

FFM_lecture04_2013.ppt
- Количество слайдов: 34
 НОРМАЛЬНАЯ ФИЗИОЛОГИЯ ЛЕКЦИЯ 4 • Потенциал действия • история вопроса • современные представления Гайдуков Александр Евгеньевич ФФМ МГУ 2013
НОРМАЛЬНАЯ ФИЗИОЛОГИЯ ЛЕКЦИЯ 4 • Потенциал действия • история вопроса • современные представления Гайдуков Александр Евгеньевич ФФМ МГУ 2013
") ПОТЕНЦИАЛ ДЕЙСТВИЯ Универсальный высокоамплитудный сигнал, быстро распространяющийся по мембране возбудимой клетки (обеспечивает передачу информации) Быстрое колебание мембранного потенциала (в ответ на раздражение) Изменение знака заряда на мембране Появление (и исчезновение) трансмембранных ионных токов Открытие (и закрытие) потенциал-активируемых ионных каналов
ПОТЕНЦИАЛ ДЕЙСТВИЯ Универсальный высокоамплитудный сигнал, быстро распространяющийся по мембране возбудимой клетки (обеспечивает передачу информации) Быстрое колебание мембранного потенциала (в ответ на раздражение) Изменение знака заряда на мембране Появление (и исчезновение) трансмембранных ионных токов Открытие (и закрытие) потенциал-активируемых ионных каналов
 ЭЛЕКТРОТОН Vm/Im = const
ЭЛЕКТРОТОН Vm/Im = const
 0 Поведение возбудимой клетки при разных условиях электрического раздражения Закон") МП 0 (м. В) 0 Поведение возбудимой клетки при разных условиях электрического раздражения Закон Ома V=IR • Сначала, при слабых толчках тока (1 -2) Мембрана ведет себя как пассивное омическое сопротивление • Увеличение силы поляризующего тока (3 -4) Деполяризующий сдвиг МП ~ нелинеен! • Еще увеличение силы тока (5 -7) Деполяризующий сдвиг МП переходит в регенеративно нарастающую кратковременную деполяризацию (ПД) -70 Последующее увеличением силы тока величина ПД заметно не меняется, а укорачивается время достижения максимума Деполяризующий ток Линейный сдвиг МП – «физический электротон» Нелинейный сдвиг МП – Гиперполяризующий «физиологический электротон» ток
МП 0 (м. В) 0 Поведение возбудимой клетки при разных условиях электрического раздражения Закон Ома V=IR • Сначала, при слабых толчках тока (1 -2) Мембрана ведет себя как пассивное омическое сопротивление • Увеличение силы поляризующего тока (3 -4) Деполяризующий сдвиг МП ~ нелинеен! • Еще увеличение силы тока (5 -7) Деполяризующий сдвиг МП переходит в регенеративно нарастающую кратковременную деполяризацию (ПД) -70 Последующее увеличением силы тока величина ПД заметно не меняется, а укорачивается время достижения максимума Деполяризующий ток Линейный сдвиг МП – «физический электротон» Нелинейный сдвиг МП – Гиперполяризующий «физиологический электротон» ток
 Вот он какой….
Вот он какой….
