Физиология возбудимых тканей.ppt
- Количество слайдов: 125



Нейрон – как структурнофункциональная единица ЦНС Мембранный электрогенез к. м. н. , доцент Сорокин О. В.

Содержание лекции Принципы рефлекторной регуляции функций Понятия раздражимости и возбудимости Строение нейрона Строение и физиологические функции мембраны клеток возбудимых тканей Мембранный потенциал покоя Потенциал действия Локальный ответ Проведение возбуждения по нервному волокну. Строение синапса и проведение возбуждения в синапсах Медиаторы их виды и физиологическая роль Механизмы формирования ВПСП и ТПСП

Механизмы управления Регуляция Гуморальная Изменение физиологической активности органов и систем под влиянием химических веществ, доставляемых через жидкие среды организма (интерстициальная жидкость, лимфа, кровь, цереброспинальная жидкость) Нервная Изменение физиологических функций под влиянием управляющих воздействий, передаваемых из ЦНС по нервным волокнам к органам

Характеристика гуморальной регуляции Филогенетически является более древней формой взаимодействия клеток ◦ Изменение деятельности под влиянием продуктов обмена веществ (СО 2, АДФ) ◦ Специализированная секреция БАВ (гормонов) Медленное распространение с током биологических жидкостей Диффузный характер управляющих воздействий Продолжительное действие
 Аутокринная – изменения функции клетки химическими субстратами, выделяемыми в межклеточную")
Формы управления (гуморальная регуляция) Аутокринная – изменения функции клетки химическими субстратами, выделяемыми в межклеточную среду самой клеткой Паракринная – химические агенты, выделяемые одной клеткой, распространяются по интерстицию и управляют функцией других клеток, удалённых от источника управляющих воздействий Телекринная – БАВ выделяются в кровь и достигают всех органов и тканей

Характеристика нервной регуляции Филогенетически более молодая форма регуляции Высокая скорость распространения информации Точная, «адресная» передача объекту регулирования управляющих воздействий Высокая надёжность осуществления связи Как правило, кратковременна В основе лежит рефлекс и рефлекторная дуга

Рефлекс Ответная реакция организма на раздражение рецепторов, которая заключается в возникновении, изменении или прекращении функциональной активности органов, тканей или целостного организма и осуществляется при обязательном участии центральной нервной системы.

Рефлекторная дуга Морфологическая основа рефлекса, состоящая из 5 -ти звеньев: ◦ Рецептор ◦ Афферентное ◦ Центральное ◦ Эфферентное ◦ Эффектор
 – заключается в том, что для осуществления рефлекса нужна")
Рефлекторная регуляция функций Детерминизм (причинности) – заключается в том, что для осуществления рефлекса нужна причина, следовательно, действие раздражителя. Принцип структурности – абсолютно необходимым условием для осуществления рефлекса является структурная и функциональная целостность всех звеньев рефлекторной дуги. Принцип анализа и синтеза – рефлекторная реакция осуществляется на основании анализа входящей информации, которая поступает из нескольких источников и синтеза – принятия решения на основании этой информации.

Схема рефлекторной дуги

Нейрон – структурнофункциональная единица ЦНС

Отделы нейрона Дендриты — ветвящиеся отростки, заканчивающиеся вблизи от тела Дендриты нейрона. В функциональном отношении дендриты — рецепторная зона регистрации сигналов от других нервных клеток. Поверхностная мембрана дендритов и перикариона одного нейрона головного мозга образует до 20 000 синаптических контактов с другими нейронами. Тело нейрона содержит ядро, комплекс Гольджи, гранулярную эндоплазматическую сеть, митохондрии, лизосомы, элементы цитоскелета. Синтез белка происходит только в перикарионе. Аксонный холмик (начальный сегмент аксона) — место, где начинается аксон и происходит генерация ПД. Аксон — длинный отросток, как правило, не ветвящийся. Объём аксона Аксон может достигать 99% суммарного объёма нейрона. Длина аксона может быть значительной — десятки сантиметров. Основной функцией аксона является проведение ПД. Терминали аксона (концевые разветвления) принимают участие в образовании синапсов с другими нейронами, мышечными и секреторными клетками. Терминали заканчивается утолщениями, содержащими синаптические пузырьки (везикулы).

Рефлекторная дуга

Рецепторы Образования, способные воспринять энергию раздражения, трансформировать её в нервный импульс и передать информацию в ЦНС
. 3. Хеморецепторы (р. Н, напряжение газов, концентрация")
Модальность рецепторов 1. Тактильные 2. Болевые (ноцицепторы). 3. Хеморецепторы (р. Н, напряжение газов, концентрация электролитов) 4. Осморецепторы 5. Терморецепторы 6. Механорецепторы (барорецепторы: давление, растяжение) 7. Высокоспециализированные клетки: рецепторы органов вкуса, обоняния (хеморецепторы), зрения (фоторецепторы), слуха, вестибулярного аппарата (механорецепторы).

Виды рецепторов

Афферентное звено Представлено отростками нейронов, которые связывают рецепторы с нервными центрами, обеспечивают центростремительную передачу возбуждения в структуры ЦНС Относятся к группе А нервных волокон (скорость передачи возбуждения 80120 м/с)

Нервные центры Совокупность нервных клеток, «ансамбль» нейронов, которые включаются в регуляцию определённой функции или в осуществлении конкретного рефлекторного акта. В зависимости от расположения НЦ: ◦ Спинальные, бульбарные, мезенцефальные, кортикальные В соответствии с выполняемой функцией: ◦ Чувствительные, вегетативные, двигательные, системно-организованные центры (сосудодвигательные, дыхательные, пищеварительные)

Эфферентное звено Путь, по которому возбуждение распространяется центробежно, от ЦНС на периферию, к рабочим органам.

