

Biology-3.ppt
- Количество слайдов: 24
 Нуклеиновые кислоты
Нуклеиновые кислоты
 Мономерами НК являются нуклеотиды, состоящие из: 1. азотистого основания 2. углевода пентозы 3. остатка фосфорной кислоты 1 3 2
Мономерами НК являются нуклеотиды, состоящие из: 1. азотистого основания 2. углевода пентозы 3. остатка фосфорной кислоты 1 3 2
 Азотистые основания – состоят из атомов углерода и азота, формирующих циклическую структуру. Различают два класса азотистых оснований:
Азотистые основания – состоят из атомов углерода и азота, формирующих циклическую структуру. Различают два класса азотистых оснований:
 Нуклеотиды обеспечивают аккумуляцию, передачу и использование энергии во всех процессах. Аденозиндифосфат:") Функции нуклеотидов 1) Нуклеотиды обеспечивают аккумуляцию, передачу и использование энергии во всех процессах. Аденозиндифосфат: в результате присоединения еще одного остатка фосфорной кислоты (фосфорилирование) превращается в АТФ. АДФ + Ф ATФ + Н 2 О Для осуществления этой реакции затрачивается 40 к. Дж/моль, следовательно, при разрыве этой связи также выделится столько же энергии.
Функции нуклеотидов 1) Нуклеотиды обеспечивают аккумуляцию, передачу и использование энергии во всех процессах. Аденозиндифосфат: в результате присоединения еще одного остатка фосфорной кислоты (фосфорилирование) превращается в АТФ. АДФ + Ф ATФ + Н 2 О Для осуществления этой реакции затрачивается 40 к. Дж/моль, следовательно, при разрыве этой связи также выделится столько же энергии.
 являются мономерами нуклеиновых кислот, которые связываются между собой за счет формирования ковалентных связей") 2) являются мономерами нуклеиновых кислот, которые связываются между собой за счет формирования ковалентных связей между пентозой первого нуклеотида и остатком фосфорной кислоты второго нуклеотида и т. д.
2) являются мономерами нуклеиновых кислот, которые связываются между собой за счет формирования ковалентных связей между пентозой первого нуклеотида и остатком фосфорной кислоты второго нуклеотида и т. д.
 ДНК – дезоксирибонуклеиновая кислота Под первичной структурой ДНК подразумевают последовательность нуклеотидов в каждой из двух нитей молекулы. ДНК включает четыре азотистых основания: 1) пуриновых – аденин и гуанин и 2) пиримидиновых – цитозин и тимин 1 2
ДНК – дезоксирибонуклеиновая кислота Под первичной структурой ДНК подразумевают последовательность нуклеотидов в каждой из двух нитей молекулы. ДНК включает четыре азотистых основания: 1) пуриновых – аденин и гуанин и 2) пиримидиновых – цитозин и тимин 1 2
 Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного нуклеотида с гидроксилом другого так, что между ними устанавливается фосфодиэфирная связь.
Соединение нуклеотидов в макромолекулу нуклеиновой кислоты происходит путем взаимодействия фосфата одного нуклеотида с гидроксилом другого так, что между ними устанавливается фосфодиэфирная связь.
 Первичная структура. Сборка полинуклеотидной цепи осуществляется при участии фермента полимеразы, который обеспечивает присоединение фосфатной группы следующего нуклеотида к гидроксильной группе, стоящей в положении 3', предыдущего нуклеотида. Благодаря отмеченной специфике, наращивание полинуклеотидной цепи происходит только на одном конце: там, где находится свободный гидроксил в положении 3'. Начало цепи всегда несет фосфатную группу в положении 5'. Это позволяет выделить в ней 5' и 3 '-концы.
