

FFM_lecture05_2013.ppt
- Количество слайдов: 50
 НОРМАЛЬНАЯ ФИЗИОЛОГИЯ ЛЕКЦИЯ 5 • Электрофизиология межклеточных взаимодействий • Электрические синапсы • Химические синапсы Гайдуков Александр Евгеньевич ФФМ МГУ 2013
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ ЛЕКЦИЯ 5 • Электрофизиология межклеточных взаимодействий • Электрические синапсы • Химические синапсы Гайдуков Александр Евгеньевич ФФМ МГУ 2013
 Камилло Гольджи") История изучения межклеточных контактов Данные микроскопических исследований (конец XIX- начало XX веков) Камилло Гольджи Теория нервной сети ( «Ретикулярная доктрина» ) Все нервные клетки перетекают одна в другую, структурно НЕРАЗРЫВНЫ! VS. Сантьяго Рамон-и-Кахал Нейронная теория ( «Нейронная доктрина» ) Нейроны – отделены один от другого! Передача электрических сигналов от нейрона к нейрону происходит через очень узкие щели, существующие между отростками нервных клеток!
История изучения межклеточных контактов Данные микроскопических исследований (конец XIX- начало XX веков) Камилло Гольджи Теория нервной сети ( «Ретикулярная доктрина» ) Все нервные клетки перетекают одна в другую, структурно НЕРАЗРЫВНЫ! VS. Сантьяго Рамон-и-Кахал Нейронная теория ( «Нейронная доктрина» ) Нейроны – отделены один от другого! Передача электрических сигналов от нейрона к нейрону происходит через очень узкие щели, существующие между отростками нервных клеток!
 Место контакта между нейронами получило название С И Н А П С 1897 г. - Чарльз Скотт Шеррингтон - «синаптеин» (греч. συνάπτειν — обнимать, обхватывать, пожимать руку ) Задача для электрофизиологии (30 -40 -е годы XX столетия) – объяснить, как именно происходит передача электрического сигнала через узкую синаптическую ЩЕЛЬ между нейронами? Гипотеза пассивного затекания электрического тока Гипотеза химического сигнала, передающего электрический сигнал от одной клетки – к другой
Место контакта между нейронами получило название С И Н А П С 1897 г. - Чарльз Скотт Шеррингтон - «синаптеин» (греч. συνάπτειν — обнимать, обхватывать, пожимать руку ) Задача для электрофизиологии (30 -40 -е годы XX столетия) – объяснить, как именно происходит передача электрического сигнала через узкую синаптическую ЩЕЛЬ между нейронами? Гипотеза пассивного затекания электрического тока Гипотеза химического сигнала, передающего электрический сигнал от одной клетки – к другой
 Bernard Katz, University College London, UK Использовал эквивалентную электрическую схему синаптической связи между клетками для расчетов эффективности и величины пассивно затекающего электрического тока ? ΔVm 1 ΔVm 2 Iвход Сm 1 Rконтакта Сm 2 Rm 2 ток Iвход клетка 1 клетка 2 Как рассчитать сдвиг МП – ΔVm 2 - возникающий во второй клетке при пассивном затекании тока, генерируемого клеткой 1?
Bernard Katz, University College London, UK Использовал эквивалентную электрическую схему синаптической связи между клетками для расчетов эффективности и величины пассивно затекающего электрического тока ? ΔVm 1 ΔVm 2 Iвход Сm 1 Rконтакта Сm 2 Rm 2 ток Iвход клетка 1 клетка 2 Как рассчитать сдвиг МП – ΔVm 2 - возникающий во второй клетке при пассивном затекании тока, генерируемого клеткой 1?