Эффекторы Скелетные, гладкие, сердечная мышцы Железы

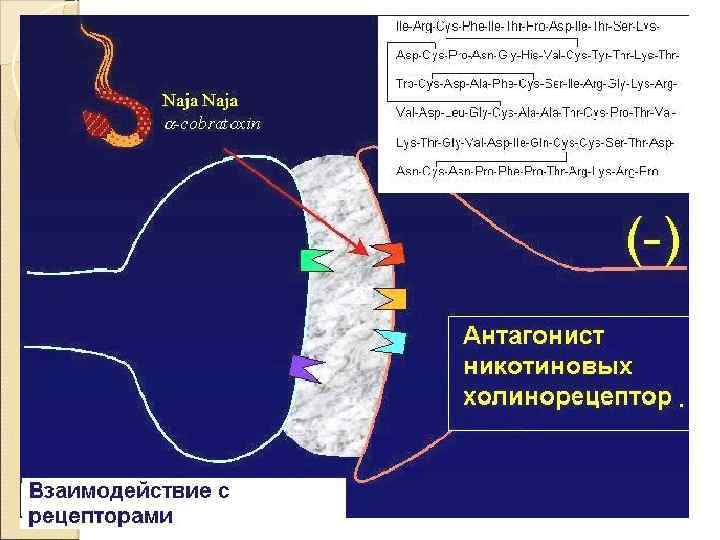
Двигательные нейроны рефлексов расположены в передних рогах серого вещества спинного мозга, эфферентные пути не прерываются, эффектор – скелетные мышцы, медиатор в нервно-мышечном синапсе – ацетилхолин, мембранные рецепторы – холинорецепторы никотинового типа

Скелетные мышцы

Вегетативные рефлексы Вегетативные : ◦ Симпатические ◦ Парасимпатические Виды влияния на органы: ◦ Пусковое – побуждает орган к совершению работы ◦ Корригирующее – усиливает или ослабляет деятельность органа (ЧСС) ◦ Адаптационно-трофическое – изменение обмена веществ рабочего органа, восстанавливающего функции

Вегетативная нервная система симпатический отдел Двигательные нейроны симпатических рефлексов расположены в боковых рогах грудного и поясничного отделов спинного мозга; эфферентные нервные волокна прерываются в ганглиях симпатического ствола, чревном и брыжеечном сплетении; медиатор в преганглионарных волокнах – АХ, мембранные рецепторы Н-хр, в постганглионарных – норадреналин, мембранные рецепторы на эффекторе – а и b - адренорецепторы

Вегетативная нервная система парасимпатический отдел Двигательные нейроны рефлексов расположены в среднем мозге, районе моста, продолговатом мозге и в 1 -5 крестцовых сегментов спинного мозга; Эфферентные волокна прерываются в ганглиях интрамурально или параорганно; Медиатор преганглионарного волокна – АХ, мембранные рецепторы – Н-хр, В постганглионарных – АХ, мембранные рецепторы на эффекторе – М-хр

Сердечная мышца

Железы

Баланс отделов ВНС

Мембранный электрогенез Возбудимые ткани: ◦ Мышечная – сокращение ◦ Железистая - секреция ◦ Нервная – генерация и проведение нервного импульса
 направлены")
Цитоплазматическая мембрана Биологическая мембрана состоит из двойного слоя фосфолипидов, гидрофильные части которых (головки) направлены к поверхности мембраны, а гидрофобные части (хвосты, стабилизирующие мембрану в виде бислоя) внутрь мембраны. И — интегральные белки погружены в мембрану. Т — трансмембранные белки пронизывают всю толщу мембраны. П — периферические белки расположены либо на наружной, либо на внутренней поверхности мембраны.

Виды белков мембраны клетки Интегральные мембранные белки – встроены в липидный бислой. К интегральным мембранным белкам относятся белки адгезии, некоторые рецепторные белки (мембранные рецепторы). Трансмембранный белок — молекула белка, проходящая через всю толщу мембраны и выступающая из неё как на наружной, так и на внутренней поверхности. К трансмембранным белкам относятся поры, ионные каналы, переносчики, насосы, некоторые рецепторные белки. ◦ Поры и каналы — трансмембранные пути, по которым между цитозолем и межклеточным пространством (и в обратном направлении) перемещаются вода, ионы и молекулы метаболитов. ◦ Переносчики осуществляют трансмембранное перемещение конкретных молекул (в том числе в сочетании с переносом ионов или молекул другого типа). ◦ Насосы перемещают ионы против их концентрационного и энергетического градиентов (электрохимический градиент) при помощи энергии, освобождаемой при гидролизе АТФ. Периферические мембранные белки находятся на одной из поверхностей клеточной мембраны (наружной или внутренней) и нековалентно связаны с интегральными мембранными белками. ◦ Примеры периферических мембранных белков, связанных с наружной поверхностью мембраны — рецепторные белки и белки адгезии. ◦ Примеры периферических мембранных белков, связанных с внутренней поверхностью мембраны, — белки цитоскелета, белки системы вторых посредников, ферменты и другие белки.

Трансмембранный транспорт Пассивный транспорт ◦ Осмос ◦ Диффузия ◦ Облеченная диффузия Активный транспорт ◦ Первично активный ◦ Вторично активный
 — движение небольших неполярных и полярных молекул в обоих направлениях")
Пассивный транспорт (пассивная диффузия) — движение небольших неполярных и полярных молекул в обоих направлениях по градиенту концентрации (разность химического потенциала) или по электрохимическому градиенту (транспорт заряженных веществ — электролитов) осуществляется без затрат энергии и характеризуется низкой специфичностью.

Пассивный транспорт путём диффузии через мембрану А — направление транспорта вещества как при простой, так и при облегчённой диффузии происходит по градиенту концентрации вещества по обе стороны плазмолеммы. Б — кинетика транспорта. По ординате — количество диффундировавшего вещества, по ординате — время. Простая диффузия не требует непосредственных затрат энергии, является ненасыщаемым процессом, её скорость линейно зависит от градиента концентрации вещества.