Первичная структура. Сборка полинуклеотидной цепи осуществляется при участии фермента полимеразы, который обеспечивает присоединение фосфатной группы следующего нуклеотида к гидроксильной группе, стоящей в положении 3', предыдущего нуклеотида. Благодаря отмеченной специфике, наращивание полинуклеотидной цепи происходит только на одном конце: там, где находится свободный гидроксил в положении 3'. Начало цепи всегда несет фосфатную группу в положении 5'. Это позволяет выделить в ней 5' и 3 '-концы.
 Вторичная структура ДНК возникает за счет того, что две цепи объединяются в единую молекулу водородными связями, возникающими между азотистыми основаниями, входящими в состав нуклеотидов, образующих разные цепи.
Вторичная структура ДНК возникает за счет того, что две цепи объединяются в единую молекулу водородными связями, возникающими между азотистыми основаниями, входящими в состав нуклеотидов, образующих разные цепи.
 Пространственная конфигурация азотистых оснований различна и количество связей между разными азотистыми основаниями неодинаково. Вследствие этого они могут соединяться только попарно (Э. Чаргафф, 1953):
Пространственная конфигурация азотистых оснований различна и количество связей между разными азотистыми основаниями неодинаково. Вследствие этого они могут соединяться только попарно (Э. Чаргафф, 1953):
 две полинуклеотидные цепи ДНК располагаются максимально близко друг") При соблюдении правила комплементарности (А-Т, Г-Ц) две полинуклеотидные цепи ДНК располагаются максимально близко друг к другу и водородные связи обеспечивают наибольшую стабильность системе. При других комбинациях расстояние между полинуклеотидными цепочками возрастает из-за более крупных размеров пуриновых оснований и прочность связей снижается. Другой важной особенностью объединения двух полинуклеотидных цепей в молекуле ДНК является их антипараллельность: 5'-конец одной цепи соединяется с З‘- концом другой, и наоборот. NА. str.
При соблюдении правила комплементарности (А-Т, Г-Ц) две полинуклеотидные цепи ДНК располагаются максимально близко друг к другу и водородные связи обеспечивают наибольшую стабильность системе. При других комбинациях расстояние между полинуклеотидными цепочками возрастает из-за более крупных размеров пуриновых оснований и прочность связей снижается. Другой важной особенностью объединения двух полинуклеотидных цепей в молекуле ДНК является их антипараллельность: 5'-конец одной цепи соединяется с З‘- концом другой, и наоборот. NА. str.
 Модель Д. Уотсона и Ф. Крика Рентгеноструктурный анализ показал, что молекула ДНК, состоящая из двух цепей, образует спираль. Один виток образован 10 парами нуклеотидов и имеет протяженность 3, 4 нм. Диаметр спирали составляет 2 нм, при этом азотистые основания занимают центральное положение, а на поверхности спирали расположены пентоза и фосфатные группы. Вода взаимодействует с пентозофосфатным каркасом и не проникает внутрь, что могло бы привести к ослаблению водородных связей между азотистыми основаниями и дестабилизации вторичной структуры.
Модель Д. Уотсона и Ф. Крика Рентгеноструктурный анализ показал, что молекула ДНК, состоящая из двух цепей, образует спираль. Один виток образован 10 парами нуклеотидов и имеет протяженность 3, 4 нм. Диаметр спирали составляет 2 нм, при этом азотистые основания занимают центральное положение, а на поверхности спирали расположены пентоза и фосфатные группы. Вода взаимодействует с пентозофосфатным каркасом и не проникает внутрь, что могло бы привести к ослаблению водородных связей между азотистыми основаниями и дестабилизации вторичной структуры.
 Третичная структура ядерной ДНК формируется при ее взаимодействии с белками гистонами. Нуклеосомная нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных белков гистонов: Н 2 А, Н 2 В, НЗ, Н 4. Они образуют, напоминающие по форме шайбу белковые тела — коры, Молекула ДНК комплексируется с белковыми корами, спирально накручиваясь на них (почти 2 полных оборота или 146 пар нуклеотидов). Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают, в среднем, 60 п. н. в зависимости от типа клетки. Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому. Вдоль нуклеосомной нити, напоминающей цепочку бус, имеются области ДНК, свободные от белковых тел. Эти области играют важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками.
