Нейрофизиология_НЕЙРОННЫЕ МЕХАНИЗМЫ.pptx
- Количество слайдов: 52
 нейрофизиология
нейрофизиология
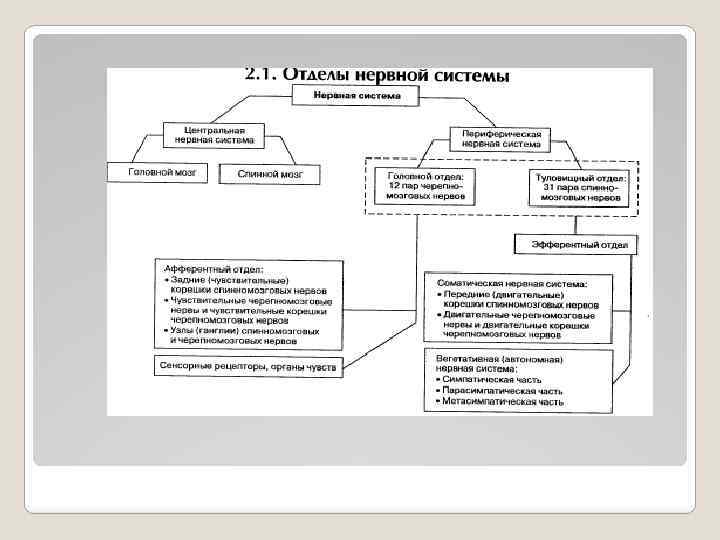
 Головной мозг
Головной мозг
 нейрон
нейрон
 — математические модели, а также") • Иску сственные нейро нные се ти (ИНС) — математические модели, а также их программные или аппаратные реализации, построенные по принципу организации и функционирования биологических нейронных сетей — сетей нервных клеток живого организма: 1. 2. 3. 4. 5. 6. 7. Основными действующими элементами нервных сетей являются отдельные клетки - нейроны. Активность нейронов при передаче и обработке нервных импульсов регулируется свойствами мембраны, которые могут меняться под воздействием синаптических медиаторов. Биологические функции нейрона могут меняться и адаптироваться к условиям функционирования. Нейроны объединяются в нейронные сети, основные типы которых, а также схемы проводящих путей мозга являются генетически запрограммированными. В процессе развития возможно локальное видоизменение нейронных сетей с формированием новых соединений между нейронами. Нейронные сети взаимодействуют с клетками других типов. Нейронные сети могут формировать функциональные системы.
• Иску сственные нейро нные се ти (ИНС) — математические модели, а также их программные или аппаратные реализации, построенные по принципу организации и функционирования биологических нейронных сетей — сетей нервных клеток живого организма: 1. 2. 3. 4. 5. 6. 7. Основными действующими элементами нервных сетей являются отдельные клетки - нейроны. Активность нейронов при передаче и обработке нервных импульсов регулируется свойствами мембраны, которые могут меняться под воздействием синаптических медиаторов. Биологические функции нейрона могут меняться и адаптироваться к условиям функционирования. Нейроны объединяются в нейронные сети, основные типы которых, а также схемы проводящих путей мозга являются генетически запрограммированными. В процессе развития возможно локальное видоизменение нейронных сетей с формированием новых соединений между нейронами. Нейронные сети взаимодействуют с клетками других типов. Нейронные сети могут формировать функциональные системы.