Облегчённая диффузия Для облегчённой диффузии веществ необходимы встроенные в мембрану белковые компоненты (поры, переносчики, каналы). Все эти компоненты относятся к интегральным (трансмембранным) белкам. Облегчённая диффузия происходит по градиенту концентрации для неполярных веществ или по электрохимическому градиенту для полярных веществ

Поры По определению, заполненный водой канал поры всегда открыт. Поры формируют разные белки (порины, перфорины, аквапорины, коннексины и др. ). В некоторых случаях образуются гигантские комплексы (например, ядерные поры), состоящие из множества разных белков.
 осуществляют транспорт через биологические мембраны множества различных ионов (Na+, Cl–, H+, HCO")
Переносчики (транспортёры) осуществляют транспорт через биологические мембраны множества различных ионов (Na+, Cl–, H+, HCO 3– и др. ) и органических веществ (глюкоза, аминокислоты, креатин, норадреналин, фолаты, лактат, пируват и др. ). Транспортёры специфичны: каждый конкретный переносчик переносит через липидный бислой, как правило и по преимуществу, одно вещество. Различают однонаправленный (унипорт), сочетанный (симпорт) и разнонаправленный (антипорт) транспорт. Унипорт — однонаправленный транспорт одного вещества. Симпорт (котранспорт). Некоторые переносчики осуществляют однонаправленный сочетанный транспорт одного вещества. Антипорт (обменник). Эти переносчики осуществляют разнонаправленный транспорт двух разных веществ (встречный транспорт).

Ионные каналы Состоят из связанных между собой белковых молекул, формирующих в мембране гидрофильную пору. Через открытую пору по электрохимическому градиенту диффундируют ионы. Свойства ионных каналов (в том числе специфичность и проводимость) определяют как аминокислотная последовательность конкретного полипептида, так и конформационные изменения, происходящие с разными частями полипептидов в составе интегрального белка канала

Ионные каналы
 по отношению к конкретным катионам и анионам")
Характеристики каналов Специфичность. Ионные каналы специфичны (селективны) по отношению к конкретным катионам и анионам (например, для Na+ [натриевый канал], K+ [калиевый канал], Ca 2+ [кальциевый канал], Cl– [хлорный канал] и др. ). Проводимость определяется количеством ионов, способных пройти через канал в единицу времени. Проводимость канала изменяется в зависимости от того, открыт канал или закрыт. Активность. Канал может находиться либо в открытом, либо в закрытом состоянии. Поэтому модель канала предусматривает наличие устройства, открывающего и закрывающего канал — воротный механизм, или ворота канала (по аналогии с открытыми и закрытыми воротами).

Функциональные компоненты Помимо ворот, модель ионного канала ворот предусматривает существование таких функциональных компонентов, как сенсор, сенсор избирательный фильтр и пора открытого канала. фильтр канала
 сенсоров к разным типам сигналов: изменениям мембранного")
Сенсор Каждый канал имеет один (иногда больше) сенсоров к разным типам сигналов: изменениям мембранного потенциала (МП), вторым посредникам (с цитоплазматической стороны мембраны), разным лигандам (с внеклеточной стороны мембраны). Эти сигналы регулируют переход между открытым и закрытым состоянием канала.

Как открываются каналы? при изменении потенциала мембраны - потенциал-зависимые. в результате взаимодействия рецептора с биологически активным веществом (гормоном, медиатором). рецептор-управляемые. при механическом воздействии
. Колебания МП приводят к конформационным изменениям белковой")
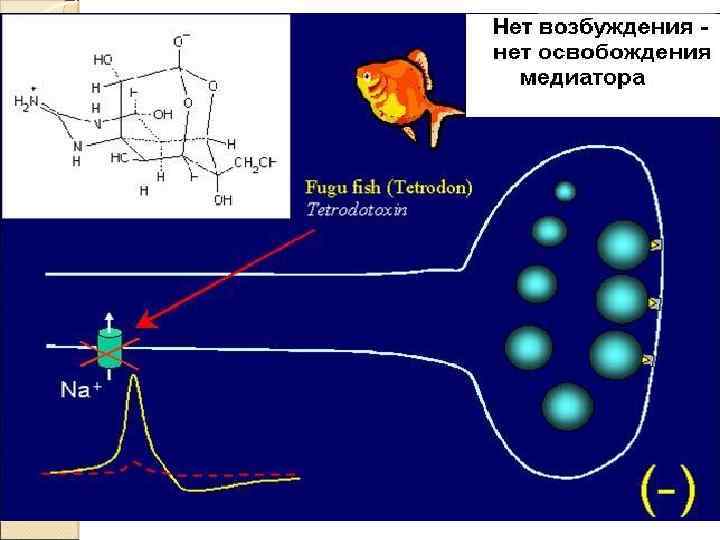
Потенциалозависимые ионные каналы Управляются мембранным потенциалом (МП). Колебания МП приводят к конформационным изменениям белковой структуры канала, что и переводит канал в открытое либо закрытое состояние. Благодаря наличию потенциалозависимых Na+‑каналов в возбудимых структурах (например, плазмолемма мышечных клеток и аксолемма нервных клеток) происходит генерация и распространение потенциала действия (ПД).
 ионные каналы соединение лиганда с рецептором вызывает конформационные изменения в канале,")
Лиганд–зависимые (рецептор– зависимые) ионные каналы соединение лиганда с рецептором вызывает конформационные изменения в канале, изменяющие его функциональное состояние. менее избирательны, пропускают несколько разных, но одинаково заряженных ионов. никотиновый холинорецептор мышц в физиологических условиях проводит Na+ внутрь клетки.

Механочувствительные ионные каналы открываются при деформации мембраны (например, волосковые клетки внутреннего уха или чувствительная нервная терминаль механорецепторов — телец Пачини).

Состояния канала Благодаря наличию ворот, сенсора, избирательного фильтра и поры ионные каналы могут быть в состояниях покоя, активации и инактивации. Состояние покоя — канал закрыт, но готов к открытию в ответ на химические, механические или электрические стимулы. Состояние активации — канал открыт и пропускает ионы. Состояние инактивации — канал закрыт и не способен к активации. Инактивация возникает тотчас после открытия канала в ответ на действие стимула и длится от нескольких до нескольких сотен миллисекунд (в зависимости от типа канала).