 В области нервно-мышечного контакта величина синаптической щели ~100 нм, и если Rконтакта < Rm , тогда ΔVm 2 << ΔVm 1 , Расчеты Б. Катца показали, что в случае нервно-мышечного синапса ΔVm 2 / ΔVm 1 << 0, 01 В нервно-мышечном синапсе ΔVm 1 ΔVm 2 Если в пресинапсе сдвиг Vm - 100 м. В, то на постсинаптической мембране - <<1 м. В !! Нервно-мышечный синапс не может передавать сигнал за счет пассивного затекания тока! – слишком велика потеря сигнала «…Невозможно нагреть пушечное ядро, прикасаясь к нему раскаленной иглой» Бернард Катц «Нерв, мышца, синапс» 1968
В области нервно-мышечного контакта величина синаптической щели ~100 нм, и если Rконтакта < Rm , тогда ΔVm 2 << ΔVm 1 , Расчеты Б. Катца показали, что в случае нервно-мышечного синапса ΔVm 2 / ΔVm 1 << 0, 01 В нервно-мышечном синапсе ΔVm 1 ΔVm 2 Если в пресинапсе сдвиг Vm - 100 м. В, то на постсинаптической мембране - <<1 м. В !! Нервно-мышечный синапс не может передавать сигнал за счет пассивного затекания тока! – слишком велика потеря сигнала «…Невозможно нагреть пушечное ядро, прикасаясь к нему раскаленной иглой» Бернард Катц «Нерв, мышца, синапс» 1968
 Первое обнаружение электрического синапса 1959 г. Гигантский синапс в ЦНС рака Электрическая передача с пре- на постсинаптическое волокно Задействована в реакции избегания рака Сm 1 Сm 2 Rm 1 Rконтакта Ксвязи = Rm 1 / Rm 2+Rконтакта Rm 2 Ксвязи = ΔV 2 / ΔV 1= 0, 6
Первое обнаружение электрического синапса 1959 г. Гигантский синапс в ЦНС рака Электрическая передача с пре- на постсинаптическое волокно Задействована в реакции избегания рака Сm 1 Сm 2 Rm 1 Rконтакта Ксвязи = Rm 1 / Rm 2+Rконтакта Rm 2 Ксвязи = ΔV 2 / ΔV 1= 0, 6
 Структурные требования к электрическому механизму синаптической передачи • Тесное прилегание пре- и постсинаптических мембран • Наличие системы щелевых контактов (gap junction) - быстрый пассивный перенос ионов между клетками (как правило, в обе стороны). В электрическом синапсе сигнал ослабляется! Главное преимущество – высокая скорость передачи Факторы, влияющие на коннексоны р. Н, уровень СО 2, АФК(супероксиды)
Структурные требования к электрическому механизму синаптической передачи • Тесное прилегание пре- и постсинаптических мембран • Наличие системы щелевых контактов (gap junction) - быстрый пассивный перенос ионов между клетками (как правило, в обе стороны). В электрическом синапсе сигнал ослабляется! Главное преимущество – высокая скорость передачи Факторы, влияющие на коннексоны р. Н, уровень СО 2, АФК(супероксиды)
 GAP JUNCTION Строение щелевого контакта
GAP JUNCTION Строение щелевого контакта
 Ксвязи = Rm 1 / Rm 2+Rконтакта ~ 1 2 мм Кардиомиоциты, связанные щелевыми контактами
Ксвязи = Rm 1 / Rm 2+Rконтакта ~ 1 2 мм Кардиомиоциты, связанные щелевыми контактами
 в разных тканях слои сетчатки глиальные клетки") гладкая мускулатура Щелевые контакты (gap junctions) в разных тканях слои сетчатки глиальные клетки
гладкая мускулатура Щелевые контакты (gap junctions) в разных тканях слои сетчатки глиальные клетки
 Основные свойства электрических синапсов • Прилегающие мембраны соединены щелевым контактом. • Ток течет из одной клетки в другую в области щелевых контактов через широкие каналы, образуемые белками - коннексонами. • Сигнал выигрывает в скорости распространения (лимитируется только диффузией), но при этом всегда теряет в амплитуде (усиление сигнала в электрическом синапсе - невозможно) • Сигналы могут передаваться в обоих направлениях (нет полупроводникового эффекта, как в химическом синапсе) – но есть исключения. • Основная функция – синхронизация электрической активности в популяции близко расположенных нейронов. • Не только ионы, но и вещества большего размера, например, АТФ, могут распространяться этим путем. • Менее распространены, чем химические (<1%).
Основные свойства электрических синапсов • Прилегающие мембраны соединены щелевым контактом. • Ток течет из одной клетки в другую в области щелевых контактов через широкие каналы, образуемые белками - коннексонами. • Сигнал выигрывает в скорости распространения (лимитируется только диффузией), но при этом всегда теряет в амплитуде (усиление сигнала в электрическом синапсе - невозможно) • Сигналы могут передаваться в обоих направлениях (нет полупроводникового эффекта, как в химическом синапсе) – но есть исключения. • Основная функция – синхронизация электрической активности в популяции близко расположенных нейронов. • Не только ионы, но и вещества большего размера, например, АТФ, могут распространяться этим путем. • Менее распространены, чем химические (<1%).
 ХИМИЧЕСКИЙ СИНАПС • 1877 Эмиль Дюбуа-Реймон предположил, что Эмиль Дюбуа-Реймон передача возбуждения с нерва на мышцу осуществляется при помощи освобождения химического вещества. • 1921 Отто Лёви показывает выделение химического вещества из окончаний блуждающего нерва (“Vagusstoff”) • 1936 Генри Дейл демонстрирует, что Генри Дейл ацетилхолин освобождается из двигательных нервных окончаний. Структурные особенности: • пресинаптическое окончание (нервная терминаль) – синтез и секреция медиатора • синаптическая щель (20 -100 нм) – диффузия медиатора • постсинаптическая мембрана – постсинаптические рецепторы: 1. Ионотропные (лиганд-управляемые ионные каналы) 2. Метаботропные (запуск метаболических процессов через систему II посредников – усиление сигнала!)
