

ОМФ 15 Глутамат.ppt
- Количество слайдов: 34
 Нейромедиаторные системы мозга Глутамат, энкефалины
Нейромедиаторные системы мозга Глутамат, энкефалины
 v Глутамат - главный возбуждающий нейротрансмиттер ЦНС, повсеместно распространённый в коре и подкорковых структурах мозга. v Глутамат, высвобождаемый из аксональных пресинаптических терминалей, генерирует электрическое возбуждение в постсинаптических нейронах.
v Глутамат - главный возбуждающий нейротрансмиттер ЦНС, повсеместно распространённый в коре и подкорковых структурах мозга. v Глутамат, высвобождаемый из аксональных пресинаптических терминалей, генерирует электрическое возбуждение в постсинаптических нейронах.
 v Глутамат – ключевая молекула в обучении, памяти и клеточном метаболизме. v Различные глутаматные рецепторные системы играют различные роли в передаче информации, пластичности нейронов.
v Глутамат – ключевая молекула в обучении, памяти и клеточном метаболизме. v Различные глутаматные рецепторные системы играют различные роли в передаче информации, пластичности нейронов.
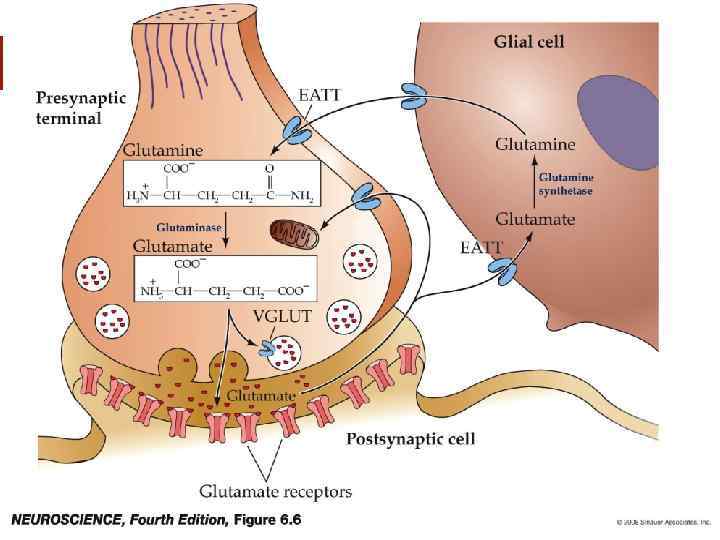
 Распространение глутамата v Глутаматергические нейроны широко распространены в коре ГМ. Они отдают проекции к большому числу подкорковых структур: включая гиппокамп, базолатеральный комплекс амигдалы, черную субстанцию и т. д.
Распространение глутамата v Глутаматергические нейроны широко распространены в коре ГМ. Они отдают проекции к большому числу подкорковых структур: включая гиппокамп, базолатеральный комплекс амигдалы, черную субстанцию и т. д.
 – EAAT 1 -4,") Транспортёры глутамата v Идентифицированы 4 типа транспортёров возбуждающих аминокислот (EAAT) – EAAT 1 -4, EAAT 1, EAAT 2, EAAT 3. EAAT 1 и 2 представлены в основном в глие, EAAT 3 – в нейронах. v Существуют блокаторы захвата глутамата (дигидрокаинат и др. ).
Транспортёры глутамата v Идентифицированы 4 типа транспортёров возбуждающих аминокислот (EAAT) – EAAT 1 -4, EAAT 1, EAAT 2, EAAT 3. EAAT 1 и 2 представлены в основном в глие, EAAT 3 – в нейронах. v Существуют блокаторы захвата глутамата (дигидрокаинат и др. ).
 v Транспортёры необходимы для снижения уровня глутамата ниже порога, для избегания повреждения нейронов. v Уменьшение транспортной способности связано с последующим увеличением нейротоксичности.
v Транспортёры необходимы для снижения уровня глутамата ниже порога, для избегания повреждения нейронов. v Уменьшение транспортной способности связано с последующим увеличением нейротоксичности.
 v Примером нейрональной дегенерации, связанной с уменьшением транспортёров глутамата является боковой амиотрофический склероз.
v Примером нейрональной дегенерации, связанной с уменьшением транспортёров глутамата является боковой амиотрофический склероз.