 Немного истории изучения природы потенциала действия… • 1848 - Emil du Bois-Reymond - открывает ПД • 1850 - Hermann von Helmholtz – впервые измеряет скорость проведения ПД в нерве • 1902 -1912 - Julius Bernstein – гипотеза: ПД – результат изменения ионной проницаемости мембраны • 1907 - Louis Lapicque – предполагает наличие порога генерации ПД Далее - начало работ на гигантском аксоне кальмара
Немного истории изучения природы потенциала действия… • 1848 - Emil du Bois-Reymond - открывает ПД • 1850 - Hermann von Helmholtz – впервые измеряет скорость проведения ПД в нерве • 1902 -1912 - Julius Bernstein – гипотеза: ПД – результат изменения ионной проницаемости мембраны • 1907 - Louis Lapicque – предполагает наличие порога генерации ПД Далее - начало работ на гигантском аксоне кальмара
 Продолжение истории изучения природы потенциала действия… • 1939 - Ken Cole & Howard Curtis – подтверждают гипотезу Бернштейна – проводимость мембраны увеличивается (а сопротивление – падает!) при развитии ПД • 1949 - Alan Hodgkin & Bernard Katz – показали ключевую роль ионов Na при развитии ПД Изменение содержания Na снаружи Блокирование K-проводимости Блокирование Na-проводимости Надо было изучить токи, текущие через мембрану, но… Проблема: протекание тока через мембрану изменение МП за счет перераспределения зарядов, а изменение МП величина протекающих токов ? ? ? Что делать… …и в 1952 году…
Продолжение истории изучения природы потенциала действия… • 1939 - Ken Cole & Howard Curtis – подтверждают гипотезу Бернштейна – проводимость мембраны увеличивается (а сопротивление – падает!) при развитии ПД • 1949 - Alan Hodgkin & Bernard Katz – показали ключевую роль ионов Na при развитии ПД Изменение содержания Na снаружи Блокирование K-проводимости Блокирование Na-проводимости Надо было изучить токи, текущие через мембрану, но… Проблема: протекание тока через мембрану изменение МП за счет перераспределения зарядов, а изменение МП величина протекающих токов ? ? ? Что делать… …и в 1952 году…
 Кульминация истории изучения природы потенциала действия!!! • 1952 – Hodgkin, Katz & Andrew Huxley – 5 статей Метод фиксации потенциала (“voltage-clamp”)
Кульминация истории изучения природы потенциала действия!!! • 1952 – Hodgkin, Katz & Andrew Huxley – 5 статей Метод фиксации потенциала (“voltage-clamp”)
 “If I have seen further, it’s by standing on the shoulders of giants” RIP Huxley († 30. 05. 2012)
“If I have seen further, it’s by standing on the shoulders of giants” RIP Huxley († 30. 05. 2012)
 VOLTAGE-CLAMP Трансмембранные токи при сдвигах потенциала
VOLTAGE-CLAMP Трансмембранные токи при сдвигах потенциала
 VOLTAGE-CLAMP Разделение токов при помощи блокаторов соответствующих каналов Na+-природа «раннего» входящего тока Потенциалозависимость «раннего» и «позднего» токов
VOLTAGE-CLAMP Разделение токов при помощи блокаторов соответствующих каналов Na+-природа «раннего» входящего тока Потенциалозависимость «раннего» и «позднего» токов
 С помощью специальных избирательных блокаторов натриевого и калиевого тока удалось разделить INa и IK и исследовать раздельно Суммарный ток Калиевые токи Суммарный ток Натриевые токи ВЫВОДЫ: • Входящий ток – ионы Na, выходящий ток – ионы K • Разная динамика токов: Na-ток – быстро нарастает и заканчивается (есть быстрая инактивация Na-каналов); K-ток – медленно нарастает и не изменяется в процессе деполяризации (у K-каналов нет инактивации)