Третичная структура ядерной ДНК формируется при ее взаимодействии с белками гистонами. Нуклеосомная нить. Этот уровень организации хроматина обеспечивается четырьмя видами нуклеосомных белков гистонов: Н 2 А, Н 2 В, НЗ, Н 4. Они образуют, напоминающие по форме шайбу белковые тела — коры, Молекула ДНК комплексируется с белковыми корами, спирально накручиваясь на них (почти 2 полных оборота или 146 пар нуклеотидов). Свободные от контакта с белковыми телами участки ДНК называют связующими или линкерными. Они включают, в среднем, 60 п. н. в зависимости от типа клетки. Отрезок молекулы ДНК длиной около 200 п. н. вместе с белковым кором составляет нуклеосому. Вдоль нуклеосомной нити, напоминающей цепочку бус, имеются области ДНК, свободные от белковых тел. Эти области играют важную роль в дальнейшей упаковке хроматина, так как содержат нуклеотидные последовательности, специфически узнаваемые различными негистоновыми белками.
 Хроматиновая фибрилла. Дальнейшая компактизация нуклеосомной нити обеспечивается гистоном Н 1, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется более компактная структура, построенная по типу соленоида, имеющая диаметр 20 -30 нм.
Хроматиновая фибрилла. Дальнейшая компактизация нуклеосомной нити обеспечивается гистоном Н 1, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется более компактная структура, построенная по типу соленоида, имеющая диаметр 20 -30 нм.
 Интерфазная хромонема. Следующий уровень структурной организации ДНК обусловлен укладкой хроматиновой фибриллы в петли. В их образовании, по-видимому, принимают участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности во вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы. Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки хроматиновая фибрилла диаметром 20— 30 нм преобразуется в структуру диаметром 100— 200 нм, называемую интерфазной хромонемой. Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяющие соседние петли с одинаковой организацией
Интерфазная хромонема. Следующий уровень структурной организации ДНК обусловлен укладкой хроматиновой фибриллы в петли. В их образовании, по-видимому, принимают участие негистоновые белки, которые способны узнавать специфические нуклеотидные последовательности во вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы. Участок ДНК, соответствующий одной петле, содержит от 20 000 до 80 000 п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки хроматиновая фибрилла диаметром 20— 30 нм преобразуется в структуру диаметром 100— 200 нм, называемую интерфазной хромонемой. Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяющие соседние петли с одинаковой организацией
 хроматина. Отдельные хромосомы") Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией (суперспирализацией) хроматина. Отдельные хромосомы становятся хорошо различимы. Суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза. A. h.
Метафазная хромосома. Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией (суперспирализацией) хроматина. Отдельные хромосомы становятся хорошо различимы. Суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза. A. h.
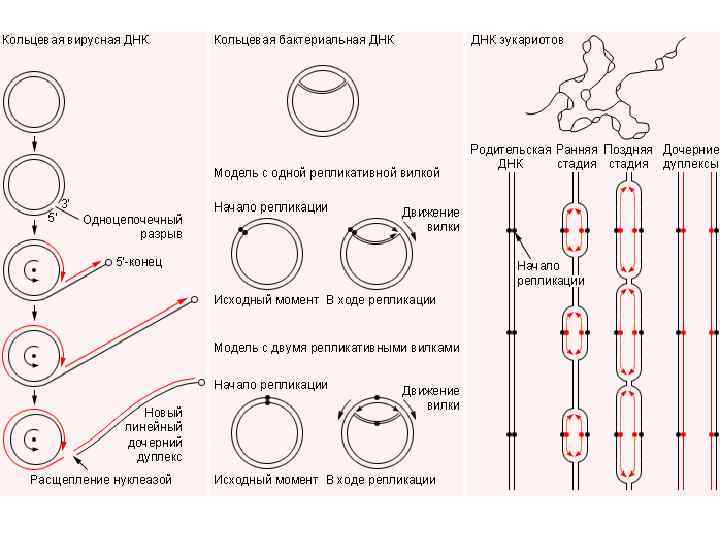
 У прокариот основная часть генетического материала заключена в единственной кольцевой молекуле ДНК длиной около 1 мм (E. coli), причем у них отсутствуют гистоны, обеспечивающие нуклеосомную организацию хроматина. Однако кольцевой молекуле ДНК прокариот также присуща укладка в виде петель. ДНК прокариот комплексируется с некоторыми негистоновыми белками, образуя нуклеоид, который гораздо менее устойчив по сравнению с хроматином эукариот.