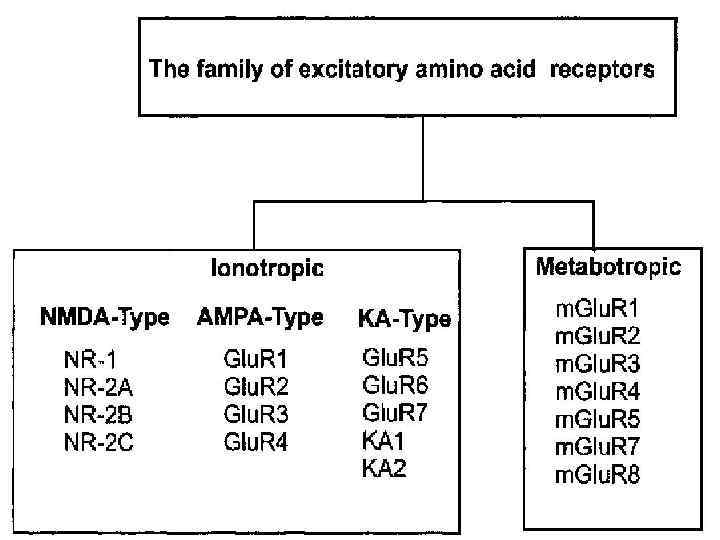
 состоят из 4 -х субъединиц (2 NR 1 -СЕ") Рецепторы глутамата v NMDA-рецепторы (N-methyl-Daspartate) состоят из 4 -х субъединиц (2 NR 1 -СЕ + 2 NR 2 -СЕ), способны пропускать ионы Na, K и Са. v NMDA-рецепторы отвечают на глутамат в относительно медленной манере (по сравнению с другими ионотропными рецепторами). Этот медленный ответ связан с тем, что ионы Mg тонически ингибируют NMDA-рецепторы.
Рецепторы глутамата v NMDA-рецепторы (N-methyl-Daspartate) состоят из 4 -х субъединиц (2 NR 1 -СЕ + 2 NR 2 -СЕ), способны пропускать ионы Na, K и Са. v NMDA-рецепторы отвечают на глутамат в относительно медленной манере (по сравнению с другими ионотропными рецепторами). Этот медленный ответ связан с тем, что ионы Mg тонически ингибируют NMDA-рецепторы.
 v Рецептор состоит из 5 трансмембранных белков с различными специфическими местами связывания (для NMDA, глицина), ассоциированными с ионным каналом (два места связывания – с Mg и некоторыми другими веществами).
v Рецептор состоит из 5 трансмембранных белков с различными специфическими местами связывания (для NMDA, глицина), ассоциированными с ионным каналом (два места связывания – с Mg и некоторыми другими веществами).
 v В состоянии покоя NMDA-рецепторы не активны, а ионный канал блокирован Mg. v При деполяризации мембраны блок снимается и в клетку входит Na, K и Са. v Са способен изменять свойства нейронов. Избыток Са токсичен для нейронов.
v В состоянии покоя NMDA-рецепторы не активны, а ионный канал блокирован Mg. v При деполяризации мембраны блок снимается и в клетку входит Na, K и Са. v Са способен изменять свойства нейронов. Избыток Са токсичен для нейронов.
 – активируемые глутаматом ионотропные рецепторы, опосредуют") v AMPA-рецепторы (Рецепторы альфаамино-3 -гидрокси-5 -метил-4 изоксазолпропионовой кислоты) – активируемые глутаматом ионотропные рецепторы, опосредуют быструю возбуждающую синаптическую трансмиссию, участвуют в обучении и памяти. Активируют Na и Кканалы.
v AMPA-рецепторы (Рецепторы альфаамино-3 -гидрокси-5 -метил-4 изоксазолпропионовой кислоты) – активируемые глутаматом ионотропные рецепторы, опосредуют быструю возбуждающую синаптическую трансмиссию, участвуют в обучении и памяти. Активируют Na и Кканалы.
 v Каинатные рецепторы активируют вход Na и К. v m. Glu. R (метаботропный рецептор глутамата) – активирует фосфолипазу С и ингибирует АЦ. Считается, что они вносят вклад в отсроченный нейрональный ответ и синаптическую пластичность.
v Каинатные рецепторы активируют вход Na и К. v m. Glu. R (метаботропный рецептор глутамата) – активирует фосфолипазу С и ингибирует АЦ. Считается, что они вносят вклад в отсроченный нейрональный ответ и синаптическую пластичность.
 v Алкоголь действует как слабый антагонист глутаматных рецепторов во время начального приёма алкоголя. v Однако хронический приём алкоголя может приводить к увеличению числа глутаматных рецепторов.
v Алкоголь действует как слабый антагонист глутаматных рецепторов во время начального приёма алкоголя. v Однако хронический приём алкоголя может приводить к увеличению числа глутаматных рецепторов.
 v Увеличение глутаматных рецепторов может привести к гипервозбудимости нейронов. v Эта гипервозбудимость проявляется в отсутствии приёма алкоголя и может быть критическим компонентом в понимании нейробиологии синдрома отнятия.
v Увеличение глутаматных рецепторов может привести к гипервозбудимости нейронов. v Эта гипервозбудимость проявляется в отсутствии приёма алкоголя и может быть критическим компонентом в понимании нейробиологии синдрома отнятия.