ХИМИЧЕСКИЙ СИНАПС • 1877 Эмиль Дюбуа-Реймон предположил, что Эмиль Дюбуа-Реймон передача возбуждения с нерва на мышцу осуществляется при помощи освобождения химического вещества. • 1921 Отто Лёви показывает выделение химического вещества из окончаний блуждающего нерва (“Vagusstoff”) • 1936 Генри Дейл демонстрирует, что Генри Дейл ацетилхолин освобождается из двигательных нервных окончаний. Структурные особенности: • пресинаптическое окончание (нервная терминаль) – синтез и секреция медиатора • синаптическая щель (20 -100 нм) – диффузия медиатора • постсинаптическая мембрана – постсинаптические рецепторы: 1. Ионотропные (лиганд-управляемые ионные каналы) 2. Метаботропные (запуск метаболических процессов через систему II посредников – усиление сигнала!)
 Медиаторы Требования к молекулярным свойствам медиаторов • Высокая скорость диффузии, а значит - низкий молекулярный вес • Относительная простота и скорость синтеза (небольшое число стадий) • Доступность исходных продуктов и наличие систем поступления их в клетку • Невысокие энергетические затраты на синтез или обратный захват нейроном • Возможность повторного использования самого медиатора или непосредственных продуктов его метаболизма Почти все медиаторы способны как возбуждать, так и тормозить Часть медиаторов могут участвовать как в быстрых, так и медленных процессах Основные медиаторы Ацетилхолин Норадреналин Серотонин (5 -HT) Гистамин Дофамин Глутамат ГАМК Глицин АТФ Аденозин Нейропептиды: опиоиды вещество Р соматостатин нейротензин и многие другие Газы: NO CO H 2 S
Медиаторы Требования к молекулярным свойствам медиаторов • Высокая скорость диффузии, а значит - низкий молекулярный вес • Относительная простота и скорость синтеза (небольшое число стадий) • Доступность исходных продуктов и наличие систем поступления их в клетку • Невысокие энергетические затраты на синтез или обратный захват нейроном • Возможность повторного использования самого медиатора или непосредственных продуктов его метаболизма Почти все медиаторы способны как возбуждать, так и тормозить Часть медиаторов могут участвовать как в быстрых, так и медленных процессах Основные медиаторы Ацетилхолин Норадреналин Серотонин (5 -HT) Гистамин Дофамин Глутамат ГАМК Глицин АТФ Аденозин Нейропептиды: опиоиды вещество Р соматостатин нейротензин и многие другие Газы: NO CO H 2 S
 Сравнительная продолжительность различных событий в синапсах
Сравнительная продолжительность различных событий в синапсах