 Первый тип составляют иерархические сети, часто встречающиеся в сенсорных и двигательных путях. Информация в таких сетях передается в процессе последовательного перехода от одного уровня иерархии к другому. Нейроны образуют два характерных типа соединений - конвергентные, когда большое число нейронов одного уровня контактирует с меньшим числом нейронов следующего уровня, и дивергентные, в которых контакты устанавливаются со все большим числом клеток последующих слоев иерархии. Сочетание конвергентных и дивергентных соединений обеспечивает многократное дублирование информационных путей, что является решающим фактором надежности нейронной сети. При гибели части клеток, сохранившиеся нейроны оказываются в состоянии поддерживать функционирование сети. Ко второму типу нейронных сетей относятся локальные сети, формируемые нейронами с ограниченными сферами влияния. Нейроны локальных сетей производят переработку информации в пределах одного уровня иерархии. Выделяют несколько основных типов нейронных сетей, отличающихся структурой и назначением:
Первый тип составляют иерархические сети, часто встречающиеся в сенсорных и двигательных путях. Информация в таких сетях передается в процессе последовательного перехода от одного уровня иерархии к другому. Нейроны образуют два характерных типа соединений - конвергентные, когда большое число нейронов одного уровня контактирует с меньшим числом нейронов следующего уровня, и дивергентные, в которых контакты устанавливаются со все большим числом клеток последующих слоев иерархии. Сочетание конвергентных и дивергентных соединений обеспечивает многократное дублирование информационных путей, что является решающим фактором надежности нейронной сети. При гибели части клеток, сохранившиеся нейроны оказываются в состоянии поддерживать функционирование сети. Ко второму типу нейронных сетей относятся локальные сети, формируемые нейронами с ограниченными сферами влияния. Нейроны локальных сетей производят переработку информации в пределах одного уровня иерархии. Выделяют несколько основных типов нейронных сетей, отличающихся структурой и назначением:
 Нервная система содержит помимо нейронов клетки других типов Астроцитарная сеть Нейроглиальная сеть «Трехчастная» модель синапса…+ элементы матрикса = «четырехчастная» модель синапс
Нервная система содержит помимо нейронов клетки других типов Астроцитарная сеть Нейроглиальная сеть «Трехчастная» модель синапса…+ элементы матрикса = «четырехчастная» модель синапс
 Биофизические. Основаны на классическом формализме Ходжкина-Хаксли, описывают динамические режимы нейронов и синапсов, позволяет верифицировать результаты моделирования с экспериментальными данными клеточной нейрофизиологии; Феноменологические (интегратор, генератор Фитц. Хью Нагумо и др. ). Описывают функциональные особенности динамики (пороговая генерация импульса, рефрактерный период и др. ). Возможность реализации в виде электронных устройств обработки информации. Биологические. Гибридный (аниматный) подход. Биофизическая детализация (молекулярные механизмы синаптической передачи и пластичности) + мониторинг и управление (обучение, память, контроль через нейроэлектронный интерфейс) СЕТЕВЫЕ МОДЕЛИ
Биофизические. Основаны на классическом формализме Ходжкина-Хаксли, описывают динамические режимы нейронов и синапсов, позволяет верифицировать результаты моделирования с экспериментальными данными клеточной нейрофизиологии; Феноменологические (интегратор, генератор Фитц. Хью Нагумо и др. ). Описывают функциональные особенности динамики (пороговая генерация импульса, рефрактерный период и др. ). Возможность реализации в виде электронных устройств обработки информации. Биологические. Гибридный (аниматный) подход. Биофизическая детализация (молекулярные механизмы синаптической передачи и пластичности) + мониторинг и управление (обучение, память, контроль через нейроэлектронный интерфейс) СЕТЕВЫЕ МОДЕЛИ
 нейрорецепция
нейрорецепция
 синапсы
синапсы
 Внешняя мембрана нейронов чувствительна к действию специальных веществ, которые выделяются из пресинаптической терминали – к нейромедиаторам. В настоящее время идентифицировано около 100 веществ, которые выполняют эту функцию. На внешней стороне мембраны расположены специализированные белковые молекулы – рецепторы, которые и взаимодействуют с нейромедиатором нейромедиация
Внешняя мембрана нейронов чувствительна к действию специальных веществ, которые выделяются из пресинаптической терминали – к нейромедиаторам. В настоящее время идентифицировано около 100 веществ, которые выполняют эту функцию. На внешней стороне мембраны расположены специализированные белковые молекулы – рецепторы, которые и взаимодействуют с нейромедиатором нейромедиация
 В результате происходит открытие каналов специфической ионной проницаемости – только определенные ионы могут массированно проходить в клетку после действия медиатора. Развивается локальная деполяризация или гиперполяризация мембраны, которая называется постсинаптическим потенциалом (ПСП). ПСП могут быть возбудительными (ВПСП) и тормозными (ТПСП). Амплитуда ПСП может достигать 20 м. В.