 Биологические эффекты глутамата: v Помимо быстрого возбуждающего эффекта на постсинаптическую мембрану, глутамат может вызывать длительные изменения нейрональной возбудимости, зависимые от активности нейрона (долговременная потенциация). v Глутамат обладает нейроэндокринными регуляторными функциями, влияя на секрецию гипофизарных гормонов.
Биологические эффекты глутамата: v Помимо быстрого возбуждающего эффекта на постсинаптическую мембрану, глутамат может вызывать длительные изменения нейрональной возбудимости, зависимые от активности нейрона (долговременная потенциация). v Глутамат обладает нейроэндокринными регуляторными функциями, влияя на секрецию гипофизарных гормонов.
 v Предполагается, что активация глутаматных рецепторов принимает участие в миграции нейронов в некоторых областях мозга в период развития. v Глутамат оказывает положительные эффекты на выживание, рост и развитие нейронов в мозжечке и может стимулировать рост дендритов пирамидальных клеток в гиппокампе.
v Предполагается, что активация глутаматных рецепторов принимает участие в миграции нейронов в некоторых областях мозга в период развития. v Глутамат оказывает положительные эффекты на выживание, рост и развитие нейронов в мозжечке и может стимулировать рост дендритов пирамидальных клеток в гиппокампе.
 v Уникальная способность мозга заключается в переводе временного опыта в бесчисленное количество воспоминаний, приписываемое длительной синаптической активности. v В ЦНС долговременные синаптические изменения по типу потенциации наиболее хорошо изучены в гиппокампе. Известно, что именно NMDA-рецепторы играют важную роль в запуске долговременной потенциации.
v Уникальная способность мозга заключается в переводе временного опыта в бесчисленное количество воспоминаний, приписываемое длительной синаптической активности. v В ЦНС долговременные синаптические изменения по типу потенциации наиболее хорошо изучены в гиппокампе. Известно, что именно NMDA-рецепторы играют важную роль в запуске долговременной потенциации.
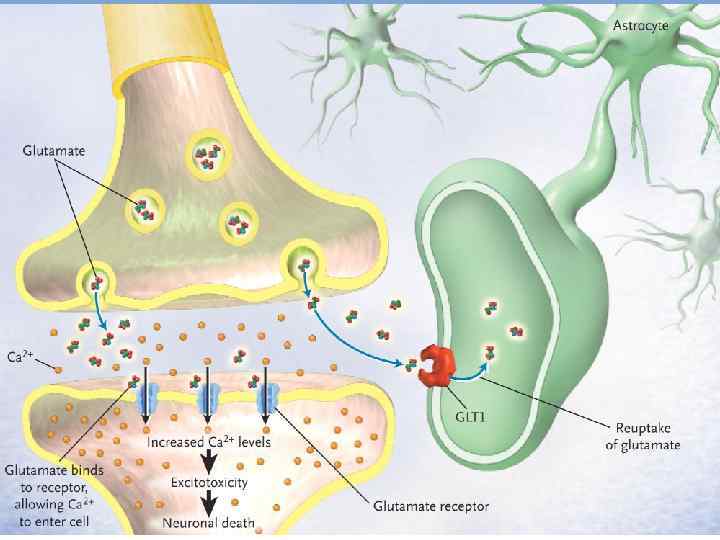
 v Глутамат предстаёт двуликим Янусом. С одной стороны, он оказывает благотворные") Токсичность возбуждения (excitotoxicity) v Глутамат предстаёт двуликим Янусом. С одной стороны, он оказывает благотворные эффекты на регуляцию роста нейронов и их дифференцировку, с другой стороны может быть опасен для мозговой ткани. v Большие количества глутамата могут освобождаться при ишемии, гипогликемии, эпилептических судорогах и нейродегенеративных заболеваниях (болезнь Альцгеймера, паркинсонизм и БАС)
Токсичность возбуждения (excitotoxicity) v Глутамат предстаёт двуликим Янусом. С одной стороны, он оказывает благотворные эффекты на регуляцию роста нейронов и их дифференцировку, с другой стороны может быть опасен для мозговой ткани. v Большие количества глутамата могут освобождаться при ишемии, гипогликемии, эпилептических судорогах и нейродегенеративных заболеваниях (болезнь Альцгеймера, паркинсонизм и БАС)
 v Вход большого количества Са приводит к нарушениям протеинов цитоскелета и активации протеаз и фосфолипаз, что приводит также к образованию свободных радикалов, повреждающих клетки. v Показана нейропротективная роль NMDA-антагонистов при ишемии головного мозга, что может быть полезно при ишемических импульсах, травматических повреждениях и эпилепсии.