Активный транспорт Энергозависимый трансмембранный перенос против электрохимического градиента. Различают первичный и вторичный активный транспорт. Первичный активный транспорт осуществляют насосы (различные насосы АТФазы), вторичный — симпортёры (сочетанный однонаправленный транспорт) и антипортёры (встречный антипортёры разнонаправленный транспорт).

Виды активного транспорта Первичный активный транспорт. Движущая сила трансмембранного переноса возникает при ферментативном гидролизе макроэргических связей АТФ. Родовой термин для таких АТФаз (например, Na+, K+-, H+, K+, Ca 2+‑АТФазы) — насосы. Вторичный активный транспорт. Движущая сила для трансмембранного переноса одного вещества (или ионов) против электрохимического градиента возникает за счёт потенциальной энергии, запасённой за счёт сочетанного переноса ионов (как правило, Na+) по электрохимическому градиенту. В большинстве случаев поступление Na+ в цитозоль из межклеточного пространства и обеспечивает вторичный активный транспорт разных ионов и веществ. Известно 2 типа вторичного активного переноса — симпорт и антипорт.

Внеклеточная жидкость АТФ Цитоплазма Мембрана клетки

Итог работы насоса + из клетки 3 иона Na + в клетку 2 иона K 3 Na+ АТФ 2 K+
")
Концентрации ионов внутри и снаружи клетки (ммоль/л)

Возбудимая клетка поляризована

МП покоя разность электрических потенциалов между внутренней и наружной поверхностью мембраны в состоянии покоя, т. е. при отсутствии электрического или химического раздражителя (сигнала). в состоянии покоя поляризация внутренней поверхности клеточной мембраны имеет отрицательную величину, поэтому значение МП покоя тоже отрицательно.

Состояние покоя

РАВНОВЕСНЫЙ калиевый ПОТЕНЦИАЛ Поток ионов калия прекращается, когда действие электрического поля компенсирует движение иона по градиенту концентрации. Следовательно, для данной разности концентраций ионов на мембране формируется так называемый РАВНОВЕСНЫЙ ПОТЕНЦИАЛ для калия.

Значение потенциала Нернста где z. Х — валентность X, T — абсолютная температура, R — газовая постоянная, F — константа Фарадея, [X] — концентрация иона X с наружной (сн) и внутренней (вн) поверхности мембраны.
![Равновесный потенциал по К+ EK+ = – 61 м. В·ln([K+вн]/[K+сн]) так как [K+вн]/[K+н] =](https://present5.com/presentation/9354683_184434808/image-58.jpg "Равновесный потенциал по К+ EK+ = – 61 м. В·ln([K+вн]/[K+сн]) так как [K+вн]/[K+н] =")
Равновесный потенциал по К+ EK+ = – 61 м. В·ln([K+вн]/[K+сн]) так как [K+вн]/[K+н] = 35, то EK+ = – 94 м. В PK : PNa : PCl = 1 : 0, 04 : 0, 45
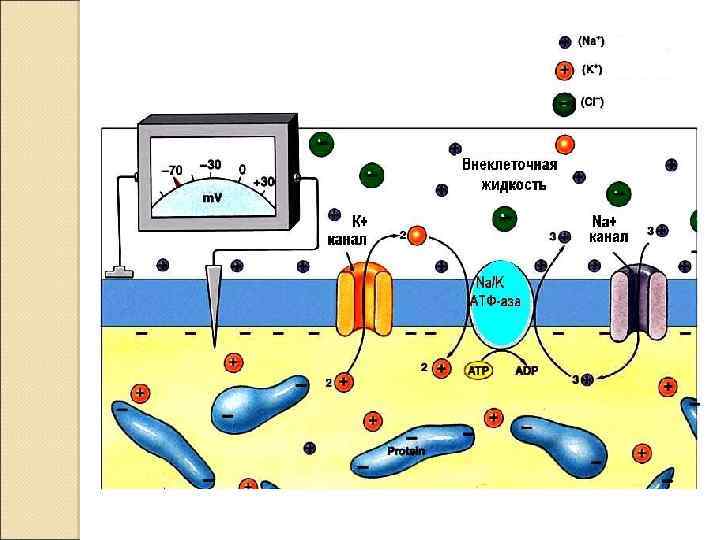
 происходит в результате повышения проницаемости")
Возбуждение клетки Переход от покоя к активному состоянию (возбуждение) происходит в результате повышения проницаемости ионных каналов для Na+

Причиной изменения проницаемости – раздражителем - может быть 1. 2. 3. 4. 5. механическое воздействие изменение потенциала мембраны взаимодействие мембранных рецепторов с биологически активным веществом изменение химизма среды изменение температуры.

Раздражители: По силе: подпороговые и пороговые 2. По биологической значимости: адекватные и неадекватные 1.

Натрий устремляется в клетку, возникает ионный ток и происходит снижение мембранного потенциала - деполяризация мембраны.

При стимуляции поляризация переходит в деполяризацию Before stimulation Inside Neuron After stimulation Na A K+
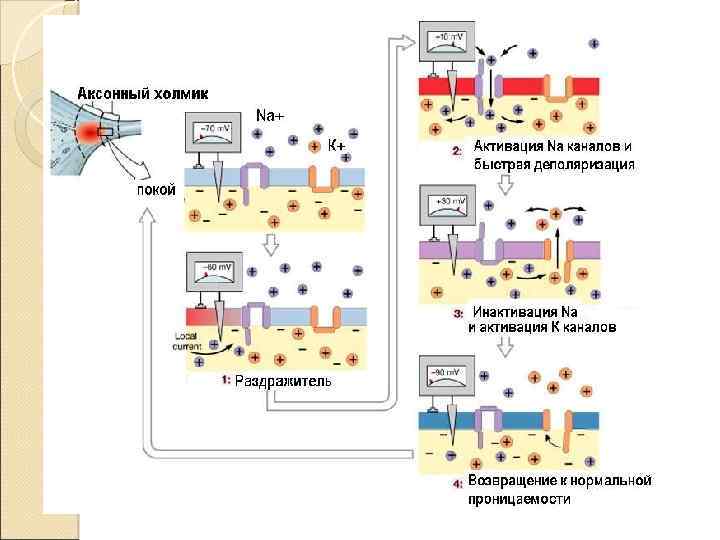

