дезоксирибонуклеотиды • Дезоксиаденозин-3 монофосфат (д. АМФ, дезоксиадениловая кислота)
") Дезоксирибонуклеотиды • Дезоксигуанозин-3 монофосфат (д. ГМФ, дезоксигуаниловая кислота)
Дезоксирибонуклеотиды • Дезоксигуанозин-3 монофосфат (д. ГМФ, дезоксигуаниловая кислота)
 • Остаток фосфорной кислоты может быть у 5 атома С рибозы • формулы – самостоятельно
• Остаток фосфорной кислоты может быть у 5 атома С рибозы • формулы – самостоятельно
 • Гидролизуются до нуклеозидов – ЦМФ") • Мононуклеотиды – сильные кислоты (? ) • Гидролизуются до нуклеозидов – ЦМФ цитидин + Н 3 РО 4 – УМФ уридин + Н 3 РО 4 – АМФ аденозин + Н 3 РО 4 – ГМФ гуанозин + Н 3 РО 4 – д. ЦМФ дезоксицитидин + Н 3 РО 4 – д. ТМФ дезокситимидин + Н 3 РО 4 – д. АМФ дезоксиаденозин + Н 3 РО 4 – д. ГМФ дезоксигуанозин + Н 3 РО 4 • Дают реакции по типу многоатомных спиртов и за счет азотистых оснований
• Мононуклеотиды – сильные кислоты (? ) • Гидролизуются до нуклеозидов – ЦМФ цитидин + Н 3 РО 4 – УМФ уридин + Н 3 РО 4 – АМФ аденозин + Н 3 РО 4 – ГМФ гуанозин + Н 3 РО 4 – д. ЦМФ дезоксицитидин + Н 3 РО 4 – д. ТМФ дезокситимидин + Н 3 РО 4 – д. АМФ дезоксиаденозин + Н 3 РО 4 – д. ГМФ дезоксигуанозин + Н 3 РО 4 • Дают реакции по типу многоатомных спиртов и за счет азотистых оснований
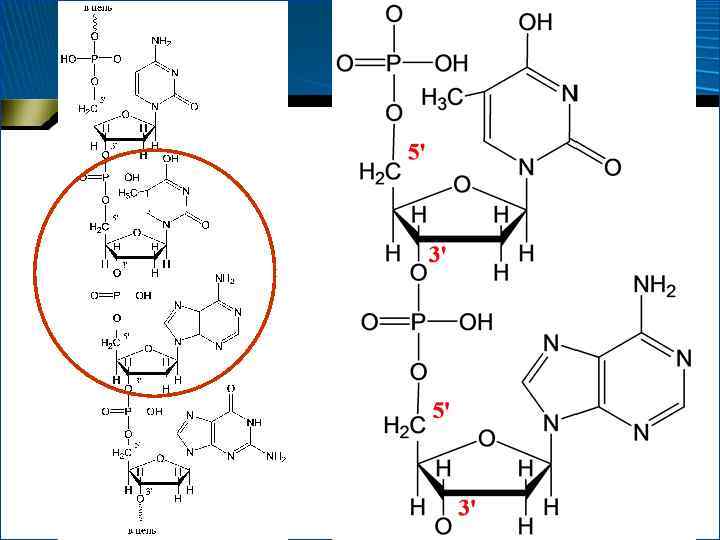
 • Мононуклеотиды, объединяясь, могут образовывать олиго- и полинуклеотиды, при этом образуются фосфодиэфирные связи между 3 -атомом С углевода одного мононуклеотида и 5 -атомом С углевода другого мононуклеотида
• Мононуклеотиды, объединяясь, могут образовывать олиго- и полинуклеотиды, при этом образуются фосфодиэфирные связи между 3 -атомом С углевода одного мононуклеотида и 5 -атомом С углевода другого мононуклеотида
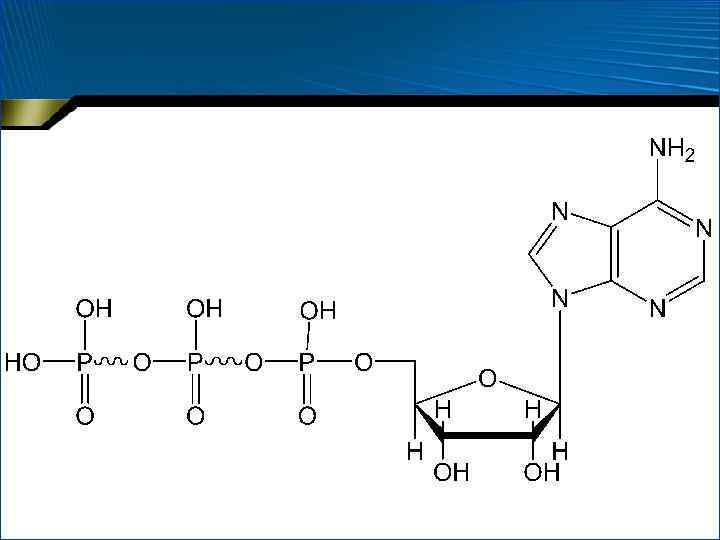
") • Кроме монофосфатов, могут быть ди- и трифосфаты • Аденозинтрифосфорная кислота (АТФ)
• Кроме монофосфатов, могут быть ди- и трифосфаты • Аденозинтрифосфорная кислота (АТФ)
 Нуклеиновые кислоты ДНК
Нуклеиновые кислоты ДНК
 ДНК … • • • дезоксирибонуклеиновая кислота – биологическая макромолекула, носитель генетической информации во всех эукариотических и прокариотических клетках и многих вирусах Длина молекулы ДНК – до сотен тысяч нм m(молекулы) – до 6 10– 12 г (Мr > 106) Из азотистых оснований в состав ДНК входят А, Г, Т, Ц; углеводный компонент – дезоксирибоза ДНК содержит всю информацию о наследственности, которая реализуется в синтезе белка
ДНК … • • • дезоксирибонуклеиновая кислота – биологическая макромолекула, носитель генетической информации во всех эукариотических и прокариотических клетках и многих вирусах Длина молекулы ДНК – до сотен тысяч нм m(молекулы) – до 6 10– 12 г (Мr > 106) Из азотистых оснований в состав ДНК входят А, Г, Т, Ц; углеводный компонент – дезоксирибоза ДНК содержит всю информацию о наследственности, которая реализуется в синтезе белка
 Первичная структура ДНК
Первичная структура ДНК
 Первичная структура ДНК … • последовательность дезоксирибонуклеотидов, связанных 3 – 5 -фосфодиэфирными связями – Один конец цепи ДНК несет 5 -ОН-группу и фосфат, а другой – 3 -ОН-группу – Последовательность оснований в ДНК всегда записывается в направлении от 5 к 3 -концу
Первичная структура ДНК … • последовательность дезоксирибонуклеотидов, связанных 3 – 5 -фосфодиэфирными связями – Один конец цепи ДНК несет 5 -ОН-группу и фосфат, а другой – 3 -ОН-группу – Последовательность оснований в ДНК всегда записывается в направлении от 5 к 3 -концу
 Генетический код
Генетический код
 • Одна из наиболее интригующих головоломок молекулярной биологии начала 1970 -х гг. – выяснение вопроса о том, каким образом четырехбуквенный «язык» ДНК переводится на двадцатибуквенную «речь» белков • Вопрос сводится к тому, из чего состоит этот таинственный код • Вероятнее всего, он заключается в определенной последовательности расположения нуклеотидов в молекуле ДНК
• Одна из наиболее интригующих головоломок молекулярной биологии начала 1970 -х гг. – выяснение вопроса о том, каким образом четырехбуквенный «язык» ДНК переводится на двадцатибуквенную «речь» белков • Вопрос сводится к тому, из чего состоит этот таинственный код • Вероятнее всего, он заключается в определенной последовательности расположения нуклеотидов в молекуле ДНК
 • До 1961 г. оставалось нерешенным, какие нуклеотиды ответственны за включение определенной аминокислоты в молекулу белка и какое число нуклеотидов определяет это включение • Теоретический разбор: – 1 нуклеотид (могут кодироваться только 4 аминокислоты) – 2 нуклеотида – дуплет (42 = 16 аминокислот) – Триплетный код – 43 = 64
• До 1961 г. оставалось нерешенным, какие нуклеотиды ответственны за включение определенной аминокислоты в молекулу белка и какое число нуклеотидов определяет это включение • Теоретический разбор: – 1 нуклеотид (могут кодироваться только 4 аминокислоты) – 2 нуклеотида – дуплет (42 = 16 аминокислот) – Триплетный код – 43 = 64
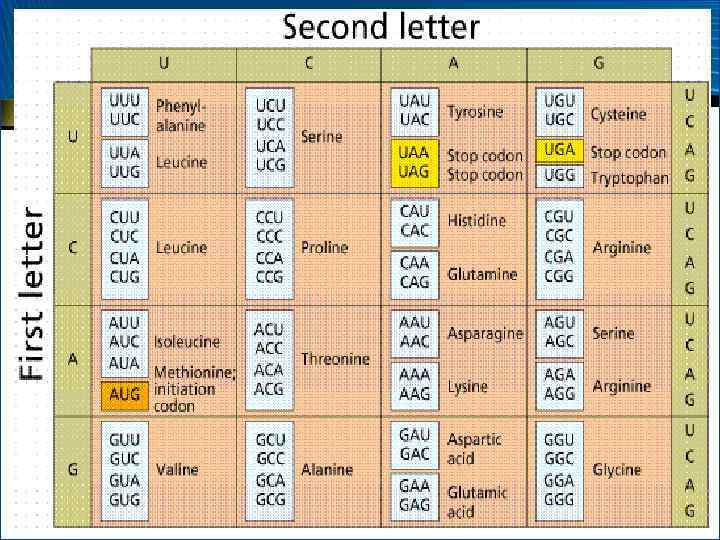
 • • М. У. Ниренберг Х. Г. Корана С. Очоа в 1966 г. представили доказательства не только состава, но и последовательности триплетов всех кодонов, ответственных за включение каждой из 20 аминокислот молекул белков
• • М. У. Ниренберг Х. Г. Корана С. Очоа в 1966 г. представили доказательства не только состава, но и последовательности триплетов всех кодонов, ответственных за включение каждой из 20 аминокислот молекул белков
 Свойства генетического кода 1. Триплетность 2. Вырожденность 3. Линейность, однонаправленность и неперекрываемость 4. Универсальность
Свойства генетического кода 1. Триплетность 2. Вырожденность 3. Линейность, однонаправленность и неперекрываемость 4. Универсальность
 Свойства генетического кода • • • Среди 64 кодонов 61 имеет смысл (кодирует определенную АК) 3 кодона бессмысленны (УАГ, УАА, УГА) Выполняют функцию окончания в синтезе белка, терминации синтеза Таким образом, генетический код – зависимость между основаниями в нуклеотидах ДНК и аминокислотами в молекуле белка Ген – участок ДНК, являющийся основной единицей наследственности
Свойства генетического кода • • • Среди 64 кодонов 61 имеет смысл (кодирует определенную АК) 3 кодона бессмысленны (УАГ, УАА, УГА) Выполняют функцию окончания в синтезе белка, терминации синтеза Таким образом, генетический код – зависимость между основаниями в нуклеотидах ДНК и аминокислотами в молекуле белка Ген – участок ДНК, являющийся основной единицей наследственности
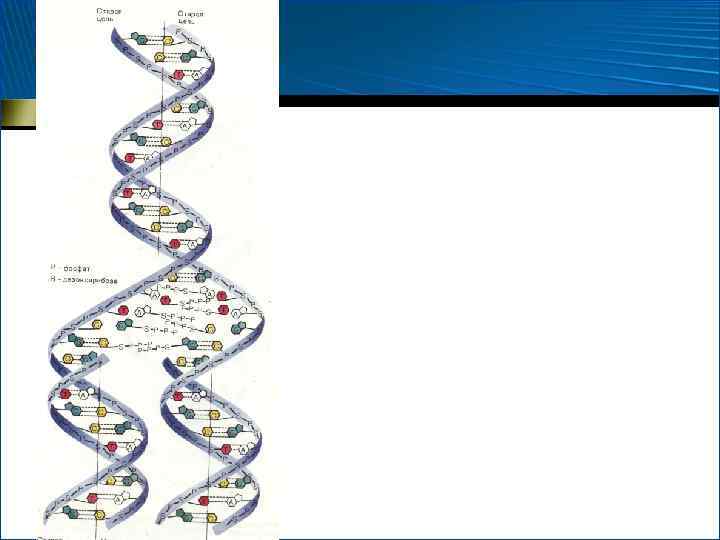
 Вторичная структура ДНК
Вторичная структура ДНК
 составлены парами") • Молекулы природной ДНК в подавляющем большинстве случаев (кроме ДНК фагов) составлены парами взаимозакрученных полидезоксирибонуклеотидных цепей, каждой из которых свойственно специфическое, но противоположное чередование нуклеотидных остатков
• Молекулы природной ДНК в подавляющем большинстве случаев (кроме ДНК фагов) составлены парами взаимозакрученных полидезоксирибонуклеотидных цепей, каждой из которых свойственно специфическое, но противоположное чередование нуклеотидных остатков
 • 1953 г. • Дж. Уотсон • Ф. Крик • Правая спираль • Двойная спираль
• 1953 г. • Дж. Уотсон • Ф. Крик • Правая спираль • Двойная спираль

") • Направление цепей взаимно противоположное (5 – 3 и 3 – 5 ) • Сахарофосфатный остов располагается по периферии двойной спирали, а азотистые основания находятся внутри, и их плоскости перпендикулярны оси спирали
• Направление цепей взаимно противоположное (5 – 3 и 3 – 5 ) • Сахарофосфатный остов располагается по периферии двойной спирали, а азотистые основания находятся внутри, и их плоскости перпендикулярны оси спирали
 • Э. Чаргафф • 1950 г.
• Э. Чаргафф • 1950 г.
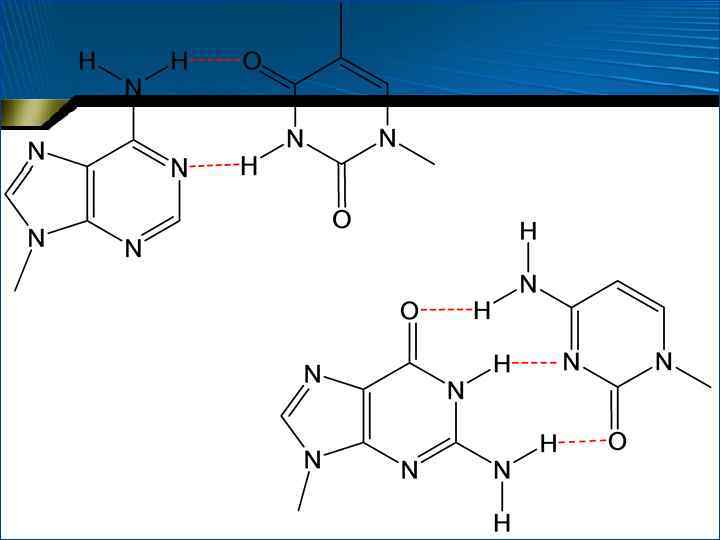