В результате происходит открытие каналов специфической ионной проницаемости – только определенные ионы могут массированно проходить в клетку после действия медиатора. Развивается локальная деполяризация или гиперполяризация мембраны, которая называется постсинаптическим потенциалом (ПСП). ПСП могут быть возбудительными (ВПСП) и тормозными (ТПСП). Амплитуда ПСП может достигать 20 м. В.
 нейромедиация
нейромедиация
 нейромедиация
нейромедиация
 нейромедиация
нейромедиация
 нейромедиация
нейромедиация
 Классификация рецепторов основывается, в первую очередь, на характере ощущений, возникающих у человека при их раздражении. Различают зрительные, слуховые, обонятельные, вкусовые, осязательные рецепторы, терморецепторы, проприои вестибулорецепторы (рецепторы положения тела и его частей в пространстве). Обсуждается вопрос существования специальных рецепторов боли.
Классификация рецепторов основывается, в первую очередь, на характере ощущений, возникающих у человека при их раздражении. Различают зрительные, слуховые, обонятельные, вкусовые, осязательные рецепторы, терморецепторы, проприои вестибулорецепторы (рецепторы положения тела и его частей в пространстве). Обсуждается вопрос существования специальных рецепторов боли.
 Рецепторы разделяют, кроме того, на внешние, или экстерорецепторы, и внутренние, или интерорецепторы. К экстерорецепторам относятся слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы. К интерорецепторам относятся вестибулорецепторы и проприорецепторы (рецепторы опорно-двигательного аппарата), а также интерорецепторы, сигнализирующие о состоянии внутренних органов
Рецепторы разделяют, кроме того, на внешние, или экстерорецепторы, и внутренние, или интерорецепторы. К экстерорецепторам относятся слуховые, зрительные, обонятельные, вкусовые и осязательные рецепторы. К интерорецепторам относятся вестибулорецепторы и проприорецепторы (рецепторы опорно-двигательного аппарата), а также интерорецепторы, сигнализирующие о состоянии внутренних органов
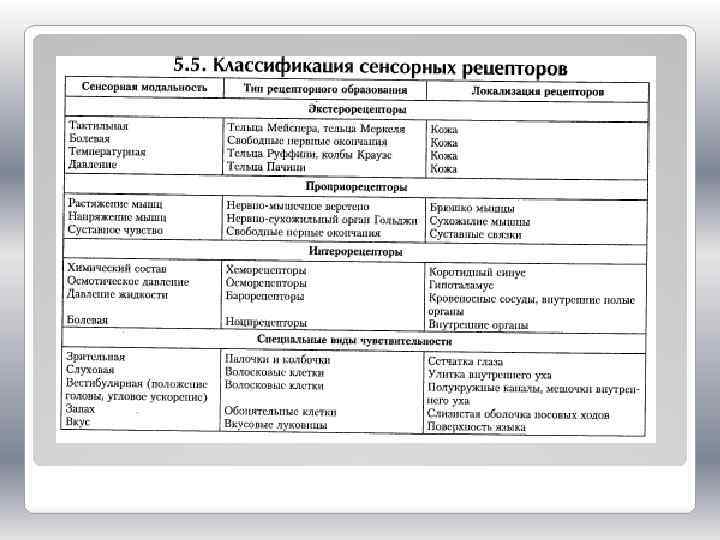
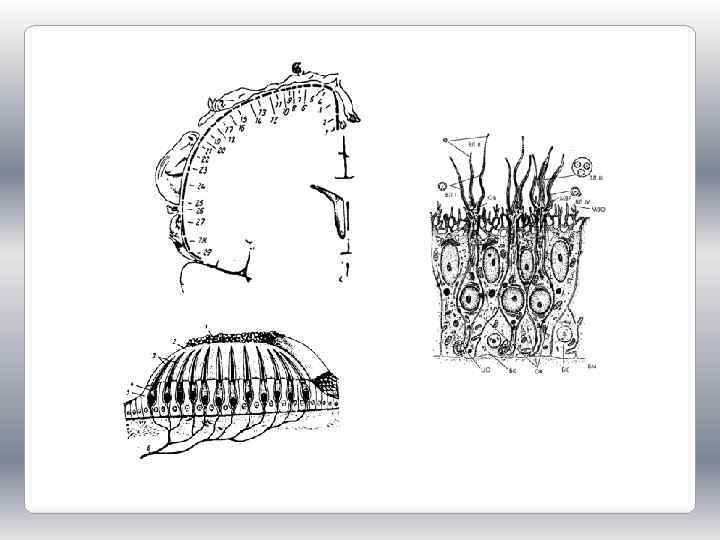
 В зависимости от природы раздражителя, на который они оптимально настроены, рецепторы можно классифицировать следующим образом: 1) фоторецепторы; 2) механорецепторы, к которым относятся рецепторы слуховые, вестибулярные, тактильные рецепторы кожи, рецепторы опорно-двигательного аппарата, барорецепторы сердечнососудистой системы; 3) хеморецепторы, включающие рецепторы вкуса и обоняния, сосудистые и тканевые рецепторы; 4) терморецепторы (кожи и внутренних органов, а также центральные термочувствительные нейроны) и 5) болевые (ноцицептивные) рецепторы.