У всех возбудимых клеток существует такой уровень деполяризации – уменьшения отрицательного заряда мембраны – при котором активируются все быстрые, потенциал-зависимые натриевые каналы Минимальная сила раздражителя, которая необходима для возникновения возбуждения, называется ПОРОГОМ ВОЗБУЖДЕНИЯ или порогом реакции Мерой возбудимости ткани служит порог возбуждения.
")
. Этот уровень деполяризации называется критическим уровнем деполяризации (КУД)

Изменения мембранного потенциала принято отображать графически

МПП

Изменение потенциала мембраны
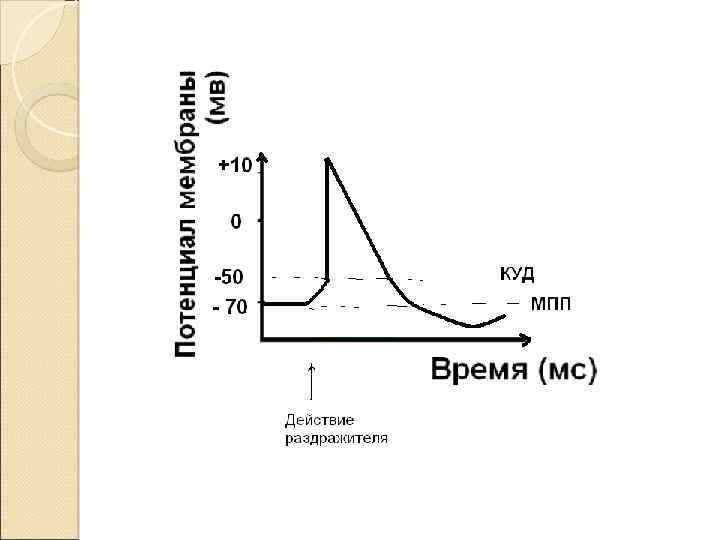

Быстрое изменение потенциала мембраны в ответ на действие раздражителя пороговой силы называется потенциал действия (ПД)

ПД характеризуется: 1. амплитудой 2. длительностью

Состояние каналов при ПД

Фазы ПД
 1.")
Три электрические характеристики мембраны МПП 2. КУД 3. Овершут (точка реверса потенциала) 1.

В какое время при ПД мембрана способна ответить на раздражитель

Во время ПД есть период, когда мембрана невозбудима - рефрактерность

Изменение возбудимости по фазам ПД

Изменение потенциала мембраны в ответ на действие раздражителя подпороговой силы – называется локальный ответ - ЛО Деполяризация не достигает КУД

Изменение возбудимости при ЛО

Зависимость ЛО от силы раздражителя

Сравнительная характеристика ЛО и ПД ЛО 1. ПД не зависит от пропорционален силе силы раздражителя и раздражителя. подчиняется закону «все или ничего» 2. ЛО может суммироваться до тех 2. ПД не суммируется 1. пор, пока изменения мембранного потенциала не достигнут КУД 3. ЛО не передается по мембране 3. ПД передается по мембране
 называют специализированные контакты между нервными")
Физиология синапсов Синапсами (от греч. synapsis — соприкосновение, соединение) называют специализированные контакты между нервными клетками или между нервными и эффекторными клетками, используемые для передачи сигналов. Термин «синапс» предложил в 1897 г. Чарлз Шеррингтон

Классификация синапсов По местоположению ◦ нервно-мышечные ◦ нейрональные, аксосоматические, аксональные, аксодендритические, дендросоматические. По характеру действия ◦ возбуждающими и тормозящими. По способу передачи сигнала ◦ электрические, химические, смешанные.

Строение синапса

Нервно-мышечный синапс

Строение химического синапса Пресинаптическая терминаль Синаптическя щель Постсинаптическое окончание

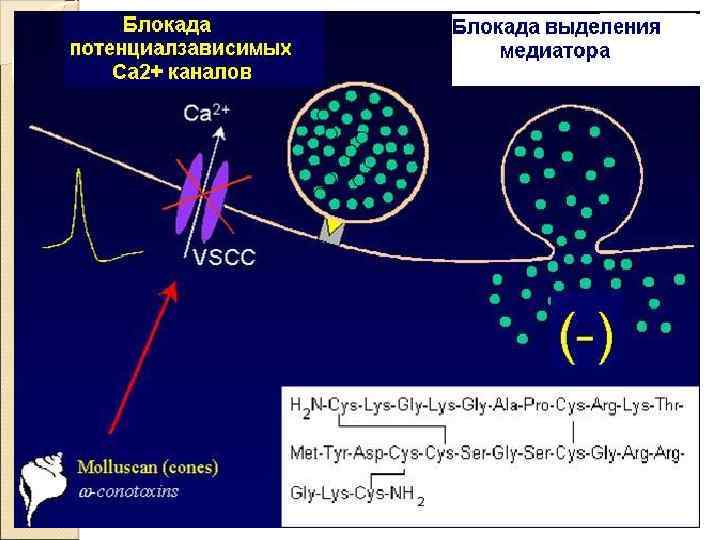
Пресинаптическая часть синаптические пузырьки с нейромедиатором элементы цитоскелета и митохондрии потенциалозависимые Ca 2+‑каналы активные зоны экзоцитоза медиатора транспортёры, осуществляющие перенос нейромедиаторов в пресинаптическую терминаль