 Правила Чаргаффа 1. 2. • • • У ДНК молярная сумма Г и А (пуриновые основания) = молярной сумме Ц и Т (пиримидиновые основания). Эта закономерность несвойственна РНК В молекулах ДНК число остатков А всегда равно числу остатков Т. В таком же отношении находятся Г и Ц. В молекулах РНК этого нет Принцип комплементарности или дополнения Между азотистыми основаниями образуются специфические водородные связи, в результате чего осуществляется так называемое уотсонкриковское спаривание А всегда образует водородные связи с Т, а Г – с Ц
Правила Чаргаффа 1. 2. • • • У ДНК молярная сумма Г и А (пуриновые основания) = молярной сумме Ц и Т (пиримидиновые основания). Эта закономерность несвойственна РНК В молекулах ДНК число остатков А всегда равно числу остатков Т. В таком же отношении находятся Г и Ц. В молекулах РНК этого нет Принцип комплементарности или дополнения Между азотистыми основаниями образуются специфические водородные связи, в результате чего осуществляется так называемое уотсонкриковское спаривание А всегда образует водородные связи с Т, а Г – с Ц
 Правила Чаргаффа
Правила Чаргаффа
 Правила Чаргаффа • • Таким образом, более объемные пурины всегда спариваются с пиримидинами, имеющими меньшие размеры Это приводит к тому, что расстояния между С 1 атомами дезоксирибозы в двух цепях оказываются одинаковыми для АТ- и ЦГ-пар и равными 1, 085 нм В результате АТ- и ЦГ-пары включаются в двойную спираль без какого-либо заметного изменения геометрии остова К тому же такое спаривание оказывается и более выгодным энергетически, т. к. реализуется максимальное число возможных водородных связей (2 между А и Т и 3 – между Г и Ц)
Правила Чаргаффа • • Таким образом, более объемные пурины всегда спариваются с пиримидинами, имеющими меньшие размеры Это приводит к тому, что расстояния между С 1 атомами дезоксирибозы в двух цепях оказываются одинаковыми для АТ- и ЦГ-пар и равными 1, 085 нм В результате АТ- и ЦГ-пары включаются в двойную спираль без какого-либо заметного изменения геометрии остова К тому же такое спаривание оказывается и более выгодным энергетически, т. к. реализуется максимальное число возможных водородных связей (2 между А и Т и 3 – между Г и Ц)
 Спаривание комплементарных оснований в молекуле ДНК
Спаривание комплементарных оснований в молекуле ДНК
 Правила Чаргаффа 3. • Отношение суммы молярных концентраций Г и Ц к сумме молярных концентраций А и Т у ДНК и А и У у РНК сильно варьирует Это отношение для ДНК и РНК называется коэффициентом специфичности нуклеиновых кислот
Правила Чаргаффа 3. • Отношение суммы молярных концентраций Г и Ц к сумме молярных концентраций А и Т у ДНК и А и У у РНК сильно варьирует Это отношение для ДНК и РНК называется коэффициентом специфичности нуклеиновых кислот
 Геометрия двойной спирали
Геометрия двойной спирали
 • Соседние пары оснований – на расстоянии 0, 34 нм и повернуты на 36 вокруг оси спирали • На один виток спирали приходится 360 /36 = 10 пар оснований • Шаг спирали 10 0, 34 = 3, 4 (нм) • Диаметр двойной спирали ~20 нм • Желобки (большой и малый) • Это связано с тем, что сахарофосфатный остов расположен дальше от оси спирали, чем основания
• Соседние пары оснований – на расстоянии 0, 34 нм и повернуты на 36 вокруг оси спирали • На один виток спирали приходится 360 /36 = 10 пар оснований • Шаг спирали 10 0, 34 = 3, 4 (нм) • Диаметр двойной спирали ~20 нм • Желобки (большой и малый) • Это связано с тем, что сахарофосфатный остов расположен дальше от оси спирали, чем основания
 Стабильность двойной спирали
Стабильность двойной спирали
 • Водородные связи между основаниями. • Стэкинг-взаимодействия (от англ. stack – стог, куча, груда, стопка) направлены вдоль оси спирали • Гидрофобный эффект
• Водородные связи между основаниями. • Стэкинг-взаимодействия (от англ. stack – стог, куча, груда, стопка) направлены вдоль оси спирали • Гидрофобный эффект
 Полиморфизм ДНК
Полиморфизм ДНК
 Полиморфизм ДНК … • • способность двойной спирали принимать различные конформации По данным рентгеноструктурного анализа, ДНК может образовывать А-, В-, С-, Z-, Г- и другие формы, отличающиеся по ряду параметров
Полиморфизм ДНК … • • способность двойной спирали принимать различные конформации По данным рентгеноструктурного анализа, ДНК может образовывать А-, В-, С-, Z-, Г- и другие формы, отличающиеся по ряду параметров
 В-ДНК • • стандартная уотсон-криковская структура, в которой плоскости пар оснований перпендикулярны оси двойной спирали наиболее адекватна для репликационных процессов
В-ДНК • • стандартная уотсон-криковская структура, в которой плоскости пар оснований перпендикулярны оси двойной спирали наиболее адекватна для репликационных процессов
 А-ДНК • • В-ДНК переходит в А -форму при понижении влажности или активности воды в растворе (добавление спирта) Уменьшается расстояние между нуклеотидными парами вдоль оси спирали до 0, 26 нм при 11 нуклеотидных остатков на виток спирали
А-ДНК • • В-ДНК переходит в А -форму при понижении влажности или активности воды в растворе (добавление спирта) Уменьшается расстояние между нуклеотидными парами вдоль оси спирали до 0, 26 нм при 11 нуклеотидных остатков на виток спирали
 А-ДНК • • Диаметр спирали увеличивается, изменяется ширина и глубина бороздок, пары оснований образуют с осью спирали угол около 20 и смещены к периферии спирали Вследствие этого спираль похожа на пологую винтовую лестницу, а внутри нее возникает полость диаметром 0, 40 нм
А-ДНК • • Диаметр спирали увеличивается, изменяется ширина и глубина бороздок, пары оснований образуют с осью спирали угол около 20 и смещены к периферии спирали Вследствие этого спираль похожа на пологую винтовую лестницу, а внутри нее возникает полость диаметром 0, 40 нм
") А-ДНК • наиболее адекватна для процесса транскрипции (передачи информации)
А-ДНК • наиболее адекватна для процесса транскрипции (передачи информации)
 С-ДНК • • • получается при еще меньшей влажности, чем А-форма Расстояние между нуклеотидными парами в ней равно 0, 33 нм при 9, 3 нуклеотидных остатка на виток спирали Эта форма наиболее адекватна для упаковки ДНК в составе хроматина (хранения информации) и содержится в составе некоторых вирусов
С-ДНК • • • получается при еще меньшей влажности, чем А-форма Расстояние между нуклеотидными парами в ней равно 0, 33 нм при 9, 3 нуклеотидных остатка на виток спирали Эта форма наиболее адекватна для упаковки ДНК в составе хроматина (хранения информации) и содержится в составе некоторых вирусов
 Z-ДНК • левая спираль ДНК, образующаяся в растворах с высокой концентрацией солей или при добавлении спирта (А. Рич, 1979 г. )
Z-ДНК • левая спираль ДНК, образующаяся в растворах с высокой концентрацией солей или при добавлении спирта (А. Рич, 1979 г. )
 Z-ДНК • Отличительная особенность – повторяющейся единицей в ней является динуклеотид, в котором 2 соседних мононуклеотида находятся в разных конформациях
Z-ДНК • Отличительная особенность – повторяющейся единицей в ней является динуклеотид, в котором 2 соседних мононуклеотида находятся в разных конформациях
 Z-ДНК • • Диаметр молекулы 1, 8 нм, число нуклеотидных остатков в витке – 12, расстояние между ними – 0, 34 нм, наклон к оси спирали – 7 В Z-форме имеется лишь один желоб (бороздка)
Z-ДНК • • Диаметр молекулы 1, 8 нм, число нуклеотидных остатков в витке – 12, расстояние между ними – 0, 34 нм, наклон к оси спирали – 7 В Z-форме имеется лишь один желоб (бороздка)
 Z-ДНК • Биологическая роль Z-формы до конца не выяснена, но предполагается, что в этой форме реализуется участие ДНК в ряде метаболических процессов
Z-ДНК • Биологическая роль Z-формы до конца не выяснена, но предполагается, что в этой форме реализуется участие ДНК в ряде метаболических процессов
 SBS-ДНК • • от англ. side by side – бок о бок характеризуется отсутствием взаимозакручивания полидезоксирибонуклеотидных цепей, что обеспечивает легкое распаривание и расхождение цепей в процессе биосинтеза ДНК