v Вход большого количества Са приводит к нарушениям протеинов цитоскелета и активации протеаз и фосфолипаз, что приводит также к образованию свободных радикалов, повреждающих клетки. v Показана нейропротективная роль NMDA-антагонистов при ишемии головного мозга, что может быть полезно при ишемических импульсах, травматических повреждениях и эпилепсии.
 Влияние алкоголя на медиаторные системы
Влияние алкоголя на медиаторные системы
 Влияние на ГАМК-А v Влияние алкоголя на ГАМК-А обеспечивает не только уменьшение тревожности и седативногипнотическое действие, но и играет роль в моторной дискоординации. v Алкоголь вызывает изменения в некоторых субтипах ГАМК-Арецепторов. v Алкоголь также способен увеличивать высвобождение ГАМК в некоторых областях мозга.
Влияние на ГАМК-А v Влияние алкоголя на ГАМК-А обеспечивает не только уменьшение тревожности и седативногипнотическое действие, но и играет роль в моторной дискоординации. v Алкоголь вызывает изменения в некоторых субтипах ГАМК-Арецепторов. v Алкоголь также способен увеличивать высвобождение ГАМК в некоторых областях мозга.
 v Хронические эффекты алкоголя на ГАМК приводят к функциональной адаптации ГАМКергической системы, что приводит к гипервозбудимости, проявляющейся как тревожность или даже судороги во время абстинентного синдрома. v Бензодиазепины, модулирующие ГАМКергические функции, являются лекарствами выбора при алкогольной абстиненции.
v Хронические эффекты алкоголя на ГАМК приводят к функциональной адаптации ГАМКергической системы, что приводит к гипервозбудимости, проявляющейся как тревожность или даже судороги во время абстинентного синдрома. v Бензодиазепины, модулирующие ГАМКергические функции, являются лекарствами выбора при алкогольной абстиненции.
 Влияние на глутамат v Алкоголь быстро ингибирует глутаматергическую трансмиссию, оказывая анксиолитический эффект. v Нейрональная пластичность, опосредованная NMDA-рецепторами также ингибируется алкоголем. v Это действие может проявляться сильными амнестическими эффектами.
Влияние на глутамат v Алкоголь быстро ингибирует глутаматергическую трансмиссию, оказывая анксиолитический эффект. v Нейрональная пластичность, опосредованная NMDA-рецепторами также ингибируется алкоголем. v Это действие может проявляться сильными амнестическими эффектами.
 v Хроническое употребление алкоголя вызывает изменения в количестве и функции NMDA-рецепторов в качестве компенсации тормозящего действия. v При абстиненции система глутамата возбуждается, приводя к гипервозбудимости нейронов и нейрональному повреждению.
v Хроническое употребление алкоголя вызывает изменения в количестве и функции NMDA-рецепторов в качестве компенсации тормозящего действия. v При абстиненции система глутамата возбуждается, приводя к гипервозбудимости нейронов и нейрональному повреждению.
 Влияние на дофаминовую систему. v Алкоголь способствует усилению выделения дофамина в мезолимбической системе (от вентральной тегментальной области к прилежащему ядру – nucleus accumbens), обеспечивая стимуляцию моторики и позитивное подкрепление приёма алкоголя.
Влияние на дофаминовую систему. v Алкоголь способствует усилению выделения дофамина в мезолимбической системе (от вентральной тегментальной области к прилежащему ядру – nucleus accumbens), обеспечивая стимуляцию моторики и позитивное подкрепление приёма алкоголя.