Синаптические гранулы Гранулы содержат химическое вещество – медиатор Гранулы могут сливаться с пресинаптической мембраной и выделять медиатор в синаптическую щель
 ◦ Кислые (глицин, ГАМК) Амины ◦ Моноамины")
Классификация нейромедиаторов Аминокислоты ◦ Нейтральные (глутамат, аспартат) ◦ Кислые (глицин, ГАМК) Амины ◦ Моноамины (ацетилхолин, серотонин, гистамин) ◦ Катехоловые амины (адреналин, норадреналин, дофамин) Нейропептиды (ТТГ-РГ, метионин- и лейцин-энкефалины; ангиотензин II, холецистокинин-подобный пептид, окситоцин, соматостатин, люлиберин, вещество Р, нейротензин, гастринрелизинг-пептид, аргинин взопрессин, b-эндорфин, АКТГ) Пурины (АТФ и аденозин) Газы (оксид азота)

Транспорт медиатора

Жизненный цикл синаптических пузырьков

Роль ионов Са 2+ Белок синаптических пузырьков – синаптотагмин реорганизует цитоскелет везикул при взаимодействии с Са 2+ обеспечивая экзоцитоз нейромедиатора

Синаптическая щель промежуток между пре- и постсинаптическими мембранами шириной 20– 35 нм находятся ферменты, расщепляющие молекулы нейромедиатора (например, ацетилхолинэстераза, гидролизующая ацетилхолин)

Судьба нейромедиатора Инактивация нейромедиатора. Кратковременность взаимодействия нейромедиатора с рецептором достигается разрушением нейромедиатора специальными ферментами (например, ацетилхолина — ацетилхолинэстеразой). Захват нейромедиатора. В большинстве синапсов передача сигналов прекращается вследствие быстрого захвата нейромедиатора пресинаптической терминалью.

Постсинаптическая терминаль Рецепторы, чувствительные к нейромедиатору Постсинаптические плотности. Область, расположенная с цитоплазматической стороны постсинаптической мембраны (рецепторы нейромедиаторов, протеинкиназы, структурные и цитоскелетные белки, а также белки, осуществляющие эндоцитоз и гликолиз) Дендритные шипики — характерная особенность 90% возбуждающих синапсов в ЦНС. Эти мелкие (менее 1 мкм длиной) удлинённые структуры, выступающие над поверхностью дендритов и имеющие постсинаптические плотности. Их функция точно не установлена; возможно, они увеличивают поверхность постсинаптической стороны или являются резервуаром для Ca 2+.

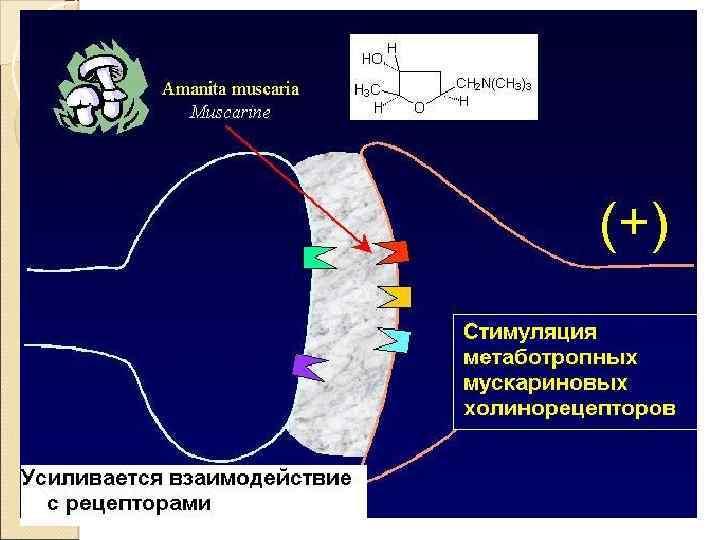
Рецепторы постсинапса Характер электрического ответа постсинаптической стороны и дальнейший физиологический эффект определяются свойствами рецепторов Ионотропные рецепторы сами по себе являются ионными каналами. Классический пример — н‑холинорецепторы Метаботропные рецепторы связаны с ферментами (аденилатциклаза или фосфолипаза C) через G‑белок. Классический пример — м‑холинорецепторы
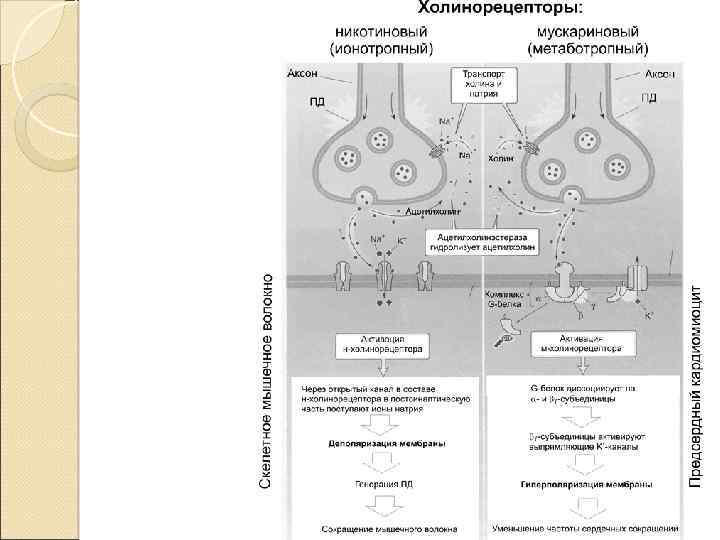
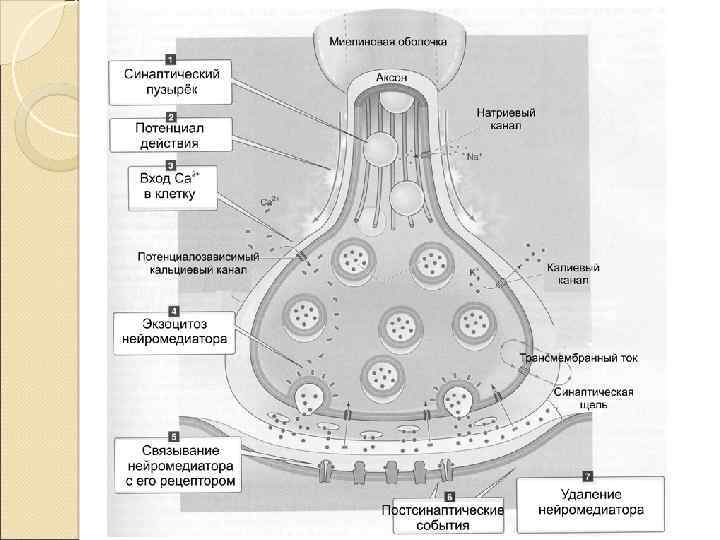