С помощью специальных избирательных блокаторов натриевого и калиевого тока удалось разделить INa и IK и исследовать раздельно Суммарный ток Калиевые токи Суммарный ток Натриевые токи ВЫВОДЫ: • Входящий ток – ионы Na, выходящий ток – ионы K • Разная динамика токов: Na-ток – быстро нарастает и заканчивается (есть быстрая инактивация Na-каналов); K-ток – медленно нарастает и не изменяется в процессе деполяризации (у K-каналов нет инактивации)
 МАТЕМАТИЧЕСКАЯ МОДЕЛЬ ПОТЕНЦИАЛА ДЕЙСТВИЯ Перевод кривых токов INa и IK в кривые проводимостей g. K и g. Na IK = g. K(VM – EK) INa = g. Na(VM - ENa) где VM – мембранный потенциал, ЕNa – равновесный потенциал по Na+. где VM – мембранный потенциал, ЕK – равновесный потенциал по K+. Кривые зависимости проводимости от Vм и t g. Na f(VM; t) g. K f(VM; t) Задача: описать математически g. Na и g. K как функции мембранного потенциала (VM) и времени (t). g. Na f(VM) f(t) g. K f(VM) f(t)
МАТЕМАТИЧЕСКАЯ МОДЕЛЬ ПОТЕНЦИАЛА ДЕЙСТВИЯ Перевод кривых токов INa и IK в кривые проводимостей g. K и g. Na IK = g. K(VM – EK) INa = g. Na(VM - ENa) где VM – мембранный потенциал, ЕNa – равновесный потенциал по Na+. где VM – мембранный потенциал, ЕK – равновесный потенциал по K+. Кривые зависимости проводимости от Vм и t g. Na f(VM; t) g. K f(VM; t) Задача: описать математически g. Na и g. K как функции мембранного потенциала (VM) и времени (t). g. Na f(VM) f(t) g. K f(VM) f(t)
 Для калиевой проводимости максимальные проводимости g. K= g. K·n 4 n - безразмерный коэффициент Для натриевой проводимости g. Na= g. Na m 3·h m и h – безразмерные коэффициенты 0 n 1 0 m 1 m f(VM; t) n f(VM; t) 0 h 1 h f(VM; t)
Для калиевой проводимости максимальные проводимости g. K= g. K·n 4 n - безразмерный коэффициент Для натриевой проводимости g. Na= g. Na m 3·h m и h – безразмерные коэффициенты 0 n 1 0 m 1 m f(VM; t) n f(VM; t) 0 h 1 h f(VM; t)
 Система дифференциальных уравнений Ходжкина и Хаксли Изменения тока (с учетом наличия у мембраны ёмкости - Cm) ток утечки ток Na+ ток K+ Для Na+-тока Для K+-тока Константы скорости перехода каналов (их значения - зависят от МП): α – в открытое состояние β– в закрытое состояние Значения коэффициентов m, h и n – тоже зависят от МП!!!
Система дифференциальных уравнений Ходжкина и Хаксли Изменения тока (с учетом наличия у мембраны ёмкости - Cm) ток утечки ток Na+ ток K+ Для Na+-тока Для K+-тока Константы скорости перехода каналов (их значения - зависят от МП): α – в открытое состояние β– в закрытое состояние Значения коэффициентов m, h и n – тоже зависят от МП!!!
 Гипотеза: g K= g K· n 4 Калиевый канал обладает четырьмя заряженными ДИПОЛЬНЫМИ группами – сенсорами напряжения При деполяризации мембраны может происходить СМЕЩЕНИЕ всех четырех группировок. Это приводит к открыванию канала n 1 – вероятность смещения одной из четырех группировок n 2 – вероятность смещения двух из четырех группировок n 3 – вероятность смещения трех из четырех группировок n 4 – вероятноcть правильного смещения всех четырех диполей сенсор мембранного потенциала
Гипотеза: g K= g K· n 4 Калиевый канал обладает четырьмя заряженными ДИПОЛЬНЫМИ группами – сенсорами напряжения При деполяризации мембраны может происходить СМЕЩЕНИЕ всех четырех группировок. Это приводит к открыванию канала n 1 – вероятность смещения одной из четырех группировок n 2 – вероятность смещения двух из четырех группировок n 3 – вероятность смещения трех из четырех группировок n 4 – вероятноcть правильного смещения всех четырех диполей сенсор мембранного потенциала