SBS-ДНК • • от англ. side by side – бок о бок характеризуется отсутствием взаимозакручивания полидезоксирибонуклеотидных цепей, что обеспечивает легкое распаривание и расхождение цепей в процессе биосинтеза ДНК
 H-ДНК • • • тройная спираль, образующаяся при р. Н 4, 0 Биологическая роль не ясна Четверные спирали ДНК – один из вариантов перехода к третичной структуре ДНК
H-ДНК • • • тройная спираль, образующаяся при р. Н 4, 0 Биологическая роль не ясна Четверные спирали ДНК – один из вариантов перехода к третичной структуре ДНК
 • Биспиральные структуры в молекулах ДНК возникают не только при взаимодействии 2 -х комплементарных цепей, но и в пределах одной и той же цепи • Это происходит, когда в комплементарных цепях ДНК присутствуют палиндромы (от греч. палин – обратно, дроме – бегу): ГАЦАЦЦАГЦТГГТГТЦ ЦТГТГГТЦГАЦЦАЦАГ • Палиндромы – зоны «узнавания» структур ДНК ферментами и регуляторными белками • Эти участки могут спирализоваться сами на себя, образуя крестообразные структуры – шпильки
• Биспиральные структуры в молекулах ДНК возникают не только при взаимодействии 2 -х комплементарных цепей, но и в пределах одной и той же цепи • Это происходит, когда в комплементарных цепях ДНК присутствуют палиндромы (от греч. палин – обратно, дроме – бегу): ГАЦАЦЦАГЦТГГТГТЦ ЦТГТГГТЦГАЦЦАЦАГ • Палиндромы – зоны «узнавания» структур ДНК ферментами и регуляторными белками • Эти участки могут спирализоваться сами на себя, образуя крестообразные структуры – шпильки
 Третичная структура ДНК
Третичная структура ДНК
 • ДНК прокариот существует в линейной и кольцевой формах, третичная структура которых характеризуется спирализацией и суперспирализацией • У эукариот ДНК в третичной структуре находится в составе хроматина ядра и хромосом и характеризуется суперспирализацией нескольких уровней – длина всех молекул ДНК человека составляет в среднем 2 1010 км (расстояние от Земли до Солнца 1, 4 108 км) – длина ДНК одной клетки 2 м, а диаметр ядра, в котором она содержится, 5 мкм
• ДНК прокариот существует в линейной и кольцевой формах, третичная структура которых характеризуется спирализацией и суперспирализацией • У эукариот ДНК в третичной структуре находится в составе хроматина ядра и хромосом и характеризуется суперспирализацией нескольких уровней – длина всех молекул ДНК человека составляет в среднем 2 1010 км (расстояние от Земли до Солнца 1, 4 108 км) – длина ДНК одной клетки 2 м, а диаметр ядра, в котором она содержится, 5 мкм
 • Первый уровень суперспирализации ДНК в хроматине поддерживается белками-гистонами, образующими комплексы с молекулой ДНК – нуклеосомы • Нуклеосома образуется гистонами Н 2 а, Н 2 b, Н 3 и Н 4, образующими октамер; и около 200 нуклеотидными парами • Участки ДНК между нуклеосомами – линкерная ДНК – состоят из 6– 60 пар нуклеотидов и покрыты гистоном Н 1 • Таким образом на первом уровне суперспирализации образуется цепь из «бусинок» – нуклеосом
• Первый уровень суперспирализации ДНК в хроматине поддерживается белками-гистонами, образующими комплексы с молекулой ДНК – нуклеосомы • Нуклеосома образуется гистонами Н 2 а, Н 2 b, Н 3 и Н 4, образующими октамер; и около 200 нуклеотидными парами • Участки ДНК между нуклеосомами – линкерная ДНК – состоят из 6– 60 пар нуклеотидов и покрыты гистоном Н 1 • Таким образом на первом уровне суперспирализации образуется цепь из «бусинок» – нуклеосом
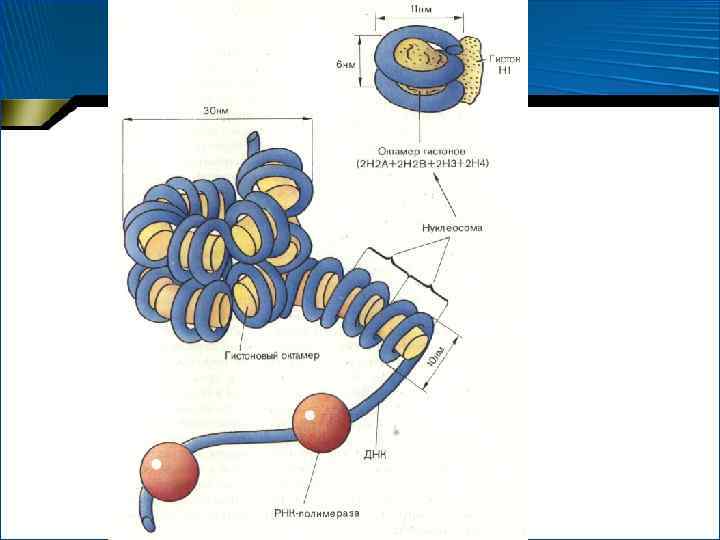
 образуются при") • Более высокие уровни суперспирализации (соленоид, диффузный кластеризованный и конденсированный хроматин) образуются при закручивании в спираль второго и последующего порядков цепочки нуклеосом вплоть до конденсации в хромосому
• Более высокие уровни суперспирализации (соленоид, диффузный кластеризованный и конденсированный хроматин) образуются при закручивании в спираль второго и последующего порядков цепочки нуклеосом вплоть до конденсации в хромосому
 • На один оборот спирали приходится: – в несуперспирализованной ДНК – 10 пар нуклеотидов – в нуклеосоме – 80 (уплотнение в 6– 7 раз) – в соленоиде – 6 нуклеосом на виток, 1200 нуклеотидных пар (уплотнение в 40 раз) – в каждой петле хроматина – 60000 нуклеотидных пар (в 680 раз) – в хромосоме – 1, 1 106 нуклеотидных пар (в 1, 2 104 раз)
• На один оборот спирали приходится: – в несуперспирализованной ДНК – 10 пар нуклеотидов – в нуклеосоме – 80 (уплотнение в 6– 7 раз) – в соленоиде – 6 нуклеосом на виток, 1200 нуклеотидных пар (уплотнение в 40 раз) – в каждой петле хроматина – 60000 нуклеотидных пар (в 680 раз) – в хромосоме – 1, 1 106 нуклеотидных пар (в 1, 2 104 раз)
 • При суперспирализации ДНК приобретает «скрытую» конформационную энергию, влияющую на любой процесс, происходящий с изменением числа витков двойной спирали • Это позволяет ферментам, локально расщепляющим ДНК (ДНК-полимеразы и др. ), легко связываться с суперспирализованной ДНК • Кроме того, появляется реальная возможность образования участков вторичной структуры, которые термодинамически невыгодны и практически не существуют в линейной или циклической форме • Переход ДНК в суперспирализованное состояние и обратно осуществляется при посредстве ферментов топоизомераз
• При суперспирализации ДНК приобретает «скрытую» конформационную энергию, влияющую на любой процесс, происходящий с изменением числа витков двойной спирали • Это позволяет ферментам, локально расщепляющим ДНК (ДНК-полимеразы и др. ), легко связываться с суперспирализованной ДНК • Кроме того, появляется реальная возможность образования участков вторичной структуры, которые термодинамически невыгодны и практически не существуют в линейной или циклической форме • Переход ДНК в суперспирализованное состояние и обратно осуществляется при посредстве ферментов топоизомераз
 Свойства ДНК
Свойства ДНК
 • ДНК – вещества белого цвета, волокнистого строения, плохо растворимые в воде (растворяются в виде солей щелочных металлов), но растворимы в крепких растворах солей • Растворы ДНК обладают высокой вязкостью и двойным лучепреломлением • Молекулы ДНК имеют отрицательный заряд и поэтому подвижны в электрическом поле • Оптически активны
• ДНК – вещества белого цвета, волокнистого строения, плохо растворимые в воде (растворяются в виде солей щелочных металлов), но растворимы в крепких растворах солей • Растворы ДНК обладают высокой вязкостью и двойным лучепреломлением • Молекулы ДНК имеют отрицательный заряд и поэтому подвижны в электрическом поле • Оптически активны
 • При температурах от 80 до 90 С происходит «плавление» ДНК, сопровождающееся изменением вязкости раствора и возрастанием поглощения в ультрафиолетовой части спектра (гиперхромный эффект)
• При температурах от 80 до 90 С происходит «плавление» ДНК, сопровождающееся изменением вязкости раствора и возрастанием поглощения в ультрафиолетовой части спектра (гиперхромный эффект)
 • ДНК химически инертна • Может вступать в реакции: – с ионами металлов (с Cu 2+ и Ме 4+ образует нерастворимые комплексы) – с полиаминами – спермидином H 2 N–(CH 2)3–NH–(CH 2)4–NH 2 и спермином H 2 N–(CH 2)3–NH–(CH 2)4–NH–(CH 2)3–NH 2 – алкилирование аминогрупп А, Г, Ц – дезаминирование Г и Ц • Последние два процесса – основа химического мутагенеза
• ДНК химически инертна • Может вступать в реакции: – с ионами металлов (с Cu 2+ и Ме 4+ образует нерастворимые комплексы) – с полиаминами – спермидином H 2 N–(CH 2)3–NH–(CH 2)4–NH 2 и спермином H 2 N–(CH 2)3–NH–(CH 2)4–NH–(CH 2)3–NH 2 – алкилирование аминогрупп А, Г, Ц – дезаминирование Г и Ц • Последние два процесса – основа химического мутагенеза
 ДНК") Репликация (редупликация, самоудвоение) ДНК
Репликация (редупликация, самоудвоение) ДНК
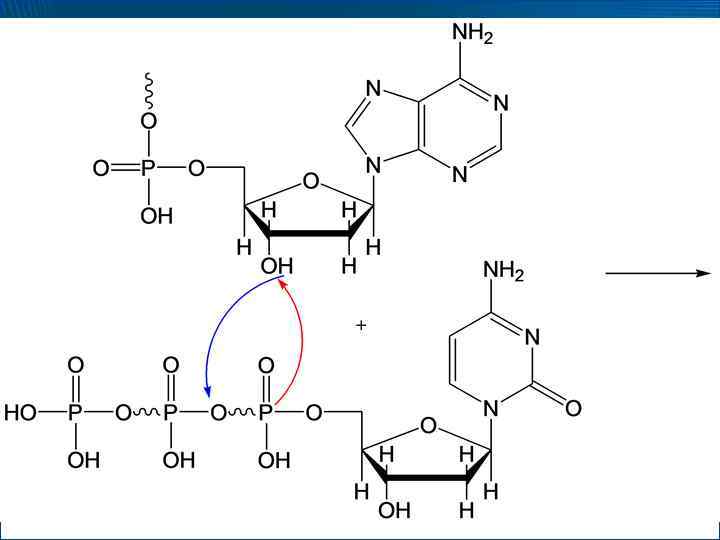