Этапы синаптической передачи 1. Молекулы нейромедиатора поступают в мембранные синаптические пузырьки, располагающиеся в пресинаптической терминали и концентрирующиеся в активных зонах пресинаптической мембраны. 2. Приходящий по аксону ПД деполяризует пресинаптическую мембрану. 3. Вследствие деполяризации открываются потенциалозависимые Са 2+‑каналы, и Са 2+ поступает в терминаль. 4. Увеличение внутриклеточного [Са 2+] запускает слияние синаптических пузырьков с пресинаптической мембраной и выброс нейромедиатора в синаптическую щель (экзоцитоз). 5. Кванты нейромедиатора, поступившие в синаптическую щель, диффундируют в ней. Часть молекул нейромедиатора связывается со специфичными для них рецепторами постсинаптической мембраны. 6. Связавшие нейромедиатор рецепторы активированы, что приводит к изменению поляризации постсинаптической мембраны либо прямо (поступление ионов через ионотропные рецепторы) либо опосредованно — активация ионных каналов через систему G‑белка (метаботропные рецепторы). 7. Инактивация нейромедиаторов происходит либо путём их ферментной деградации, либо молекулы нейромедиатора захватываются клетками.
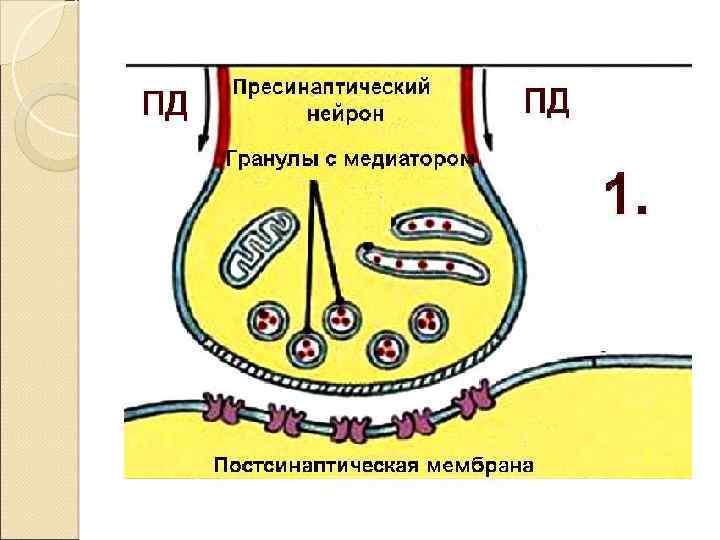
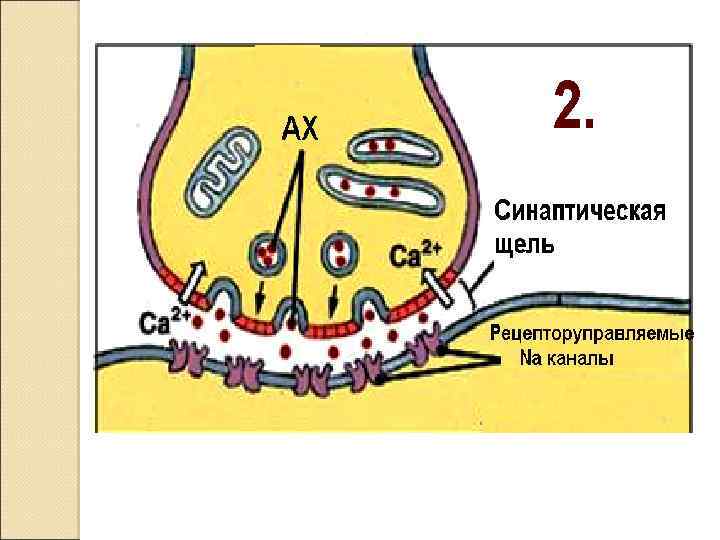
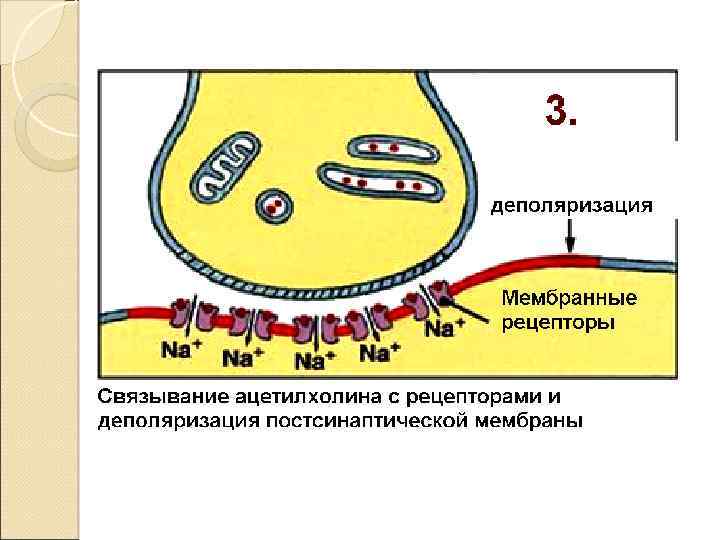
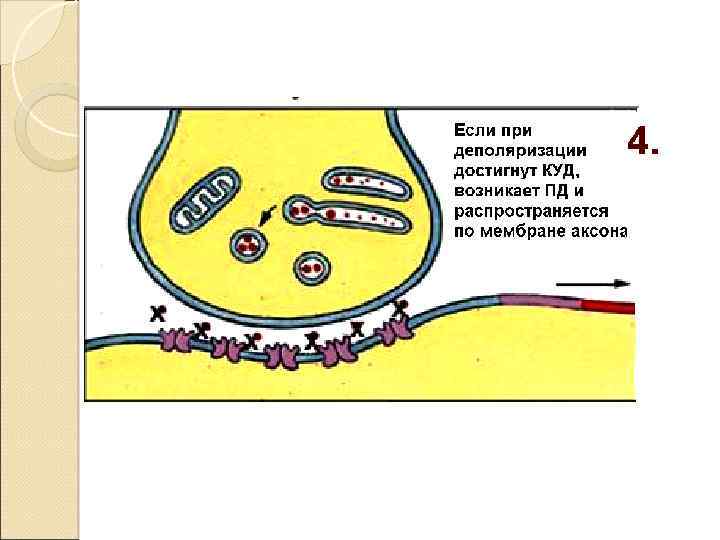