В зависимости от природы раздражителя, на который они оптимально настроены, рецепторы можно классифицировать следующим образом: 1) фоторецепторы; 2) механорецепторы, к которым относятся рецепторы слуховые, вестибулярные, тактильные рецепторы кожи, рецепторы опорно-двигательного аппарата, барорецепторы сердечнососудистой системы; 3) хеморецепторы, включающие рецепторы вкуса и обоняния, сосудистые и тканевые рецепторы; 4) терморецепторы (кожи и внутренних органов, а также центральные термочувствительные нейроны) и 5) болевые (ноцицептивные) рецепторы.
 Абсолютная чувствительность сенсорной системы основана на ее свойстве обнаруживать слабые, короткие или маленькие по размеру раздражители. Абсолютную чувствительность измеряют порогом той или иной реакции организма на сенсорное воздействие. Обычно принимают за пороговую такую силу стимула, вероятность восприятия которого равна 0, 5– 0, 75 Сенсорные пороги
Абсолютная чувствительность сенсорной системы основана на ее свойстве обнаруживать слабые, короткие или маленькие по размеру раздражители. Абсолютную чувствительность измеряют порогом той или иной реакции организма на сенсорное воздействие. Обычно принимают за пороговую такую силу стимула, вероятность восприятия которого равна 0, 5– 0, 75 Сенсорные пороги
 Абсолютная чувствительность
Абсолютная чувствительность
 Дифференциальная сенсорная чувствительность основана на способности сенсорной системы к различению сигналов. Важная характеристика каждой сенсорной системы – способность замечать различия в свойствах одновременно или последовательно действующих раздражителей. ДИФФЕРЕНЦИАЛЬНАЯ СЕНСОРНАЯ ЧУВСТВИТЕЛЬНОСТЬ
Дифференциальная сенсорная чувствительность основана на способности сенсорной системы к различению сигналов. Важная характеристика каждой сенсорной системы – способность замечать различия в свойствах одновременно или последовательно действующих раздражителей. ДИФФЕРЕНЦИАЛЬНАЯ СЕНСОРНАЯ ЧУВСТВИТЕЛЬНОСТЬ
 выражается следующей формулой: Е =") Зависимость силы ощущения от силы раздражения (закон Вебера–Фехнера) выражается следующей формулой: Е = a log I + b где Е – величина ощущения, I – сила раздражения, а и b – константы, различные для разных модальностей стимулов. Эта формула показывает, что ощущение усиливается пропорционально логарифму интенсивности раздражения.
Зависимость силы ощущения от силы раздражения (закон Вебера–Фехнера) выражается следующей формулой: Е = a log I + b где Е – величина ощущения, I – сила раздражения, а и b – константы, различные для разных модальностей стимулов. Эта формула показывает, что ощущение усиливается пропорционально логарифму интенсивности раздражения.