 – процесс,") • Репликация ДНК (от англ. replication – копирование; reduplication – удвоение) – процесс, при котором информация, закодированная в последовательности нуклеотидов родительской ДНК, передается с максимальной точностью дочерней ДНК • Биосинтез ДНК протекает из дезоксирибонуклеозидтрифосфатов при действии ДНК-полимераз, которые катализируют перенос дезоксирибонуклеотидных фрагментов от дезоксирибонуклеозид-5 -трифосфатов на группу –ОН 3 -концевого фрагмента растущей цепи
• Репликация ДНК (от англ. replication – копирование; reduplication – удвоение) – процесс, при котором информация, закодированная в последовательности нуклеотидов родительской ДНК, передается с максимальной точностью дочерней ДНК • Биосинтез ДНК протекает из дезоксирибонуклеозидтрифосфатов при действии ДНК-полимераз, которые катализируют перенос дезоксирибонуклеотидных фрагментов от дезоксирибонуклеозид-5 -трифосфатов на группу –ОН 3 -концевого фрагмента растущей цепи
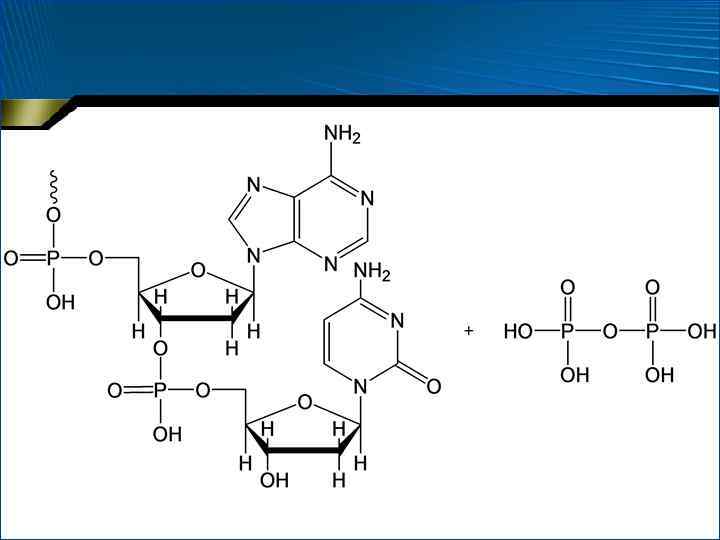
 Участие ДНК-полимеразы в репликации ДНК
Участие ДНК-полимеразы в репликации ДНК
 • Таким образом, происходят 2 сопряженных процесса – гидролиз макроэргической связи ( G << 0) и образование фосфодиэфирной связи ( G > 0)
• Таким образом, происходят 2 сопряженных процесса – гидролиз макроэргической связи ( G << 0) и образование фосфодиэфирной связи ( G > 0)
 –") Механизм репликации 1. • • • Инициация синтеза (от англ. initiation – введение) – начало синтеза Биосинтез ДНК начинается в строго определенном участке молекулы – участке ori (от англ. origin of replication) и от него распространяется в обе стороны Для инициации репликации ДНК расплетается в участке ori, образуя репликативную вилку В этом участвуют ферменты геликазы, перемещающиеся вдоль молекулы ДНК в двух направлениях
Механизм репликации 1. • • • Инициация синтеза (от англ. initiation – введение) – начало синтеза Биосинтез ДНК начинается в строго определенном участке молекулы – участке ori (от англ. origin of replication) и от него распространяется в обе стороны Для инициации репликации ДНК расплетается в участке ori, образуя репликативную вилку В этом участвуют ферменты геликазы, перемещающиеся вдоль молекулы ДНК в двух направлениях
 Участие геликазы в формировании репликативной вилки
Участие геликазы в формировании репликативной вилки
 Механизм репликации • В отличие от геликаз, ДНК-полимеразы могут перемещаться только от 3 - к 5 концу полинуклеотидной цепи, поэтому непрерывный синтез ДНК происходит только на одной нити ДНК – ведущей цепи
Механизм репликации • В отличие от геликаз, ДНК-полимеразы могут перемещаться только от 3 - к 5 концу полинуклеотидной цепи, поэтому непрерывный синтез ДНК происходит только на одной нити ДНК – ведущей цепи
 цепи 1) должен образоваться затравочный нуклеотидный фрагмент") Механизм репликации • На другой, запаздывающей (ведомой) цепи 1) должен образоваться затравочный нуклеотидный фрагмент – праймер (от англ. primer – запал), состоящий из рибонуклеотидов – В этом участвует фермент ДНК-зависимая РНКполимераза (праймаза) – Праймер образуется лишь в определенных участках запаздывающей цепи и состоит из нескольких десятков рибонуклеотидных звеньев 2) Синтез ДНК идет в противоположном направлении (фермент – ДНК-полимераза)
Механизм репликации • На другой, запаздывающей (ведомой) цепи 1) должен образоваться затравочный нуклеотидный фрагмент – праймер (от англ. primer – запал), состоящий из рибонуклеотидов – В этом участвует фермент ДНК-зависимая РНКполимераза (праймаза) – Праймер образуется лишь в определенных участках запаздывающей цепи и состоит из нескольких десятков рибонуклеотидных звеньев 2) Синтез ДНК идет в противоположном направлении (фермент – ДНК-полимераза)
 – продолжение") Механизм репликации 2. • • • Элонгация (от англ. elongate – удлинять) – продолжение синтеза На ведущей цепи происходит непрерывный синтез, а на запаздывающей – образуется новый праймер, т. е. синтез идет не непрерывно, а в виде фрагментов – фрагментов Т. Оказаки (1967 г. ), состоящих из примерно 200 нуклеотидных остатков (соответствует длине нуклеосомных участков ДНК) Как только второй фрагмент Оказаки достигает первый, праймер I удаляется с помощью РНКазы, а фрагменты «сшиваются» при посредстве ДНК-лигазы Далее процессы повторяются
Механизм репликации 2. • • • Элонгация (от англ. elongate – удлинять) – продолжение синтеза На ведущей цепи происходит непрерывный синтез, а на запаздывающей – образуется новый праймер, т. е. синтез идет не непрерывно, а в виде фрагментов – фрагментов Т. Оказаки (1967 г. ), состоящих из примерно 200 нуклеотидных остатков (соответствует длине нуклеосомных участков ДНК) Как только второй фрагмент Оказаки достигает первый, праймер I удаляется с помощью РНКазы, а фрагменты «сшиваются» при посредстве ДНК-лигазы Далее процессы повторяются
") Репликация лидирующей и запаздывающей цепей (I)
Репликация лидирующей и запаздывающей цепей (I)
") Репликация лидирующей и запаздывающей цепей (II)
Репликация лидирующей и запаздывающей цепей (II)
") Репликация лидирующей и запаздывающей цепей (III)
Репликация лидирующей и запаздывающей цепей (III)
 синтеза (от англ. termination – предел) Прекращение") Механизм репликации 3. • • Терминация (окончание) синтеза (от англ. termination – предел) Прекращение биосинтеза ДНК предположительно кодируется особой нуклеотидной последовательностью (палиндромом) на конце хромосомы Кроме того, репликация прекращается при встрече двух репликативных вилок
Механизм репликации 3. • • Терминация (окончание) синтеза (от англ. termination – предел) Прекращение биосинтеза ДНК предположительно кодируется особой нуклеотидной последовательностью (палиндромом) на конце хромосомы Кроме того, репликация прекращается при встрече двух репликативных вилок
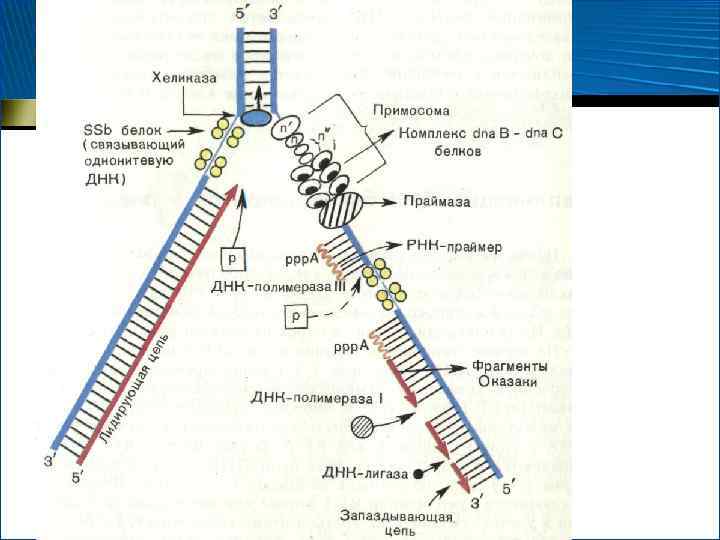
 Белковые факторы репликации 1. 2. 3. ДНК-связывающий белок ДНК-раскручивающий белок. Антагонист – ДНК-закручивающий белок Комплекс белковых факторов, обеспечивающих продвижение репликативной вилки и ее функционирование – праймосома
Белковые факторы репликации 1. 2. 3. ДНК-связывающий белок ДНК-раскручивающий белок. Антагонист – ДНК-закручивающий белок Комплекс белковых факторов, обеспечивающих продвижение репликативной вилки и ее функционирование – праймосома
 Закономерности репликации • • По принципу комплементарности, Молекула ДНК родительской клетки является матрицей, на которой синтезируются новые цепочки ДНК Гомологический механизм Полуконсервативный способ удвоения
Закономерности репликации • • По принципу комплементарности, Молекула ДНК родительской клетки является матрицей, на которой синтезируются новые цепочки ДНК Гомологический механизм Полуконсервативный способ удвоения
 Консервативная модель репликации • Консервативная репликация предполагает образование совершенно новой молекулы ДНК
Консервативная модель репликации • Консервативная репликация предполагает образование совершенно новой молекулы ДНК
 Полуконсервативная репликация • Semiconservative replication would produce two DNA molecules, each of which was composed of one-half of the parental DNA along with an entirely new complementary strand • In other words the new DNA would consist of one new and one old strand of DNA • The existing strands would serve as complementary templates for the new strand
Полуконсервативная репликация • Semiconservative replication would produce two DNA molecules, each of which was composed of one-half of the parental DNA along with an entirely new complementary strand • In other words the new DNA would consist of one new and one old strand of DNA • The existing strands would serve as complementary templates for the new strand
") Транскрипция (синтез РНК)
Транскрипция (синтез РНК)
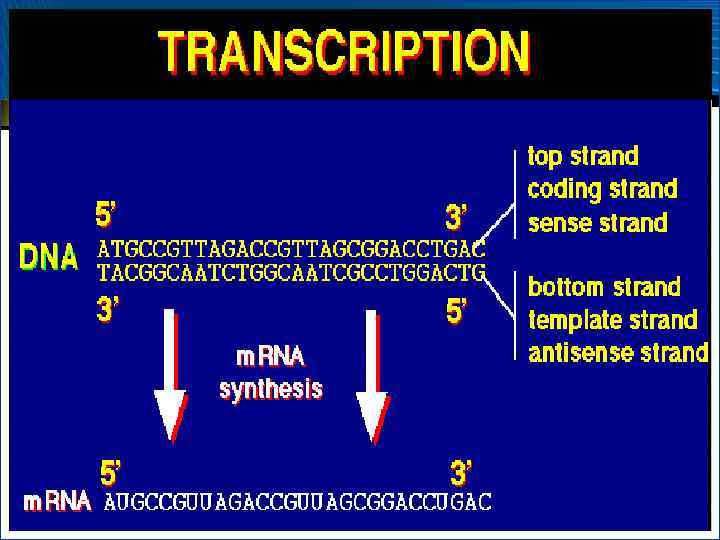