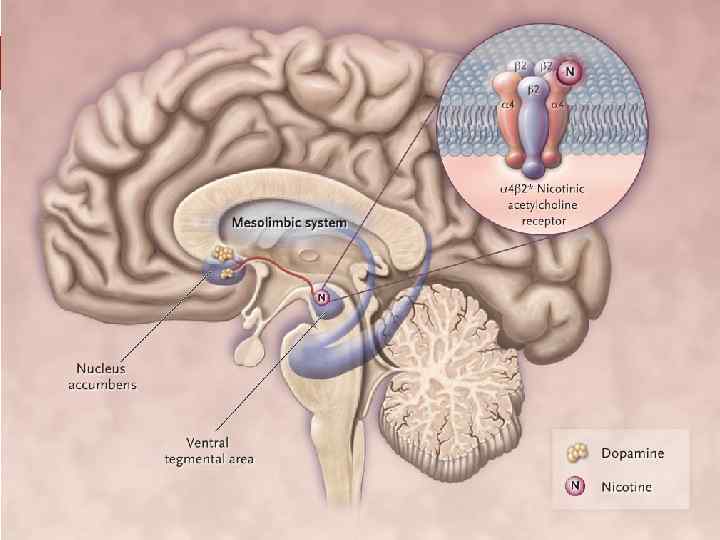
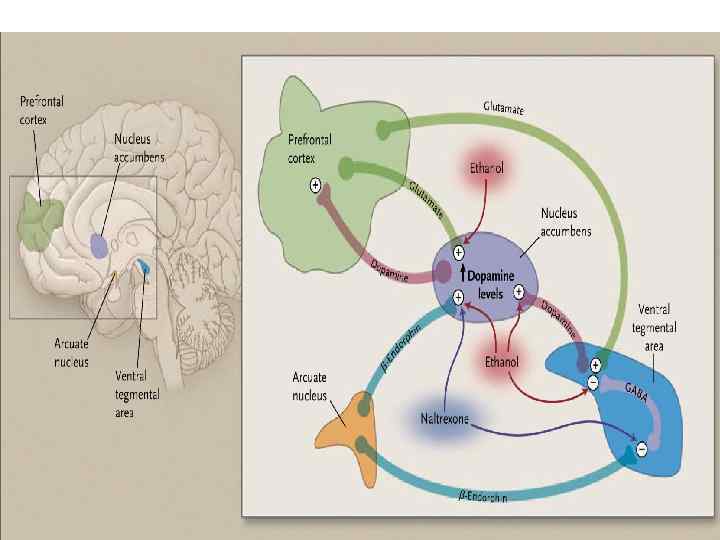
 v Хотя влияние на дофаминергическую систему не является критической, так как удаление мезолимбической системы значительно не изменяет самоназначение алкоголя в исследованиях на животных.
v Хотя влияние на дофаминергическую систему не является критической, так как удаление мезолимбической системы значительно не изменяет самоназначение алкоголя в исследованиях на животных.
 Влияние на серотониновую систему v Усиление влияния серотонина на 5 НТ 3 -рецепторы вносит вклад в позитивное подкрепление приёма алкоголя и его стимулирующие эффекты, так как плотно расположенные 5 -НТ-3 -рецепторы усиливают высвобождение дофамина в мезолимбической системе.
Влияние на серотониновую систему v Усиление влияния серотонина на 5 НТ 3 -рецепторы вносит вклад в позитивное подкрепление приёма алкоголя и его стимулирующие эффекты, так как плотно расположенные 5 -НТ-3 -рецепторы усиливают высвобождение дофамина в мезолимбической системе.
 v Блокада 5 -НТ-3 рецепторов вызывает уменьшение выделения дофамина, вызванное алкоголем, также как и уменьшение приёма алкоголя у человека.
v Блокада 5 -НТ-3 рецепторов вызывает уменьшение выделения дофамина, вызванное алкоголем, также как и уменьшение приёма алкоголя у человека.
 Влияние на β-эндорфин v Эндорфин опосредует позитивные гедонические эффекты алкоголя и вносит вклад в его подкрепляющие свойства, посредством стимуляции выделения дофамина в прилежащем ядре. v Антагонисты опиатных рецепторов уменьшают выделение дофамина, вызванное алкоголем, а следовательно и потребление алкоголя у животных и людей.
Влияние на β-эндорфин v Эндорфин опосредует позитивные гедонические эффекты алкоголя и вносит вклад в его подкрепляющие свойства, посредством стимуляции выделения дофамина в прилежащем ядре. v Антагонисты опиатных рецепторов уменьшают выделение дофамина, вызванное алкоголем, а следовательно и потребление алкоголя у животных и людей.