Закономерности проведения импульса в синапсе Одностороннее проведение Синаптическая задержка (0. 5 – 2. 5 мсек) Трансформация ритма Взаимодействие синапсов (регуляция) Высокая утомляемость Чувствительность к токсинам, недостатку О 2 !! Формирование следовых процессов (тренировка синапсов)

Регуляция деятельности синапсов

Аксо-аксональный синапс

Пресинаптическое ингибирование

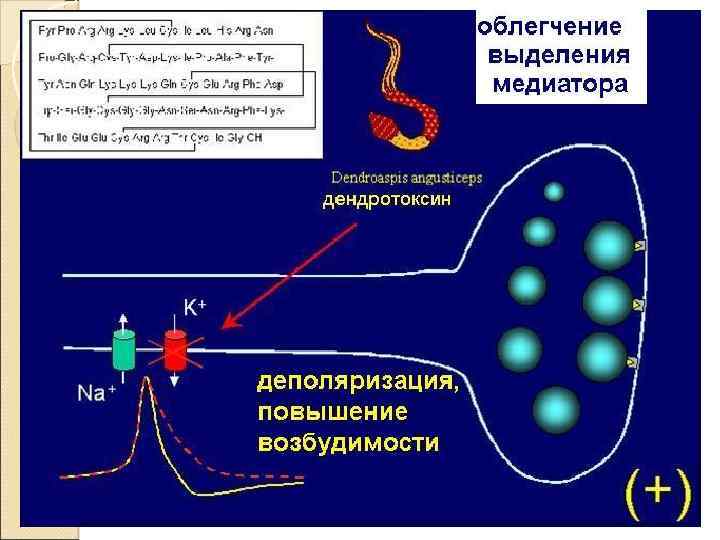
Пресинаптическое облегчение

Синаптоактивные вещества

Симпатический синапс

Парасимпатический синапс

Возбуждающие и тормозные синапсы

Изменение потенциала постсинаптической мембраны возбуждающего тормозного

Суммация – МП =

Возникновение ПД является результатом суммации ВПСП и ТПСП на нейроне
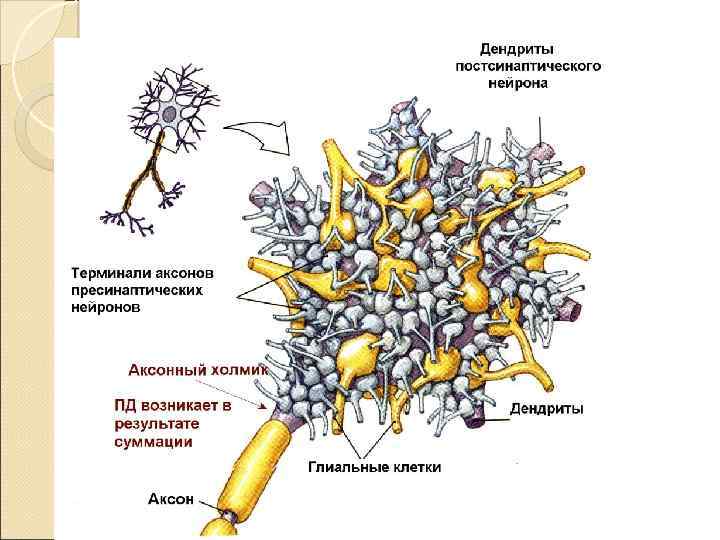
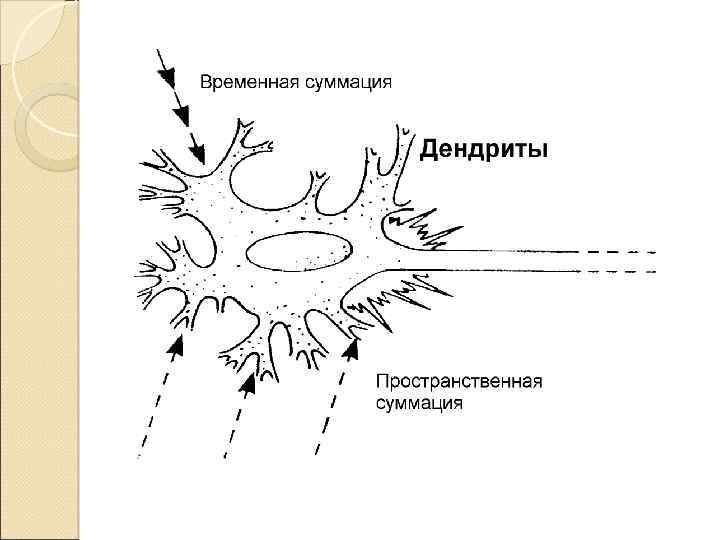

-4 Суммация +6 -12 -4 +6 +6 +10 ПД dendrites Cell body axon
Физиология возбудимых тканей.ppt