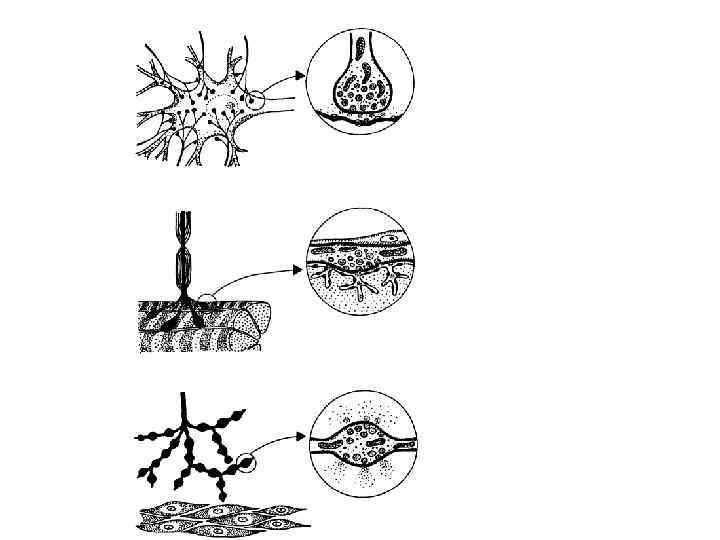
 КВАНТОВАЯ ГИПОТЕЗА ВЫСВОБОЖДЕНИЯ МЕДИАТОРА 1952 -1954 года… Бернард Катц сотоварищи (Дель Кастильо, Фэтт и др. ) детально описывают ПКП и МПКП в нервно-мышечном синапсе. Медиатор освобождается определенными порциями- квантами Нервно-мышечный синапс - классический экспериментальный объект ХХ века для изучения химической синаптической передачи
КВАНТОВАЯ ГИПОТЕЗА ВЫСВОБОЖДЕНИЯ МЕДИАТОРА 1952 -1954 года… Бернард Катц сотоварищи (Дель Кастильо, Фэтт и др. ) детально описывают ПКП и МПКП в нервно-мышечном синапсе. Медиатор освобождается определенными порциями- квантами Нервно-мышечный синапс - классический экспериментальный объект ХХ века для изучения химической синаптической передачи
 Освобождение медиатора контролируется электрическим потенциалом Проблема: «по команде» - много молекул медиатора должно появиться в синаптической щели практически одномоментно… Решение: «упаковка» молекул медиатора в СИНАПТИЧЕСКИЕ ВЕЗИКУЛЫ
Освобождение медиатора контролируется электрическим потенциалом Проблема: «по команде» - много молекул медиатора должно появиться в синаптической щели практически одномоментно… Решение: «упаковка» молекул медиатора в СИНАПТИЧЕСКИЕ ВЕЗИКУЛЫ
 –") Квантовое освобождение медиатора • Медиаторы секретируются мультимолекулярными пакетами (» 7 -10 тыс. молекул) – квантами • Квант соответствует содержимому одной (1) синаптической везикулы • МПКП есть результат постсинаптического действия одного кванта медиатора МПКП • Возникают спонтанно, вне зависимости от стимуляции нерва • Имеют тот же временной ход, что и вызванные ответы • Амплитуда МПКП зависит от состояния рецепторов и концентрации медиатора в везикуле • Частота МПКП зависит от состояния пресинаптического окончания ПКП • Вызванные стимуляцией двигательного нерва потенциалы концевой пластинки (ПКП) – суммарный ответ на синхронное освобождение целого числа квантов: 1, 2, 3…n (>100 ) • Воздействия на нервную терминаль влияют на число этих квантов, но не на амплитуду ответа на один квант
Квантовое освобождение медиатора • Медиаторы секретируются мультимолекулярными пакетами (» 7 -10 тыс. молекул) – квантами • Квант соответствует содержимому одной (1) синаптической везикулы • МПКП есть результат постсинаптического действия одного кванта медиатора МПКП • Возникают спонтанно, вне зависимости от стимуляции нерва • Имеют тот же временной ход, что и вызванные ответы • Амплитуда МПКП зависит от состояния рецепторов