 В сенсорной системе сигналы кодируются двоичным кодом, т. е. наличием или отсутствием электрического импульса в тот или иной момент времени [Сомьен, 1975]. Такой способ кодирования крайне прост и устойчив к помехам. Информация о раздражении и его параметрах передается в виде отдельных импульсов, а также групп, или «пачек» импульсов КОДИРОВАНИЕ ИНФОРМАЦИИ
В сенсорной системе сигналы кодируются двоичным кодом, т. е. наличием или отсутствием электрического импульса в тот или иной момент времени [Сомьен, 1975]. Такой способ кодирования крайне прост и устойчив к помехам. Информация о раздражении и его параметрах передается в виде отдельных импульсов, а также групп, или «пачек» импульсов КОДИРОВАНИЕ ИНФОРМАЦИИ
 Амплитуда, длительность и форма каждого импульса одинаковы, но количество импульсов в пачке, частота их следования, длительность пачек и интервалов между ними, а также временной «рисунок» (pattern) пачки различны и зависят от характеристик стимула. Сенсорная информация кодируется также числом одновременно возбужденных нейронов и их расположением в нейронном слое.
Амплитуда, длительность и форма каждого импульса одинаковы, но количество импульсов в пачке, частота их следования, длительность пачек и интервалов между ними, а также временной «рисунок» (pattern) пачки различны и зависят от характеристик стимула. Сенсорная информация кодируется также числом одновременно возбужденных нейронов и их расположением в нейронном слое.
 В коре мозга сигналы кодируются также последовательностью включения параллельно работающих нейронных каналов, синхронностью ритмических импульсных разрядов возбужденных нейронов, изменением их числа. В коре одним из основных используемых способов становится позиционное кодирование. Оно заключается в том, что какой-то признак раздражителя вызывает возбуждение определенного нейрона или небольшой группы нейронов, расположенных в определенном месте нейронного слоя
В коре мозга сигналы кодируются также последовательностью включения параллельно работающих нейронных каналов, синхронностью ритмических импульсных разрядов возбужденных нейронов, изменением их числа. В коре одним из основных используемых способов становится позиционное кодирование. Оно заключается в том, что какой-то признак раздражителя вызывает возбуждение определенного нейрона или небольшой группы нейронов, расположенных в определенном месте нейронного слоя
 Например, возбуждение небольшой локальной группы нейронов зрительной коры означает, что в определенной части поля зрения появилась световая полоска определенного размера и ориентации. Возбуждение определенных нейронов височной коры сигнализирует о появлении в поле зрения знакомого лица. Для периферических отделов сенсорной системы типично временное кодирование признаков раздражителя, а на высших уровнях происходит переход к преимущественно пространственному (в основном позиционному) коду.
Например, возбуждение небольшой локальной группы нейронов зрительной коры означает, что в определенной части поля зрения появилась световая полоска определенного размера и ориентации. Возбуждение определенных нейронов височной коры сигнализирует о появлении в поле зрения знакомого лица. Для периферических отделов сенсорной системы типично временное кодирование признаков раздражителя, а на высших уровнях происходит переход к преимущественно пространственному (в основном позиционному) коду.