 У прокариот – одна РНК-полимераза, состоящая из 5 субъединиц У эукариот используется три РНК-полимеразы: полимераза I локализована в ядрышке, где она катализирует синтез р. РНК; полимераза II находится в нуклеоплазме и участвует в синтезе • Транскрипция РНК (от англ. transcription – м. РНК; полимераза III также локализована в переписывание) – процесс переноса генетической нуклеоплазме и участвует в синтезе т. РНК и 5 Sинформации от ДНК-матрицы к РНК р. РНК • Биосинтез РНК осуществляется при посредстве ДНК-зависимых РНК-полимераз, представляющих собой комплекс, состоящий из нескольких субъединиц – холофермент
У прокариот – одна РНК-полимераза, состоящая из 5 субъединиц У эукариот используется три РНК-полимеразы: полимераза I локализована в ядрышке, где она катализирует синтез р. РНК; полимераза II находится в нуклеоплазме и участвует в синтезе • Транскрипция РНК (от англ. transcription – м. РНК; полимераза III также локализована в переписывание) – процесс переноса генетической нуклеоплазме и участвует в синтезе т. РНК и 5 Sинформации от ДНК-матрицы к РНК р. РНК • Биосинтез РНК осуществляется при посредстве ДНК-зависимых РНК-полимераз, представляющих собой комплекс, состоящий из нескольких субъединиц – холофермент
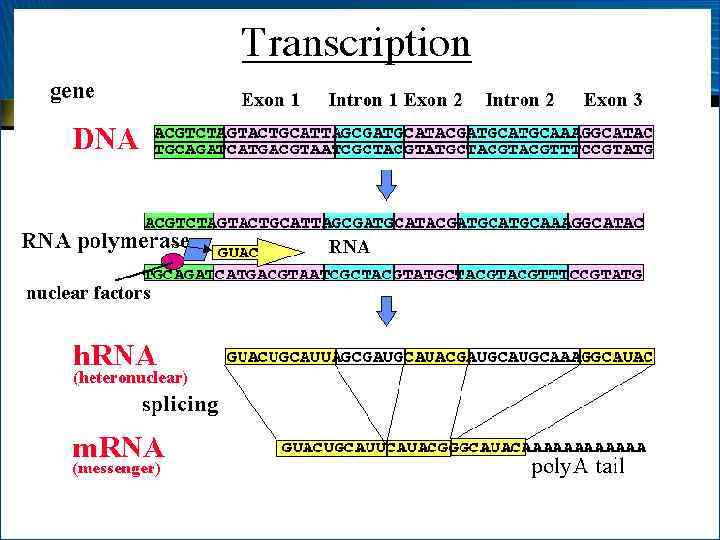
 • Та часть молекулы ДНК, которая копируется в процессе биосинтеза РНК на ней, называется транскриптоном • Транскриптон содержит информативные (экзоны) и неинформативные (интроны) зоны • Информативные зоны содержат информацию о РНК с определенной функциональной активностью • Неинформативная зона содержит регуляторные последовательности, с которыми взаимодействуют регуляторные белковые факторы, ускоряющие или замедляющие процесс транскрипции
• Та часть молекулы ДНК, которая копируется в процессе биосинтеза РНК на ней, называется транскриптоном • Транскриптон содержит информативные (экзоны) и неинформативные (интроны) зоны • Информативные зоны содержат информацию о РНК с определенной функциональной активностью • Неинформативная зона содержит регуляторные последовательности, с которыми взаимодействуют регуляторные белковые факторы, ускоряющие или замедляющие процесс транскрипции
 Этапы транскрипции 1. • Инициация. Транскрипция начинается на определенном участке ДНК – промоторе (от англ. promoter – то, что способствует чему-либо, подстрекатель) Промотор – участок молекулы ДНК, имеющий размер около 40 пар оснований, определенную нуклеотидную последовательность и расположенный непосредственно перед участком (сайтом, от англ. site – участок) инициации транскрипции
Этапы транскрипции 1. • Инициация. Транскрипция начинается на определенном участке ДНК – промоторе (от англ. promoter – то, что способствует чему-либо, подстрекатель) Промотор – участок молекулы ДНК, имеющий размер около 40 пар оснований, определенную нуклеотидную последовательность и расположенный непосредственно перед участком (сайтом, от англ. site – участок) инициации транскрипции
") Начало транскрипции (I)
Начало транскрипции (I)
 РНК-полимераза ІІ
РНК-полимераза ІІ
 Инициация транскрипции
Инициация транскрипции
 Этапы транскрипции • • Когда полимераза связывается с промотором, происходит локальное расплетание двойной спирали ДНК и образуется промоторный комплекс Синтез РНК всегда начинается с оснований А или Г «+» -цепи (5 – 3 ) и идет на одной цепи ДНК, а именно, на так называемой «–» цепи (3 – 5 ) в направлении 3 – 5
Этапы транскрипции • • Когда полимераза связывается с промотором, происходит локальное расплетание двойной спирали ДНК и образуется промоторный комплекс Синтез РНК всегда начинается с оснований А или Г «+» -цепи (5 – 3 ) и идет на одной цепи ДНК, а именно, на так называемой «–» цепи (3 – 5 ) в направлении 3 – 5
") Транскрипция РНК (II)
Транскрипция РНК (II)
 Этапы транскрипции 2. • • Элонгация наступает после присоединения примерно 8 рибонуклеотидов Синтез РНК идет из рибонуклеозилтрифосфатов При этом к растущей цепи присоединяются те рибонуклеозидтрифосфаты, которые обеспечивают правильное спаривание с «–» цепью ДНК (принцип комплементарности) Движущийся вдоль ДНК фермент действует подобно застежке-молнии «Раскрытая» ферментом область простирается только на несколько пар оснований
Этапы транскрипции 2. • • Элонгация наступает после присоединения примерно 8 рибонуклеотидов Синтез РНК идет из рибонуклеозилтрифосфатов При этом к растущей цепи присоединяются те рибонуклеозидтрифосфаты, которые обеспечивают правильное спаривание с «–» цепью ДНК (принцип комплементарности) Движущийся вдоль ДНК фермент действует подобно застежке-молнии «Раскрытая» ферментом область простирается только на несколько пар оснований
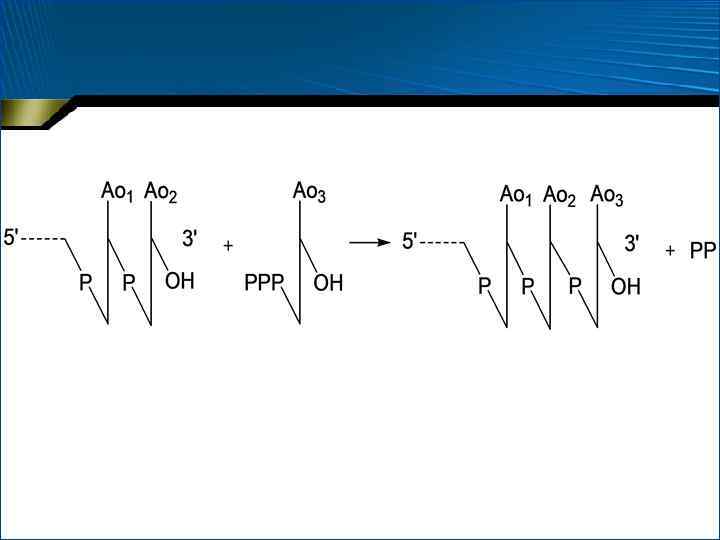
 цепи РНК происходит на специфических") Этапы транскрипции 3. • • • Терминация (прекращение роста) цепи РНК происходит на специфических участках ДНК – терминаторах Начало этих участков (сайтов) обычно обогащено ГЦ-парами, а остальная последовательность – АТ-парами ГЦ-богатый участок часто представляет собой палиндром Важную роль играет белковый фактор терминации – -белок
Этапы транскрипции 3. • • • Терминация (прекращение роста) цепи РНК происходит на специфических участках ДНК – терминаторах Начало этих участков (сайтов) обычно обогащено ГЦ-парами, а остальная последовательность – АТ-парами ГЦ-богатый участок часто представляет собой палиндром Важную роль играет белковый фактор терминации – -белок
") Окончание транскрипции (III)
Окончание транскрипции (III)
") Первичный транскрипт (IV)
Первичный транскрипт (IV)
 Схема транскрипции
Схема транскрипции
 – процесс созревания") Этапы транскрипции 4. • Посттранскрипционный процессинг (от англ. processing – обработка) – процесс созревания РНК, при котором первичный РНК-транскрипт (от англ. transcript – копия) модифицируется и превращается в зрелую РНК Характер и степень модификации зависит от типа РНК и включает:
Этапы транскрипции 4. • Посттранскрипционный процессинг (от англ. processing – обработка) – процесс созревания РНК, при котором первичный РНК-транскрипт (от англ. transcript – копия) модифицируется и превращается в зрелую РНК Характер и степень модификации зависит от типа РНК и включает:
 – удаление последовательностей РНК,") Этапы транскрипции • • Сплайсинг (от англ. splice – сращивать) – удаление последовательностей РНК, соответствующих интронам ДНК, и соединение участков, соответствующих экзонам Образование специфических фрагментов (зон): кэпструктуры (от англ. cap – шапочка) на 5 и поли. Апоследовательности – на 3 -конце – для м. РНК; последовательности ЦЦА на 3 -конце – для т. РНК Расщепление спейсеров (от англ. spacer – прокладка) – участков РНК, соединяющих участки, соответствующие 18 S-, 5, 8 S- и 28 S-р. РНК эукариот (16 S-, 23 S- и 5 S-р. РНК прокариот) Модификацию азотистых оснований с превращением их в минорные (для т. РНК)
Этапы транскрипции • • Сплайсинг (от англ. splice – сращивать) – удаление последовательностей РНК, соответствующих интронам ДНК, и соединение участков, соответствующих экзонам Образование специфических фрагментов (зон): кэпструктуры (от англ. cap – шапочка) на 5 и поли. Апоследовательности – на 3 -конце – для м. РНК; последовательности ЦЦА на 3 -конце – для т. РНК Расщепление спейсеров (от англ. spacer – прокладка) – участков РНК, соединяющих участки, соответствующие 18 S-, 5, 8 S- и 28 S-р. РНК эукариот (16 S-, 23 S- и 5 S-р. РНК прокариот) Модификацию азотистых оснований с превращением их в минорные (для т. РНК)
 Продукты транскрипции: пре-м. РНК
Продукты транскрипции: пре-м. РНК
 Продукты транскрипции: пре-р. РНК
Продукты транскрипции: пре-р. РНК
 Продукты транскрипции: пре-т. РНК
Продукты транскрипции: пре-т. РНК
 Центральная догма
Центральная догма
 Нуклеиновые кислоты РНК
Нуклеиновые кислоты РНК