и концентрации медиатора в везикуле • Частота МПКП зависит от состояния пресинаптического окончания ПКП • Вызванные стимуляцией двигательного нерва потенциалы концевой пластинки (ПКП) – суммарный ответ на синхронное освобождение целого числа квантов: 1, 2, 3…n (>100 ) • Воздействия на нервную терминаль влияют на число этих квантов, но не на амплитуду ответа на один квант
 распределению Гистограмма") Анализ многоквантовой природы ПКП Гистограмма амплитудных распределений МПКП – соответствует нормальному (Гауссовому) распределению Гистограмма амплитудных распределений ПКП Регистрация ПКП в ответ на одиночные раздражения нерва в условиях сниженного входа кальция в терминаль ([Mg]нар - 5 -7 м. М) Полимодальное скошенное распределение соответствует случаю Пуассоновского распределения с низкой вероятностью независимых событий
Анализ многоквантовой природы ПКП Гистограмма амплитудных распределений МПКП – соответствует нормальному (Гауссовому) распределению Гистограмма амплитудных распределений ПКП Регистрация ПКП в ответ на одиночные раздражения нерва в условиях сниженного входа кальция в терминаль ([Mg]нар - 5 -7 м. М) Полимодальное скошенное распределение соответствует случаю Пуассоновского распределения с низкой вероятностью независимых событий
 Квантовая секреция медиатора – Са 2+-зависимый процесс 1. Повышение уровня Са 2+ в аксоплазме от 10 -8 М (покой) до 10 -4 М (активация) вызывает массивное освобождение квантов медиатора (экзоцитоз) 2. Основной путь входа Са 2+ в терминаль – потенциал-зависимые Са 2+ каналы в пресинаптической мембране, открываемые при деполяризации. Блокирование этих каналов - устраняет эффект стимуляции Calyx of Held
Квантовая секреция медиатора – Са 2+-зависимый процесс 1. Повышение уровня Са 2+ в аксоплазме от 10 -8 М (покой) до 10 -4 М (активация) вызывает массивное освобождение квантов медиатора (экзоцитоз) 2. Основной путь входа Са 2+ в терминаль – потенциал-зависимые Са 2+ каналы в пресинаптической мембране, открываемые при деполяризации. Блокирование этих каналов - устраняет эффект стимуляции Calyx of Held
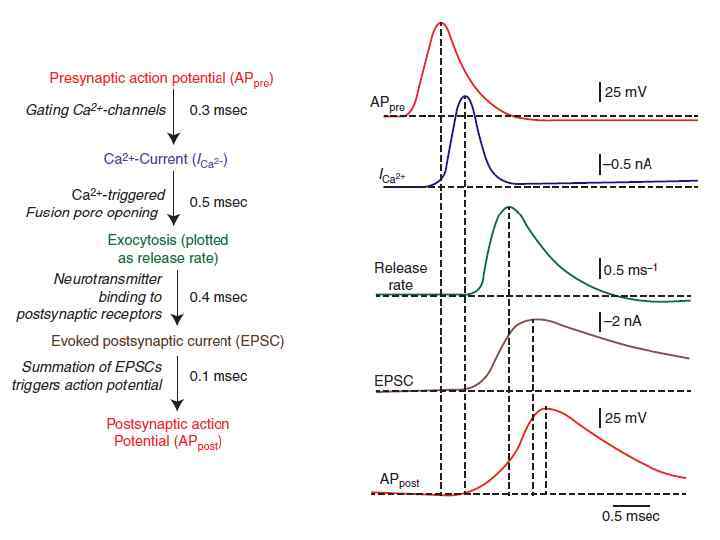
 Квантовая секреция медиатора – Са 2+-зависимый процесс Основная последовательность явлений: • Приход потенциала действия (ПД) в терминаль • Активация (открытие) Са 2+- каналов • Быстрый локальный рост уровня Са 2+ • Вызванное Са 2+ повышение вероятности экзоцитоза медиатора Эти процессы занимают 1 - 2 мс, что и соответствует синаптической задержке между стимулом (пресинаптическим ПД) и постсинаптическим ответом