 Сведения, накопленные за последние десятилетия о нейронах сенсорных систем, подтверждают детекторный принцип нейронной организации самых разных анализаторов. Для зрительной коры были описаны нейроныдетекторы, избирательно отвечающие на элементы фигуры, контура — линии, полосы, углы Американские ученые Д. Хьюбел и Т. Визель разработали классификацию нейронов-детекторов зрительной коры, селективно чувствительных к различной ориентации линий и их размеру, связав их с простыми, сложными и сверхсложными рецептивными полями. НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
Сведения, накопленные за последние десятилетия о нейронах сенсорных систем, подтверждают детекторный принцип нейронной организации самых разных анализаторов. Для зрительной коры были описаны нейроныдетекторы, избирательно отвечающие на элементы фигуры, контура — линии, полосы, углы Американские ученые Д. Хьюбел и Т. Визель разработали классификацию нейронов-детекторов зрительной коры, селективно чувствительных к различной ориентации линий и их размеру, связав их с простыми, сложными и сверхсложными рецептивными полями. НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
 Важным шагом в развитии теории сенсорных систем явилось открытие константных нейроновдетекторов, учитывающих, кроме зрительных сигналов, сигналы о положении глаз в орбитах. В теменной коре реакция константных нейроновдетекторов привязана к определенной области внешнего пространства, образуя константный экран. Другой тип константных нейронов-детекторов, кодирующих цвет, открыт С. Зеки в экстрастриарной зрительной коре, в поле V 4. Их реакция на определенные отражательные свойства цветовой поверхности объекта не зависит от условий освещения. НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
Важным шагом в развитии теории сенсорных систем явилось открытие константных нейроновдетекторов, учитывающих, кроме зрительных сигналов, сигналы о положении глаз в орбитах. В теменной коре реакция константных нейроновдетекторов привязана к определенной области внешнего пространства, образуя константный экран. Другой тип константных нейронов-детекторов, кодирующих цвет, открыт С. Зеки в экстрастриарной зрительной коре, в поле V 4. Их реакция на определенные отражательные свойства цветовой поверхности объекта не зависит от условий освещения. НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
 Это конечная и наиболее сложная операция сенсорной системы. Она заключается в отнесении образа к тому или иному классу объектов, с которыми ранее встречался организм, т. е. в классификации образов. Синтезируя сигналы от нейроновдетекторов, высший отдел сенсорной системы формирует «образ» раздражителя и сравнивает его со множеством образов, хранящихся в памяти. ОПОЗНАНИЕ ОБРАЗОВ
Это конечная и наиболее сложная операция сенсорной системы. Она заключается в отнесении образа к тому или иному классу объектов, с которыми ранее встречался организм, т. е. в классификации образов. Синтезируя сигналы от нейроновдетекторов, высший отдел сенсорной системы формирует «образ» раздражителя и сравнивает его со множеством образов, хранящихся в памяти. ОПОЗНАНИЕ ОБРАЗОВ
 Переработка информации в сенсорной системе осуществляется с помощью процессов возбудительного и тормозного межнейронного взаимодействия. Это взаимодействие осуществляется по горизонтали, т. е. в пределах одного нейронного слоя, и по вертикали, т. е. между нейронами соседних слоев. МЕХАНИЗМЫ ПЕРЕРАБОТКИ ИНФОРМАЦИИ В СЕНСОРНОЙ СИСТЕМЕ
Переработка информации в сенсорной системе осуществляется с помощью процессов возбудительного и тормозного межнейронного взаимодействия. Это взаимодействие осуществляется по горизонтали, т. е. в пределах одного нейронного слоя, и по вертикали, т. е. между нейронами соседних слоев. МЕХАНИЗМЫ ПЕРЕРАБОТКИ ИНФОРМАЦИИ В СЕНСОРНОЙ СИСТЕМЕ
 Возбудительное взаимодействие по вертикали заключается в том, что аксон каждого нейрона, приходя в вышележащий слой, контактирует с несколькими нейронами, каждый из которых получает сигналы от нескольких клеток предыдущего слоя. В результате подобного взаимодействия формируются так называемые рецептивные и проекционные поля сенсорных нейронов, играющие ключевую роль в переработке сенсорных сигналов.
Возбудительное взаимодействие по вертикали заключается в том, что аксон каждого нейрона, приходя в вышележащий слой, контактирует с несколькими нейронами, каждый из которых получает сигналы от нескольких клеток предыдущего слоя. В результате подобного взаимодействия формируются так называемые рецептивные и проекционные поля сенсорных нейронов, играющие ключевую роль в переработке сенсорных сигналов.
 Совокупность рецепторов, сигналы с которых поступают на данный нейрон, называют его рецептивным полем. В пределах рецептивного поля происходит пространственная суммация, т. е. ответ нейрона увеличивается, а порог его реакции снижается при увеличении площади светового пятна или участка стимулируемой кожной поверхности.