 Рибонуклеиновые кислоты • повсеместно распространены в живой природе – во всех микроорганизмах, растительных и животных клетках и являются носителями наследственной информации во многих вирусах • обеспечивают реализацию в клетке наследственной информации, которая передается с помощью ДНК • содержат: из азотистых оснований – А, Г, У, Ц, углевод рибозу и имеют молекулярную массу (0, 5– 2 106)
Рибонуклеиновые кислоты • повсеместно распространены в живой природе – во всех микроорганизмах, растительных и животных клетках и являются носителями наследственной информации во многих вирусах • обеспечивают реализацию в клетке наследственной информации, которая передается с помощью ДНК • содержат: из азотистых оснований – А, Г, У, Ц, углевод рибозу и имеют молекулярную массу (0, 5– 2 106)
 Первичная структура РНК
Первичная структура РНК
 Первичная структура РНК … • последовательность рибонуклеотидов, связанных 3 – 5 -фосфодиэфирными связями (похожа на ДНК)
Первичная структура РНК … • последовательность рибонуклеотидов, связанных 3 – 5 -фосфодиэфирными связями (похожа на ДНК)
 Вторичная структура РНК
Вторичная структура РНК
 • Макромолекулы большинства природных РНК построены из одной полирибонуклеотидной цепи • Основной элемент вторичной структуры – сравнительно короткие двойные спирали, образованные комплементарными участками одной и той же цепи и перемежающиеся ее однотяжевыми сегментами • Полирибонуклеотидные цепи в таких двуспиральных структурах антипараллельны, а сами двойные спирали, находящиеся в А-форме, не идеальны: в них имеются дефекты в виде неспаренных остатков или не вписывающихся в двойную спираль однотяжевых петель (шпильки)
• Макромолекулы большинства природных РНК построены из одной полирибонуклеотидной цепи • Основной элемент вторичной структуры – сравнительно короткие двойные спирали, образованные комплементарными участками одной и той же цепи и перемежающиеся ее однотяжевыми сегментами • Полирибонуклеотидные цепи в таких двуспиральных структурах антипараллельны, а сами двойные спирали, находящиеся в А-форме, не идеальны: в них имеются дефекты в виде неспаренных остатков или не вписывающихся в двойную спираль однотяжевых петель (шпильки)
 и межплоскостными (стэкинг) взаимодействиями") • Стабильность двутяжевых районов поддерживается комплементарными (А–У и Г–Ц) и межплоскостными (стэкинг) взаимодействиями оснований • В однотяжевых участках наблюдаются сильные стэкинг-взаимодействия оснований, вследствие чего они стремятся принять конформацию однотяжевой спирали
• Стабильность двутяжевых районов поддерживается комплементарными (А–У и Г–Ц) и межплоскостными (стэкинг) взаимодействиями оснований • В однотяжевых участках наблюдаются сильные стэкинг-взаимодействия оснований, вследствие чего они стремятся принять конформацию однотяжевой спирали
 Третичная структура РНК
Третичная структура РНК
 • В физиологических условиях однотяжевые РНК характеризуются компактной и упорядоченной третичной структурой, которая возникает за счет взаимодействия шпилькообразных элементов их вторичной структуры • т. РНК – единственные представители природных полирибонуклеотидов, которые удалось закристаллизовать и изучить методом рентгеноструктурного анализа • Поэтому представления о принципах организации структуры РНК практически полностью основаны на данных о пространственной структуре т. РНК
• В физиологических условиях однотяжевые РНК характеризуются компактной и упорядоченной третичной структурой, которая возникает за счет взаимодействия шпилькообразных элементов их вторичной структуры • т. РНК – единственные представители природных полирибонуклеотидов, которые удалось закристаллизовать и изучить методом рентгеноструктурного анализа • Поэтому представления о принципах организации структуры РНК практически полностью основаны на данных о пространственной структуре т. РНК
 1. Элементы вторичной структуры РНК располагаются друг относительно друга так, чтобы обеспечить максимальный стэкинг оснований в макромолекуле в целом 2. Контакты между отдельными элементами вторичной структуры осуществляются за счет нескольких типов так называемых «третичных» внутримолекулярных взаимодействий – а) за счет образования дополнительных, зачастую неуотсон-криковских (например, Г–У) пар оснований между нуклеотидными остатками удаленных друг от друга (в первичной и вторичной структурах) однотяжевых участков и триплетов оснований между нуклеотидными остатками однотяжевых и двутяжевых элементов
1. Элементы вторичной структуры РНК располагаются друг относительно друга так, чтобы обеспечить максимальный стэкинг оснований в макромолекуле в целом 2. Контакты между отдельными элементами вторичной структуры осуществляются за счет нескольких типов так называемых «третичных» внутримолекулярных взаимодействий – а) за счет образования дополнительных, зачастую неуотсон-криковских (например, Г–У) пар оснований между нуклеотидными остатками удаленных друг от друга (в первичной и вторичной структурах) однотяжевых участков и триплетов оснований между нуклеотидными остатками однотяжевых и двутяжевых элементов
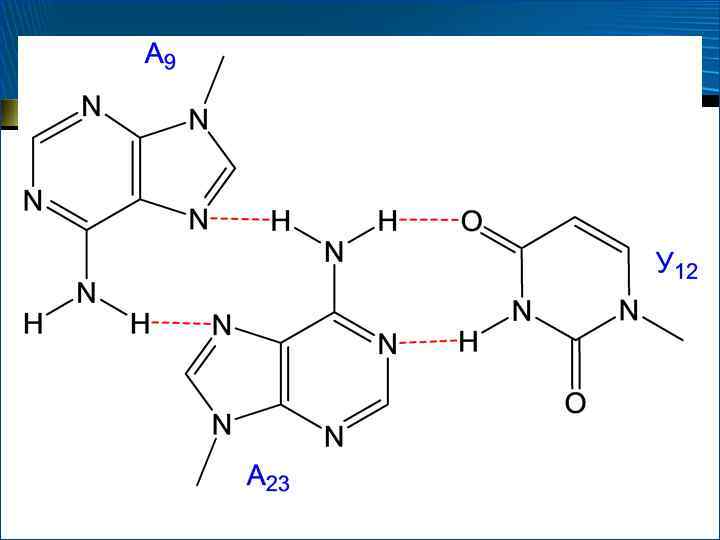
 за счет дополнительных ( «третичных» ) стэкинг-взаимодействий после интеркаляции (от англ. intercalation") – б) за счет дополнительных ( «третичных» ) стэкинг-взаимодействий после интеркаляции (от англ. intercalation – внедрение) оснований одного участка между двумя соседними основаниями другого однотяжевого участка – в) за счет образования дополнительных водородных связей между 2 -ОН-группами остатков рибозы и основаниями, а также другими группами сахарофосфатного остова 3. Третичная структура РНК стабилизирована ионами двухвалентных металлов, которые связываются не только с фосфатными группами, но и с основаниями
– б) за счет дополнительных ( «третичных» ) стэкинг-взаимодействий после интеркаляции (от англ. intercalation – внедрение) оснований одного участка между двумя соседними основаниями другого однотяжевого участка – в) за счет образования дополнительных водородных связей между 2 -ОН-группами остатков рибозы и основаниями, а также другими группами сахарофосфатного остова 3. Третичная структура РНК стабилизирована ионами двухвалентных металлов, которые связываются не только с фосфатными группами, но и с основаниями
 Классы РНК
Классы РНК
 – рибосомальные") • По составу, структуре и функциям различают – транспортные (т. РНК) – рибосомальные (р. РНК) – информационные (и. РНК, или РНК-посредники, мессенджер-РНК, от англ. messenger – курьер, посыльный, м. РНК, матричные РНК) – ядерные (я. РНК, или гетерогенные ядерные РНК – гя. РНК)
• По составу, структуре и функциям различают – транспортные (т. РНК) – рибосомальные (р. РНК) – информационные (и. РНК, или РНК-посредники, мессенджер-РНК, от англ. messenger – курьер, посыльный, м. РНК, матричные РНК) – ядерные (я. РНК, или гетерогенные ядерные РНК – гя. РНК)
 Транспортные РНК
Транспортные РНК
 • Основная функция т. РНК – перенос аминокислот к местам синтеза белка – к рибосомам • Каждая индивидуальная т. РНК способная переносить единственную аминокислоту • Конкретные т. РНК называют по имени этой аминокислоты – аланиновая т. РНК, или т. РНКала – Если одна и та же аминокислота переносится несколькими т. РНК, то последние называют изоакцепторными и нумеруют (т. РНКвал 1, т. РНКвал 2)
• Основная функция т. РНК – перенос аминокислот к местам синтеза белка – к рибосомам • Каждая индивидуальная т. РНК способная переносить единственную аминокислоту • Конкретные т. РНК называют по имени этой аминокислоты – аланиновая т. РНК, или т. РНКала – Если одна и та же аминокислота переносится несколькими т. РНК, то последние называют изоакцепторными и нумеруют (т. РНКвал 1, т. РНКвал 2)