Квантовая секреция медиатора – Са 2+-зависимый процесс Основная последовательность явлений: • Приход потенциала действия (ПД) в терминаль • Активация (открытие) Са 2+- каналов • Быстрый локальный рост уровня Са 2+ • Вызванное Са 2+ повышение вероятности экзоцитоза медиатора Эти процессы занимают 1 - 2 мс, что и соответствует синаптической задержке между стимулом (пресинаптическим ПД) и постсинаптическим ответом
 Потенциал-управляемые Са 2+-каналы Особенности: • Наличие потенциал-зависимой и кальций-зависимой инактивации • Время открытого состояния канала зависит от степени его фосфорилирования! – работой этих каналов можно управлять (модуляция синаптической передачи)
Потенциал-управляемые Са 2+-каналы Особенности: • Наличие потенциал-зависимой и кальций-зависимой инактивации • Время открытого состояния канала зависит от степени его фосфорилирования! – работой этих каналов можно управлять (модуляция синаптической передачи)
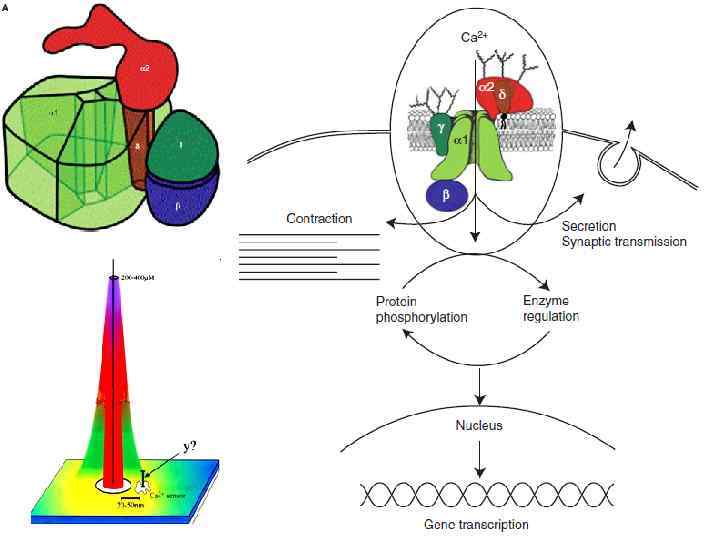
 Классификация потенциал-зависимых Са 2+-каналов
Классификация потенциал-зависимых Са 2+-каналов
 Изменения концентрации Са 2+ в аксоплазме терминали Домены Са 2+ вблизи внутренней поверхности пресинаптической мембраны Значение взаимного расположения Са 2+ каналов и участков экзоцитоза Са 2+ входит в аксоплазму: локальный подъем концентрации от 0. 1 m. M до 600 -800 m. M Через 1 -1. 5 мс после закрытия Са 2+каналов: остается < 10% вошедшего Са 2+, затем уровень Са 2+ быстро снижается внутриклеточными буферными системами и Са 2+насосами до уровня покоя
Изменения концентрации Са 2+ в аксоплазме терминали Домены Са 2+ вблизи внутренней поверхности пресинаптической мембраны Значение взаимного расположения Са 2+ каналов и участков экзоцитоза Са 2+ входит в аксоплазму: локальный подъем концентрации от 0. 1 m. M до 600 -800 m. M Через 1 -1. 5 мс после закрытия Са 2+каналов: остается < 10% вошедшего Са 2+, затем уровень Са 2+ быстро снижается внутриклеточными буферными системами и Са 2+насосами до уровня покоя
… АКТИВНЫЕ ЗОНЫ") Реконструкция (метод замораживанияскалывания)… АКТИВНЫЕ ЗОНЫ
Реконструкция (метод замораживанияскалывания)… АКТИВНЫЕ ЗОНЫ
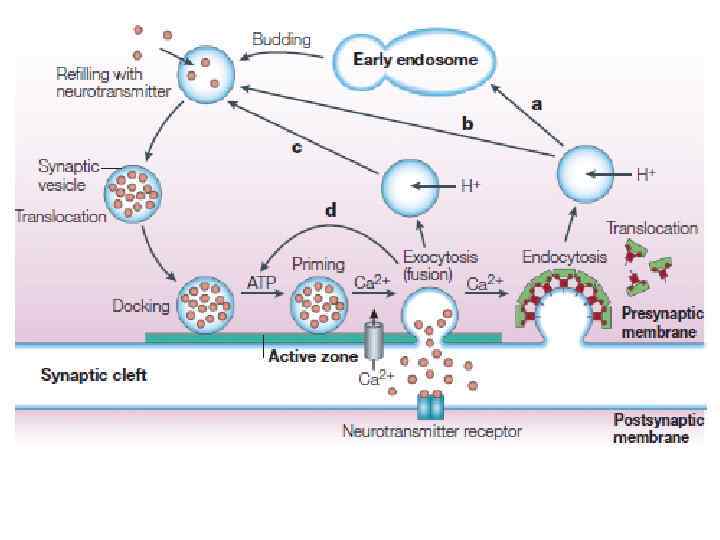
 ПУТИ СЕКРЕЦИИ В НЕЙРОНАХ
ПУТИ СЕКРЕЦИИ В НЕЙРОНАХ