Совокупность рецепторов, сигналы с которых поступают на данный нейрон, называют его рецептивным полем. В пределах рецептивного поля происходит пространственная суммация, т. е. ответ нейрона увеличивается, а порог его реакции снижается при увеличении площади светового пятна или участка стимулируемой кожной поверхности.
 Проекционным полем сенсорного нейрона называют совокупность нейронов более высокого слоя, которые получают его сигналы. Наличие у нейронов проекционных полей обеспечивает сенсорной системе высокую устойчивость к повреждающим воздействиям и способность к восстановлению функций, нарушенных патологическим процессом.
Проекционным полем сенсорного нейрона называют совокупность нейронов более высокого слоя, которые получают его сигналы. Наличие у нейронов проекционных полей обеспечивает сенсорной системе высокую устойчивость к повреждающим воздействиям и способность к восстановлению функций, нарушенных патологическим процессом.
 и проекционного (б ) полей сенсорного нейрона и нервной") Схема рецептивного ( а ) и проекционного (б ) полей сенсорного нейрона и нервной сети ( в ).
Схема рецептивного ( а ) и проекционного (б ) полей сенсорного нейрона и нервной сети ( в ).
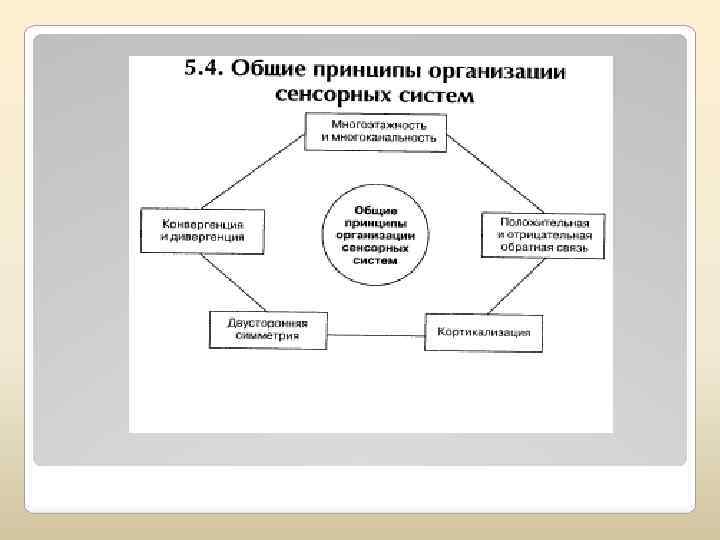
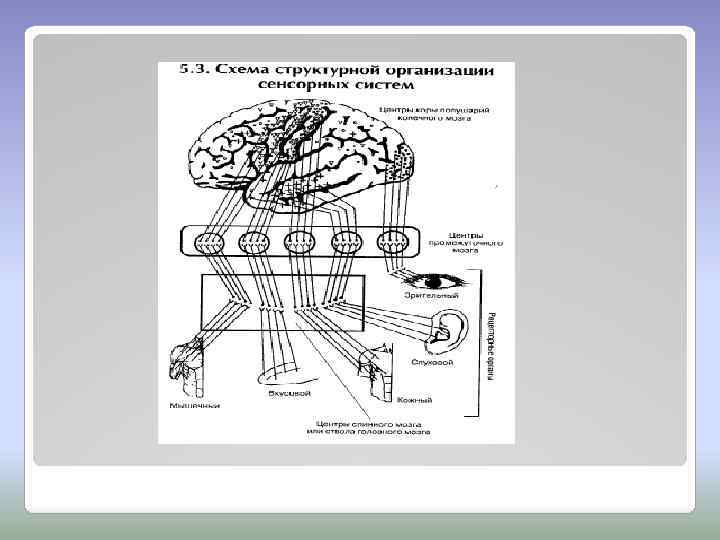
 Все сенсорные системы человека организованы по некоторым общим принципам Важнейшие из них следующие: многослойность, многоканальность, наличие так называемых «сенсорных воронок» , а также дифференциация систем по вертикали и по горизонтали. Многослойность сводится к наличию в каждой системе Многоканальность сенсорной системы заключается в том, что в каждом нейронном слое имеется множество (от десятков тысяч до миллионов) нервных клеток, связанных нервными волокнами со множеством клеток следующего слоя нескольких слоев нейронов, первый из которых связан с рецепторами, а последний – с нейронами моторных областей коры мозга. ОБЩИЕ СВОЙСТВА СЕНСОРНЫХ СИСТЕМ
Все сенсорные системы человека организованы по некоторым общим принципам Важнейшие из них следующие: многослойность, многоканальность, наличие так называемых «сенсорных воронок» , а также дифференциация систем по вертикали и по горизонтали. Многослойность сводится к наличию в каждой системе Многоканальность сенсорной системы заключается в том, что в каждом нейронном слое имеется множество (от десятков тысяч до миллионов) нервных клеток, связанных нервными волокнами со множеством клеток следующего слоя нескольких слоев нейронов, первый из которых связан с рецепторами, а последний – с нейронами моторных областей коры мозга. ОБЩИЕ СВОЙСТВА СЕНСОРНЫХ СИСТЕМ
 Изучение вертикальных и горизонтальных связей нейронов-детекторов различного типа привело к открытию общих принципов нейронной архитектуры коры. В. Маунткасл (V. Mountcastle) — ученый из медицинской школы Университета Джонса Гопкинса — в 60 -х годах впервые описал вертикальный принцип организации коры больших полушарий. НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