 (в открытое и закрытое") Изменения коэффициента n и скоростей перехода n-cтворок αиβ f (Vm) (в открытое и закрытое состояния) n f (t) αиβ f (t) n f (Vm)
Изменения коэффициента n и скоростей перехода n-cтворок αиβ f (Vm) (в открытое и закрытое состояния) n f (t) αиβ f (t) n f (Vm)
 Ток возникает только в ответ на деполяризацию мембраны. Увеличивается линейно по мере деполяризации по формуле IK= g. K (Vм - ЕК равн) Воротные токи калиевого канала Смещение положительных зарядов в составе S 4 -домена приводит к появлению асимметричного «воротного» тока Проводимость одиночного К-канала одиночный К-канал работает в режиме случайных переходов из закрытого состояния - в открытое, по принципу «все или ничего» При увеличении деполяризации растет время пребывания в открытом состоянии
Ток возникает только в ответ на деполяризацию мембраны. Увеличивается линейно по мере деполяризации по формуле IK= g. K (Vм - ЕК равн) Воротные токи калиевого канала Смещение положительных зарядов в составе S 4 -домена приводит к появлению асимметричного «воротного» тока Проводимость одиночного К-канала одиночный К-канал работает в режиме случайных переходов из закрытого состояния - в открытое, по принципу «все или ничего» При увеличении деполяризации растет время пребывания в открытом состоянии
 Гипотеза: Натриевый канал имеет два типа «ворот» : • m-ворота – для активации канала (его открывания) g. Na=g. Na m 3·h • h-ворота – для инактивации канала (его запирания) Оба типа ворот - и m-, и h-ворота – это положительно заряженные группировки или диполи в составе молекулы Na-канала, для смещения которых необходима деполяризация мембраны 0 m 1; m f (VM; t) 0 h 1; h f (VM; t) коэффициент h отражает число каналов с ОТКРЫТОЙ инактивационной створкой
Гипотеза: Натриевый канал имеет два типа «ворот» : • m-ворота – для активации канала (его открывания) g. Na=g. Na m 3·h • h-ворота – для инактивации канала (его запирания) Оба типа ворот - и m-, и h-ворота – это положительно заряженные группировки или диполи в составе молекулы Na-канала, для смещения которых необходима деполяризация мембраны 0 m 1; m f (VM; t) 0 h 1; h f (VM; t) коэффициент h отражает число каналов с ОТКРЫТОЙ инактивационной створкой
 Изменения во времени коэффициентов m и h и их скоростей перехода (α и β ) в ответ на мгновенную деполяризацию мембраны αиβ f (t) Суммарная кривая проводимости g ~ m 3 ·h mиh f (t)
Изменения во времени коэффициентов m и h и их скоростей перехода (α и β ) в ответ на мгновенную деполяризацию мембраны αиβ f (t) Суммарная кривая проводимости g ~ m 3 ·h mиh f (t)
 Изменения коэффициентов m∞ и h∞ в зависимости от уровня фиксации потенциала на мембране Vm коэффициент h отражает число каналов (1 - 100 %) с ОТКРЫТОЙ инактивационной створкой При Vm = ППо (Vr) = -65 м. В коэффициент m ~ равен 0, а коэффициент h = 0, 6
Изменения коэффициентов m∞ и h∞ в зависимости от уровня фиксации потенциала на мембране Vm коэффициент h отражает число каналов (1 - 100 %) с ОТКРЫТОЙ инактивационной створкой При Vm = ППо (Vr) = -65 м. В коэффициент m ~ равен 0, а коэффициент h = 0, 6