 «Жизненный цикл» синаптических везикул Заполнение везикулы медиатором Вторично-активный транспорт У каждого медиатора свой транспортер, но принципы их работы - общие • Докинг – подход и начальное заякоревание везикулы (из RP) • Прайминг – финальное причаливание везикулы, образование SNARE-комплекса (RRP) • Вход кальция – взаимодействие его с синаптотагмином (везикулярный Са 2+-сенсор) • Экзоцитоз SNARE-Soluble N-ethylmaleimide-sensitive factor Attachment protein Receptor
«Жизненный цикл» синаптических везикул Заполнение везикулы медиатором Вторично-активный транспорт У каждого медиатора свой транспортер, но принципы их работы - общие • Докинг – подход и начальное заякоревание везикулы (из RP) • Прайминг – финальное причаливание везикулы, образование SNARE-комплекса (RRP) • Вход кальция – взаимодействие его с синаптотагмином (везикулярный Са 2+-сенсор) • Экзоцитоз SNARE-Soluble N-ethylmaleimide-sensitive factor Attachment protein Receptor
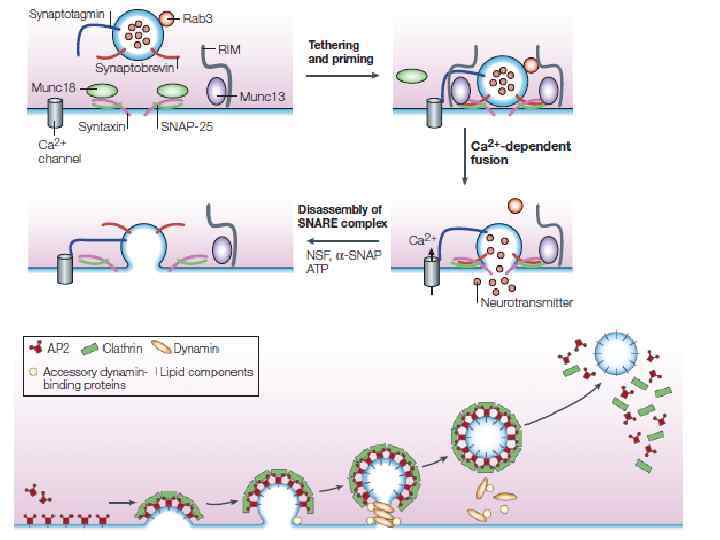
 Взаимодействие белков синаптических везикул и пресинаптической мембраны в активной зоне
Взаимодействие белков синаптических везикул и пресинаптической мембраны в активной зоне
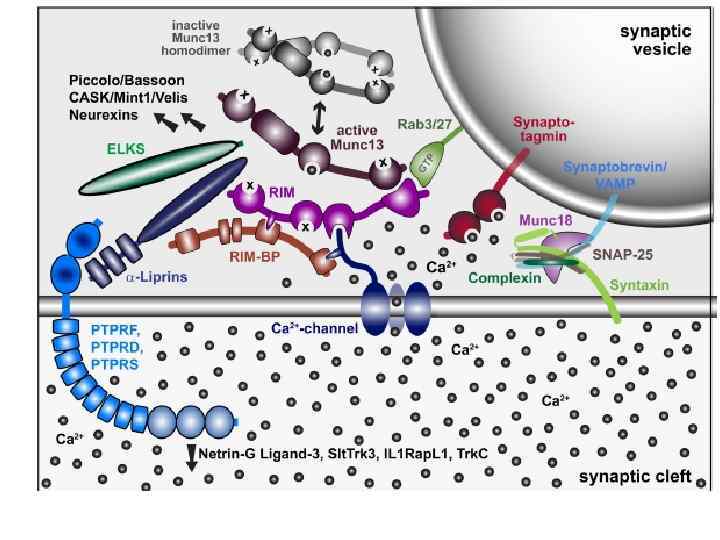
 Взаимодействие белков синаптических везикул и мембраны активной зоны при экзоцитозе
Взаимодействие белков синаптических везикул и мембраны активной зоны при экзоцитозе
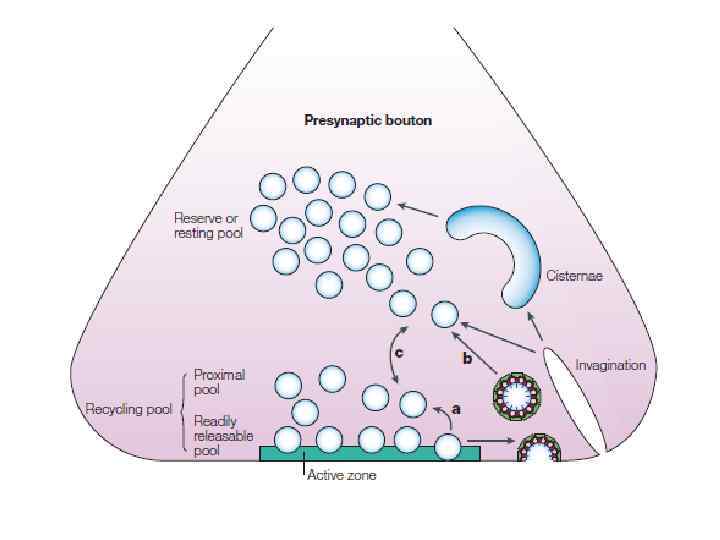
 ВАРИАНТЫ ЭКЗОЦИТОЗА
ВАРИАНТЫ ЭКЗОЦИТОЗА
 ПОСЛЕ ЭКЗОЦИТОЗА Медиаторы попадают в синаптическую щель… Синаптическая щель – не просто пространство между пре- и постсинаптическими мембранами, где идет пассивная диффузия медиатора • Молекулы клеточной адгезии • Внеклеточный матрикс (из структурных белков, секретированных клетками) В нервно-мышечном синапсе формируется базальная мембрана (сеть, на которой заякорены ферменты (напр. АХЭ) и другие сигнальные молекулы)
ПОСЛЕ ЭКЗОЦИТОЗА Медиаторы попадают в синаптическую щель… Синаптическая щель – не просто пространство между пре- и постсинаптическими мембранами, где идет пассивная диффузия медиатора • Молекулы клеточной адгезии • Внеклеточный матрикс (из структурных белков, секретированных клетками) В нервно-мышечном синапсе формируется базальная мембрана (сеть, на которой заякорены ферменты (напр. АХЭ) и другие сигнальные молекулы)
 Механизмы удаления медиатора из зоны синапса Разрушение в синаптической щели с последующим захватом продуктов гидролиза в пресинаптическое окончание Метаболизм Пример: АХЭ в нервно-мышечном синапсе ацетилхолина в холинергическом синапсе