 • т. РНК богаты минорными нуклеотидными остатками и содержат нуклеозиды и нуклеотиды своеобразного строения Псевдоуридиловая кислота ( )
• т. РНК богаты минорными нуклеотидными остатками и содержат нуклеозиды и нуклеотиды своеобразного строения Псевдоуридиловая кислота ( )
 • минорные нуклеотидные остатки – защищают т. РНК от атаки рибонуклеазами – принимают участие в кодировании аминокислот – важны для узнавания аминоацил-т. РНКсинтетазой нужной т. РНК • В т. РНК заметно преобладает (Г+Ц) над (А+У) • Молекулярные массы т. РНК 17– 35 тыс.
• минорные нуклеотидные остатки – защищают т. РНК от атаки рибонуклеазами – принимают участие в кодировании аминокислот – важны для узнавания аминоацил-т. РНКсинтетазой нужной т. РНК • В т. РНК заметно преобладает (Г+Ц) над (А+У) • Молекулярные массы т. РНК 17– 35 тыс.
 Первичная структура т. РНК • • Р. У. Холли, 1965 г. , т. РНКала пекарских дрожжей А. А. Баев, 1967 г. , т. РНКвал дрожжей
Первичная структура т. РНК • • Р. У. Холли, 1965 г. , т. РНКала пекарских дрожжей А. А. Баев, 1967 г. , т. РНКвал дрожжей
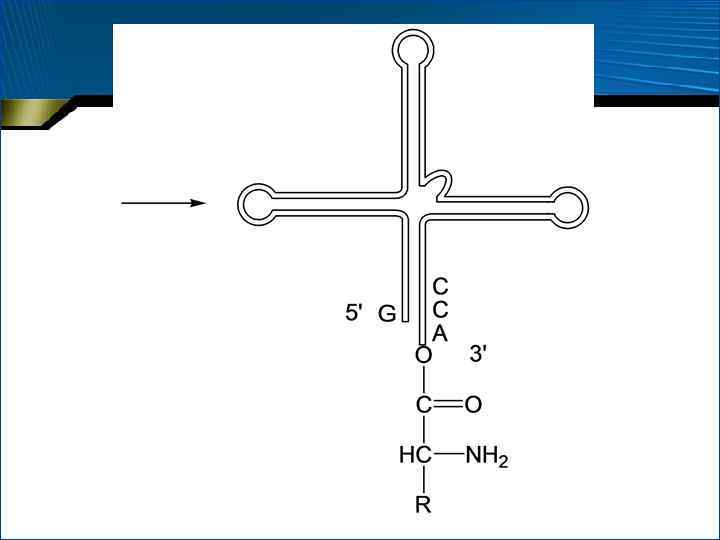
 Закономерности первичной структуры т. РНК • • Все т. РНК начинаются с фосфорилированного 5 -конца; в 75% случаев первым основанием является Г На 3 -конце всегда присутствуют три основания – ЦЦА, причем остаток А служит для связывания (акцептирования) аминокислоты В первичной структуре т. РНК имеются гомологичные блоки, крайне близкие по чередованию нуклеотидных остатков Кроме того, минорные нуклеотидные остатки закономерно распределены в первичной структуре т. РНК.
Закономерности первичной структуры т. РНК • • Все т. РНК начинаются с фосфорилированного 5 -конца; в 75% случаев первым основанием является Г На 3 -конце всегда присутствуют три основания – ЦЦА, причем остаток А служит для связывания (акцептирования) аминокислоты В первичной структуре т. РНК имеются гомологичные блоки, крайне близкие по чередованию нуклеотидных остатков Кроме того, минорные нуклеотидные остатки закономерно распределены в первичной структуре т. РНК.
 конец АС –") Вторичная структура т. РНК • • • АА – аминоацильный (акцепторный) конец АС – антикодоновая петля Т – псевдоуридиловая петля, в ней содержится последовательность ГТ Ц D – дигидроуридиловая петля V – вариабельная (добавочная) петля
Вторичная структура т. РНК • • • АА – аминоацильный (акцепторный) конец АС – антикодоновая петля Т – псевдоуридиловая петля, в ней содержится последовательность ГТ Ц D – дигидроуридиловая петля V – вариабельная (добавочная) петля
 Третичная структура т. РНК • • • Стала ясна после рентгеноструктурного анализа т. РНКфен из дрожжей А. Рич А. Клуг
Третичная структура т. РНК • • • Стала ясна после рентгеноструктурного анализа т. РНКфен из дрожжей А. Рич А. Клуг
 Рибосомальная РНК
Рибосомальная РНК
 • У всех организмов найдено три вида р. РНК, отличающиеся по молекулярным массам и локализации в рибосомах, обязательной составной частью которых они являются • Две р. РНК высокомолекулярны • третья сравнительно низкополимерна • Кроме того, в рибосомах эукариот присутствует еще одна низкомолекулярная р. РНК
• У всех организмов найдено три вида р. РНК, отличающиеся по молекулярным массам и локализации в рибосомах, обязательной составной частью которых они являются • Две р. РНК высокомолекулярны • третья сравнительно низкополимерна • Кроме того, в рибосомах эукариот присутствует еще одна низкомолекулярная р. РНК
 • Различают: – 16– 18 S р. РНК, локализованную в 30– 40 S субчастицах рибосом (Мr = 0, 55– 0, 79 106) – 23– 29 S р. РНК, локализованную в 50– 60 S субчастицах рибосом (Мr = 1, 07– 1, 6 106) – 5 S р. РНК присутствует во всех рибосомах и локализована в 50– 60 S субчастицах (Мr = 40 тыс. ) – 5, 8 S р. РНК характерна только для эукариотических рибосом (Мr ~ 50 тыс. )
• Различают: – 16– 18 S р. РНК, локализованную в 30– 40 S субчастицах рибосом (Мr = 0, 55– 0, 79 106) – 23– 29 S р. РНК, локализованную в 50– 60 S субчастицах рибосом (Мr = 1, 07– 1, 6 106) – 5 S р. РНК присутствует во всех рибосомах и локализована в 50– 60 S субчастицах (Мr = 40 тыс. ) – 5, 8 S р. РНК характерна только для эукариотических рибосом (Мr ~ 50 тыс. )
 Первичная структура р. РНК • • • Нуклеотидный состав высокополимерных р. РНК варьирует в довольно широких пределах и по мере усложнения организма все более смещается в сторону преобладания ГЦ-пар р. РНК митохондрий отличаются резким преобладанием АУ-пар (митохондриальная РНК) Высокомолекулярные р. РНК содержат в 2– 5 раз меньше минорных оснований, чем т. РНК
Первичная структура р. РНК • • • Нуклеотидный состав высокополимерных р. РНК варьирует в довольно широких пределах и по мере усложнения организма все более смещается в сторону преобладания ГЦ-пар р. РНК митохондрий отличаются резким преобладанием АУ-пар (митохондриальная РНК) Высокомолекулярные р. РНК содержат в 2– 5 раз меньше минорных оснований, чем т. РНК
 Первичная структура р. РНК • • В 5 S р. РНК совершенно нет минорных оснований (иногда – только псевдоуридиловая кислота) 5 S р. РНК относится к ГЦ-типу 5 S р. РНК содержит 120 нуклеотидных остатков (за редким исключением), последовательность которых в незначительной мере отличается у разных организмов. Молекулы 5, 8 S р. РНК в подавляющем большинстве случаев содержат 158 нуклеотидных остатков
Первичная структура р. РНК • • В 5 S р. РНК совершенно нет минорных оснований (иногда – только псевдоуридиловая кислота) 5 S р. РНК относится к ГЦ-типу 5 S р. РНК содержит 120 нуклеотидных остатков (за редким исключением), последовательность которых в незначительной мере отличается у разных организмов. Молекулы 5, 8 S р. РНК в подавляющем большинстве случаев содержат 158 нуклеотидных остатков
 Первичная структура р. РНК • • Работы по выяснению первичной структуры высокомолекулярных р. РНК начаты в 1974 г. К настоящему времени раскрыты первичные структуры более сотни 16– 18 S р. РНК и некоторых 23– 29 S р. РНК
Первичная структура р. РНК • • Работы по выяснению первичной структуры высокомолекулярных р. РНК начаты в 1974 г. К настоящему времени раскрыты первичные структуры более сотни 16– 18 S р. РНК и некоторых 23– 29 S р. РНК
 Вторичная структура р. РНК • характеризуется спирализацией самой на себя полирибонуклеотидной цепи за счет взаимодействия комплементарных оснований, в результате в молекуле возникает различное число биспиральных участков (> в 5 S и 5, 8 S и < в 16– 18 S р. РНК)
Вторичная структура р. РНК • характеризуется спирализацией самой на себя полирибонуклеотидной цепи за счет взаимодействия комплементарных оснований, в результате в молекуле возникает различное число биспиральных участков (> в 5 S и 5, 8 S и < в 16– 18 S р. РНК)
 Третичная структура р. РНК • образуется за счет укладки плоскостных биспиральных и линейных участков молекул в более компактные структуры
Третичная структура р. РНК • образуется за счет укладки плоскостных биспиральных и линейных участков молекул в более компактные структуры
 Функциональная роль р. РНК • 16– 18 S и 23– 29 S р. РНК – являются структурной основой для формирования рибонуклеопротеинового тяжа, который дает начало 30– 40 S и 50– 60 S субчастицам рибосом – взаимодействуют с м. РНК и аминоацил-т. РНК – их участки распознаются белковыми факторами, принимающими участие в синтезе белка на рибосомах – могут контактировать друг с другом при образовании 70– 80 S рибосом из 30– 40 S и 50– 60 S субчастиц и формировании активного центра рибосомы • 5 S р. РНК участвует в связывании т. РНК с рибосомой за счет псевдоуридиловой петли
Функциональная роль р. РНК • 16– 18 S и 23– 29 S р. РНК – являются структурной основой для формирования рибонуклеопротеинового тяжа, который дает начало 30– 40 S и 50– 60 S субчастицам рибосом – взаимодействуют с м. РНК и аминоацил-т. РНК – их участки распознаются белковыми факторами, принимающими участие в синтезе белка на рибосомах – могут контактировать друг с другом при образовании 70– 80 S рибосом из 30– 40 S и 50– 60 S субчастиц и формировании активного центра рибосомы • 5 S р. РНК участвует в связывании т. РНК с рибосомой за счет псевдоуридиловой петли