 Скорости смещения") Потенциало-зависимость скоростей смещения m и h створок (определяются изменениями α и β) Скорости смещения m – (вверху) и h – створок (внизу) различаются на порядок! (обратите внимание на значения по осям Y !!!) Этим объясняется возможность одновременного срабатывания (смещения) створок в ответ на деполяризацию мембраны. Но движутся они с разной скоростью: • m – быстро открывается • h – медленно закрывается
Потенциало-зависимость скоростей смещения m и h створок (определяются изменениями α и β) Скорости смещения m – (вверху) и h – створок (внизу) различаются на порядок! (обратите внимание на значения по осям Y !!!) Этим объясняется возможность одновременного срабатывания (смещения) створок в ответ на деполяризацию мембраны. Но движутся они с разной скоростью: • m – быстро открывается • h – медленно закрывается
 Экспериментальная проверка математической модели Ходжкина–Хаксли • Выявление внутримембранных «емкостных» токов (смещений фиксированных зарядов внутри мембраны), отражающих работу активационных m-ворот и инактивационных h-ворот. Асимметричные токи смещения Вывод: Деполяризация Гиперполяризация В ответ на деполяризацию возникает асимметричный емкостной «ток смещения» , отражающий работу h-ворот + + • Выявление расположения h-частицы с помощью протеолитического удаления инактивации натриевого тока Na-ток в аксоне Проназа 10 -5 М: • снаружи • изнутри Проназа блокирует инактивацию Na-тока, но только если действует изнутри!!! Вывод: Инактивационная h-частица расположена изнутри - на цитоплазматической поверхности Na-канала
Экспериментальная проверка математической модели Ходжкина–Хаксли • Выявление внутримембранных «емкостных» токов (смещений фиксированных зарядов внутри мембраны), отражающих работу активационных m-ворот и инактивационных h-ворот. Асимметричные токи смещения Вывод: Деполяризация Гиперполяризация В ответ на деполяризацию возникает асимметричный емкостной «ток смещения» , отражающий работу h-ворот + + • Выявление расположения h-частицы с помощью протеолитического удаления инактивации натриевого тока Na-ток в аксоне Проназа 10 -5 М: • снаружи • изнутри Проназа блокирует инактивацию Na-тока, но только если действует изнутри!!! Вывод: Инактивационная h-частица расположена изнутри - на цитоплазматической поверхности Na-канала
 Доменная организация молекулы натриевого канала
Доменная организация молекулы натриевого канала
 Суммарная схема - модель изменений проводимости при GNa и GK при генерации ПД по Ходжкину- Хаксли
Суммарная схема - модель изменений проводимости при GNa и GK при генерации ПД по Ходжкину- Хаксли
 Вольтамперные кривые натриевого и калиевого токов, текущих через мембрану аксона при генерации ПД
Вольтамперные кривые натриевого и калиевого токов, текущих через мембрану аксона при генерации ПД
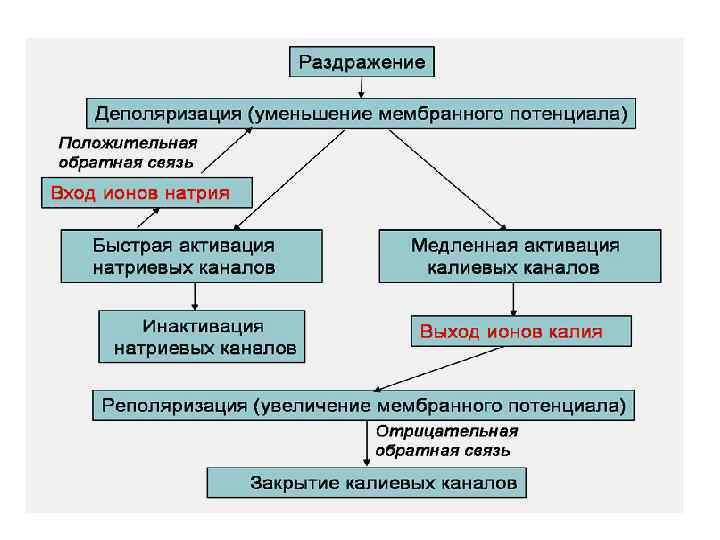
 Свойства ПД в терминах математической модели Ходжкина–Хаксли Аккомодация
Свойства ПД в терминах математической модели Ходжкина–Хаксли Аккомодация
 Следствия математической модели Ходжкина-Хаксли: 1. Описано поведение и свойства натриевых и калиевых каналов, управляемых изменениями электрического поля на мембране 2. Дана научная интерпретация классическим феноменам: • Порог возбуждения – уровень МП, при котором m=n (m 3 =n 4) • Рефрактерность – абсолютная - все Na-каналы инактивированы; относительная - часть Na-каналов инактивирована • Аккомодация – это процесс медленной деполяризации мембраны, при котором dh/dt > dm/dt • Латентный период генерации ПД – это время для смещения активационных m-ворот Na-каналов в открытое состояние