У прокариот основная часть генетического материала заключена в единственной кольцевой молекуле ДНК длиной около 1 мм (E. coli), причем у них отсутствуют гистоны, обеспечивающие нуклеосомную организацию хроматина. Однако кольцевой молекуле ДНК прокариот также присуща укладка в виде петель. ДНК прокариот комплексируется с некоторыми негистоновыми белками, образуя нуклеоид, который гораздо менее устойчив по сравнению с хроматином эукариот.


 Третичной структурой называют распределение в пространстве всех атомов белковой молекулы. При этом не учитывают взаимодействие этой глобулы с соседними глобулами или субъединицами. Третичная структура – основа функционирования белка, которая требует точной пространственной организации больших ансамблей, построенных из множества аминокислотных остатков и их боковых групп. Такие ансамбли формируют активные центры ферментов, зоны связывания других молекул и т. д. , поэтому нарушение третичной структуры белка (денатурация) приводит к утрате ими способности функционировать. Первичная структура Вторичная структура Третичная структура
Третичной структурой называют распределение в пространстве всех атомов белковой молекулы. При этом не учитывают взаимодействие этой глобулы с соседними глобулами или субъединицами. Третичная структура – основа функционирования белка, которая требует точной пространственной организации больших ансамблей, построенных из множества аминокислотных остатков и их боковых групп. Такие ансамбли формируют активные центры ферментов, зоны связывания других молекул и т. д. , поэтому нарушение третичной структуры белка (денатурация) приводит к утрате ими способности функционировать. Первичная структура Вторичная структура Третичная структура
 сложные белки b) Четвертичная структура. Гемоглобин") a) сложные белки b) Четвертичная структура. Гемоглобин
a) сложные белки b) Четвертичная структура. Гемоглобин
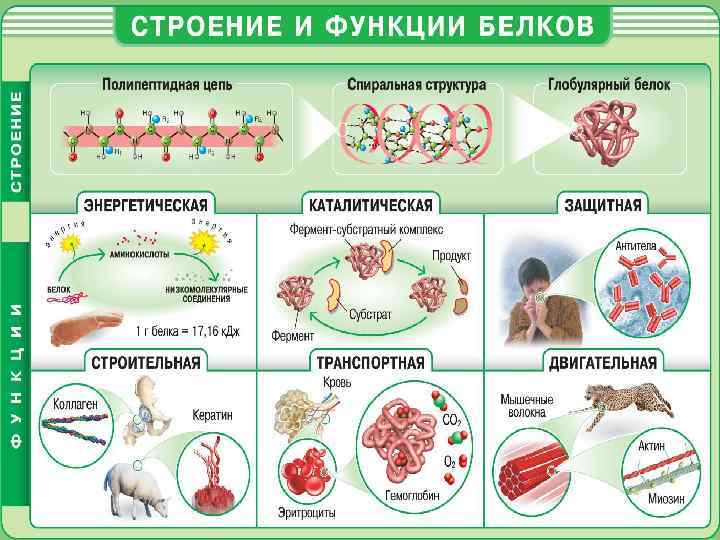
 Можно добавить материал на 15 мин
Можно добавить материал на 15 мин