 Информационные РНК
Информационные РНК
 • 1958 г. А. Н. Белозерский, А. С. Спирин • 1962 г. , м. РНК выделена в свободном виде • Молекулярные массы м. РНК варьируют от нескольких сотен тыс. до нескольких млн. • Функция и. РНК – копирование генетической информации с ДНК и перевод ее в аминокислотную последовательность (и. РНК – матрица для синтеза белка)
• 1958 г. А. Н. Белозерский, А. С. Спирин • 1962 г. , м. РНК выделена в свободном виде • Молекулярные массы м. РНК варьируют от нескольких сотен тыс. до нескольких млн. • Функция и. РНК – копирование генетической информации с ДНК и перевод ее в аминокислотную последовательность (и. РНК – матрица для синтеза белка)
 • Моноцистронные и. РНК • Полицистронные и. РНК • Нуклеотидный состав крайне разнообразен
• Моноцистронные и. РНК • Полицистронные и. РНК • Нуклеотидный состав крайне разнообразен
 Первичная структура и. РНК • В составе и. РНК есть информативные, работающие как матрицы в процессе биосинтеза белка, зоны и неинформативные участки
Первичная структура и. РНК • В составе и. РНК есть информативные, работающие как матрицы в процессе биосинтеза белка, зоны и неинформативные участки
 Неинформативные участки и. РНК • Полиадениловые фрагменты длиной от 50 до 400 нуклеотидных остатков (кроме гистоновых и. РНК) на 3 -конце, которые – участвуют в процессе созревания и. РНК – предопределяют время жизни и. РНК – способствуют переносу ее из ядра в цитоплазму – принимают участие в трансляции
Неинформативные участки и. РНК • Полиадениловые фрагменты длиной от 50 до 400 нуклеотидных остатков (кроме гистоновых и. РНК) на 3 -конце, которые – участвуют в процессе созревания и. РНК – предопределяют время жизни и. РНК – способствуют переносу ее из ядра в цитоплазму – принимают участие в трансляции
 Неинформативные участки и. РНК • Небольшие повторяющиеся последовательности по 30 нуклеотидных остатков, располагающиеся неподалеку от полиаденилового фрагмента и необходимые для взаимодействия и. РНК с рибосомой или отдельными белковыми факторами
Неинформативные участки и. РНК • Небольшие повторяющиеся последовательности по 30 нуклеотидных остатков, располагающиеся неподалеку от полиаденилового фрагмента и необходимые для взаимодействия и. РНК с рибосомой или отдельными белковыми факторами
 Неинформативные участки и. РНК • Кэп на 5 -конце – нуклеотидная последовательность, азотистые основания в которой метилированы, а один из нуклеотидных остатков – 7 -метил. Г, присоединен через трифосфатную группировку • Кэп нужен – для защиты и. РНК от экзонуклеаз – для связывания белковых факторов при взаимодействии с р. РНК – Играет сигнальную роль присоединении и. РНК к рибосоме – участвует в трансляции
Неинформативные участки и. РНК • Кэп на 5 -конце – нуклеотидная последовательность, азотистые основания в которой метилированы, а один из нуклеотидных остатков – 7 -метил. Г, присоединен через трифосфатную группировку • Кэп нужен – для защиты и. РНК от экзонуклеаз – для связывания белковых факторов при взаимодействии с р. РНК – Играет сигнальную роль присоединении и. РНК к рибосоме – участвует в трансляции
 начинается с АУГ или ГУГ на 5 -конце и") • Информативный фрагмент (транслируемый) начинается с АУГ или ГУГ на 5 -конце и заканчивается УГА, УАГ или УАА на 3 -конце • АУГ или ГУГ соответствует антикодон формилметионил-т. РНК; УАА, УГА и УАГ являются терминирующими трансляцию кодонами
• Информативный фрагмент (транслируемый) начинается с АУГ или ГУГ на 5 -конце и заканчивается УГА, УАГ или УАА на 3 -конце • АУГ или ГУГ соответствует антикодон формилметионил-т. РНК; УАА, УГА и УАГ являются терминирующими трансляцию кодонами
 Вторичная и третичная структуры и. РНК • • Вторичная структура полностью не выяснена, но известно, что полинуклеотидная цепь и. РНК спирализована сама на себя О третичной структуре и. РНК известно только то, что она менее компактна, чем у р. РНК
Вторичная и третичная структуры и. РНК • • Вторичная структура полностью не выяснена, но известно, что полинуклеотидная цепь и. РНК спирализована сама на себя О третичной структуре и. РНК известно только то, что она менее компактна, чем у р. РНК
 Вирусы
Вирусы
 Вирусы… • • • частицы, построенные из одной или нескольких молекул нуклеиновой кислоты и нескольких белков, иногда некоторых других дополнительных компонентов, например, фосфолипидов Вне клеток никаких признаков жизнедеятельности не проявляют Однако, проникая внутрь клеток, с помощью своей нуклеиновой кислоты перепрограммируют работу клетки и в ней начинается размножение вирусной нуклеиновой кислоты и производство вирусных белков
Вирусы… • • • частицы, построенные из одной или нескольких молекул нуклеиновой кислоты и нескольких белков, иногда некоторых других дополнительных компонентов, например, фосфолипидов Вне клеток никаких признаков жизнедеятельности не проявляют Однако, проникая внутрь клеток, с помощью своей нуклеиновой кислоты перепрограммируют работу клетки и в ней начинается размножение вирусной нуклеиновой кислоты и производство вирусных белков
 • • • Процесс, как правило, завершается формированием зрелых частиц вируса и разрушением плазматической мембраны клетки, после чего вышедшие наружу вирусы получают возможность инфицировать новые хозяйственные клетки Существуют вирусы, паразитирующие в растительных и животных клетках, в том числе в клетках человека, вызывая различные вирусные заболевания Вирусы могут паразитировать и на бактериальных клетках (бактериофаги)
• • • Процесс, как правило, завершается формированием зрелых частиц вируса и разрушением плазматической мембраны клетки, после чего вышедшие наружу вирусы получают возможность инфицировать новые хозяйственные клетки Существуют вирусы, паразитирующие в растительных и животных клетках, в том числе в клетках человека, вызывая различные вирусные заболевания Вирусы могут паразитировать и на бактериальных клетках (бактериофаги)
 состоит из одной или нескольких молекул нуклеиновой кислоты") • Зрелая вирусная частица (вирион) состоит из одной или нескольких молекул нуклеиновой кислоты (ДНК или РНК), которая может быть одно- или двухцепочечной, линейной или кольцевой; и белковой оболочки – капсида, который имеет спиральную (палочки) или икосаэдрическую форму
• Зрелая вирусная частица (вирион) состоит из одной или нескольких молекул нуклеиновой кислоты (ДНК или РНК), которая может быть одно- или двухцепочечной, линейной или кольцевой; и белковой оболочки – капсида, который имеет спиральную (палочки) или икосаэдрическую форму
") Синтез белка (трансляция)
Синтез белка (трансляция)
 в цитоплазму к") Матричная теория • • предусматривает: Перенос информации из ядра (от ДНК) в цитоплазму к рибосомам в виде нуклеотидной последовательности м. РНК в процессе транскрипции Активацию аминокислот и перенос активированных аминокислот на полисомы (рибосомы) с помощью транспортных т. РНК Синтез пептидных связей – трансляцию
Матричная теория • • предусматривает: Перенос информации из ядра (от ДНК) в цитоплазму к рибосомам в виде нуклеотидной последовательности м. РНК в процессе транскрипции Активацию аминокислот и перенос активированных аминокислот на полисомы (рибосомы) с помощью транспортных т. РНК Синтез пептидных связей – трансляцию
 Активирование аминокислот
Активирование аминокислот
 Перенос активированных аминокислот на т. РНК
Перенос активированных аминокислот на т. РНК
 Инициация синтеза белка
Инициация синтеза белка
 Элонгация синтеза белка
Элонгация синтеза белка
 Терминация синтеза белка
Терминация синтеза белка