Следствия математической модели Ходжкина-Хаксли: 1. Описано поведение и свойства натриевых и калиевых каналов, управляемых изменениями электрического поля на мембране 2. Дана научная интерпретация классическим феноменам: • Порог возбуждения – уровень МП, при котором m=n (m 3 =n 4) • Рефрактерность – абсолютная - все Na-каналы инактивированы; относительная - часть Na-каналов инактивирована • Аккомодация – это процесс медленной деполяризации мембраны, при котором dh/dt > dm/dt • Латентный период генерации ПД – это время для смещения активационных m-ворот Na-каналов в открытое состояние
 Метод PATCH-CLAMP – регистрация активности одиночных каналов
Метод PATCH-CLAMP – регистрация активности одиночных каналов
 Анализ работы одиночных Na-каналов 1. Канал работает по закону «все или ничего» 2. Проводимость одиночного открытого канала g. Na – величина постоянная, не зависит от Vm 3. Время открытого состояния – короткое и не соответствует продолжительности деполяризации 4. Вероятность пребывания в открытом состоянии (время открытого состояния) возрастает по мере увеличения деполяризации
Анализ работы одиночных Na-каналов 1. Канал работает по закону «все или ничего» 2. Проводимость одиночного открытого канала g. Na – величина постоянная, не зависит от Vm 3. Время открытого состояния – короткое и не соответствует продолжительности деполяризации 4. Вероятность пребывания в открытом состоянии (время открытого состояния) возрастает по мере увеличения деполяризации
 Анализ работы одиночных К-каналов 1. одиночный К-канал работает в режиме случайных переходов из закрытого состояния - в открытое, по принципу «все или ничего» 2. При увеличении деполяризации растет время пребывания в открытом состоянии
Анализ работы одиночных К-каналов 1. одиночный К-канал работает в режиме случайных переходов из закрытого состояния - в открытое, по принципу «все или ничего» 2. При увеличении деполяризации растет время пребывания в открытом состоянии
 ЗАКЛЮЧЕНИЕ • Интегральные натриевый и калиевый токи, регистрируемые при генерации ПД, отражают потенциало-зависимость и временной ход натриевой и калиевой проводимости мембраны. • Изменения g. Na и g. K отражают особенности перехода и пребывания в открытом состоянии отдельных натриевых и калиевых каналов, а также инактивированного состояния отдельных натриевых каналов. The Nobel Prize in Physiology or Medicine 1963 "for their discoveries concerning the ionic mechanisms involved in excitation and inhibition in the peripheral and central portions of the nerve cell membrane" Alan Lloyd Hodgkin Andrew Fielding Huxley
ЗАКЛЮЧЕНИЕ • Интегральные натриевый и калиевый токи, регистрируемые при генерации ПД, отражают потенциало-зависимость и временной ход натриевой и калиевой проводимости мембраны. • Изменения g. Na и g. K отражают особенности перехода и пребывания в открытом состоянии отдельных натриевых и калиевых каналов, а также инактивированного состояния отдельных натриевых каналов. The Nobel Prize in Physiology or Medicine 1963 "for their discoveries concerning the ionic mechanisms involved in excitation and inhibition in the peripheral and central portions of the nerve cell membrane" Alan Lloyd Hodgkin Andrew Fielding Huxley
 Спасибо за внимание… Вопросы? ? ?
Спасибо за внимание… Вопросы? ? ?