Механизмы удаления медиатора из зоны синапса Разрушение в синаптической щели с последующим захватом продуктов гидролиза в пресинаптическое окончание Метаболизм Пример: АХЭ в нервно-мышечном синапсе ацетилхолина в холинергическом синапсе
 Механизмы удаления медиатора из зоны синапса Захват неразрушенного медиатора с помощью специфических транспортеров в пресинаптическое окончание Пример: синапсы с биогенными аминами в качестве медиаторов
Механизмы удаления медиатора из зоны синапса Захват неразрушенного медиатора с помощью специфических транспортеров в пресинаптическое окончание Пример: синапсы с биогенными аминами в качестве медиаторов
 Механизмы удаления медиатора из зоны синапса Захват неразрушенного медиатора с помощью специфических транспортеров не только в пресинаптическое окончание, но и в соседние глиальные клетки Пример: глутаматные синапсы и синапсы с тормозными медиаторами (ГАМК или глицин)
Механизмы удаления медиатора из зоны синапса Захват неразрушенного медиатора с помощью специфических транспортеров не только в пресинаптическое окончание, но и в соседние глиальные клетки Пример: глутаматные синапсы и синапсы с тормозными медиаторами (ГАМК или глицин)
 Медиаторы диффундируют к постсинаптической мембране… …там их ждут постсинаптические рецепторы
Медиаторы диффундируют к постсинаптической мембране… …там их ждут постсинаптические рецепторы
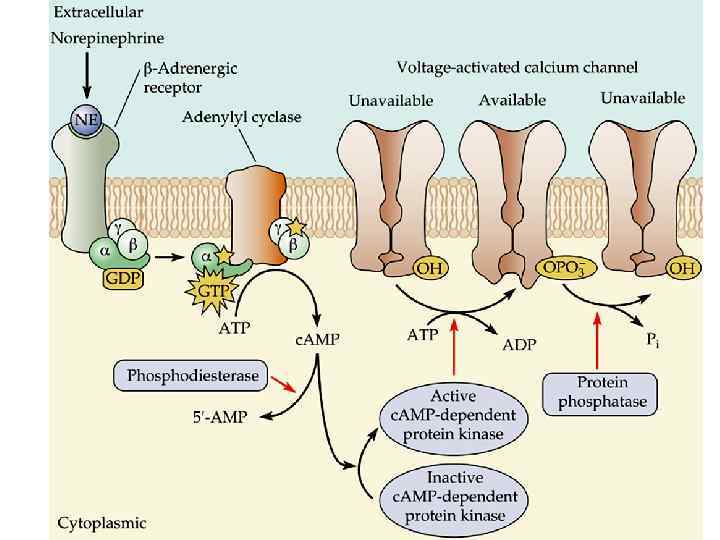
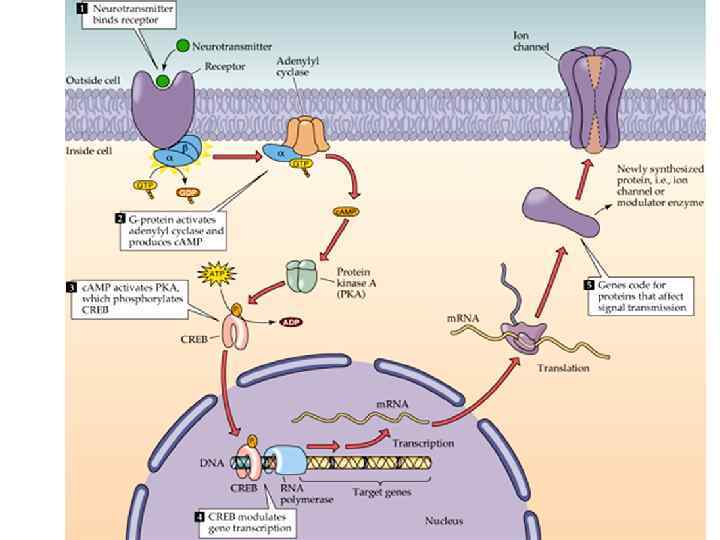
 Быстрый путь, но низкий коэффициент") 2 основных класса постсинаптических рецепторов Ионотропные (лиганд-зависимые ионные каналы) Быстрый путь, но низкий коэффициент усиления! Метаботропные (комплекс с G-белком) Более длинный путь до мишени, но высокий коэффициент усиления!
2 основных класса постсинаптических рецепторов Ионотропные (лиганд-зависимые ионные каналы) Быстрый путь, но низкий коэффициент усиления! Метаботропные (комплекс с G-белком) Более длинный путь до мишени, но высокий коэффициент усиления!
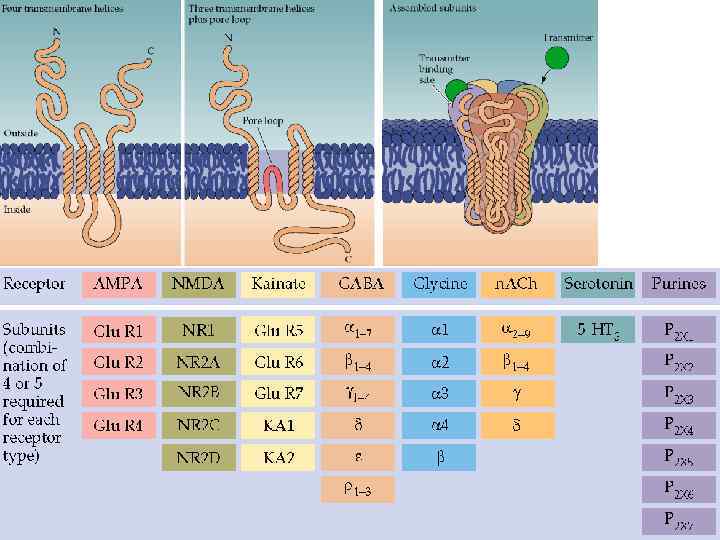
 СУПЕРСЕМЕЙСТВО МЕТАБОТРОПНЫХ РЕЦЕПТОРОВ
СУПЕРСЕМЕЙСТВО МЕТАБОТРОПНЫХ РЕЦЕПТОРОВ
 ТОКИ ЧЕРЕЗ КАНАЛЫ ИОНОТРОПНЫХ РЕЦЕПТОРОВ Na+ Ca 2+ каналы Cl – каналы Возбуждающий постсинаптический ток = ВПСТ K+ каналы Сl – каналы Тормозный постсинаптический ток = ТПСТ
ТОКИ ЧЕРЕЗ КАНАЛЫ ИОНОТРОПНЫХ РЕЦЕПТОРОВ Na+ Ca 2+ каналы Cl – каналы Возбуждающий постсинаптический ток = ВПСТ K+ каналы Сl – каналы Тормозный постсинаптический ток = ТПСТ
 Спасибо за внимание… Вопросы? ? ?
Спасибо за внимание… Вопросы? ? ?