Изучение вертикальных и горизонтальных связей нейронов-детекторов различного типа привело к открытию общих принципов нейронной архитектуры коры. В. Маунткасл (V. Mountcastle) — ученый из медицинской школы Университета Джонса Гопкинса — в 60 -х годах впервые описал вертикальный принцип организации коры больших полушарий. НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
 Нейроны соматосенсорной по модальности сгруппированы в вертикальные колонки. Колонки имеют вид трехмерных прямоугольных блоков разной величины и проходят через все клеточные слои коры. Со стороны поверхности коры они выглядят как пластины размером от 2050 мкм до 0, 25 -0, 5 мм. НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
Нейроны соматосенсорной по модальности сгруппированы в вертикальные колонки. Колонки имеют вид трехмерных прямоугольных блоков разной величины и проходят через все клеточные слои коры. Со стороны поверхности коры они выглядят как пластины размером от 2050 мкм до 0, 25 -0, 5 мм. НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
 В. Маунткасл формулируя основные положения своей теории о колончатой организации коры, отмечал, что «основной единицей активности в новой коре служит вертикально расположенная группа клеток с множеством связей между этими клетками по вертикальной оси и малым их числом в горизонтальном направлении» . НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
В. Маунткасл формулируя основные положения своей теории о колончатой организации коры, отмечал, что «основной единицей активности в новой коре служит вертикально расположенная группа клеток с множеством связей между этими клетками по вертикальной оси и малым их числом в горизонтальном направлении» . НЕЙРОННЫЕ МЕХАНИЗМЫ ВОСПРИЯТИЯ
 Применение микроэлектродной техники позволило выполнить тонкие измерения, характеризующие основные электрофизиологические характеристики нервных клеток Измерения показали, что каждая нервная клетка имеет отрицательный заряд, величина которого равна (40) – (-65) м. В. Нейронная активность
Применение микроэлектродной техники позволило выполнить тонкие измерения, характеризующие основные электрофизиологические характеристики нервных клеток Измерения показали, что каждая нервная клетка имеет отрицательный заряд, величина которого равна (40) – (-65) м. В. Нейронная активность
 Главное отличие нервной клетки от любой другой заключается в том, что она способна быстро изменять величину заряда вплоть до противоположного. Критический уровень деполяризации нейрона, при достижении которого возникает быстрый разряд, называется порогом генерации потенциала действия (ПД). Длительность потенциала действия различна у позвоночных и беспозвоночных животных – у беспозвоночных она равна 0, 1 мс, а у позвоночных 1– 2 мс. Серия потенциалов действия, распределенных во времени, является основой для пространственно-временного кодирования. Нейронная активность
Главное отличие нервной клетки от любой другой заключается в том, что она способна быстро изменять величину заряда вплоть до противоположного. Критический уровень деполяризации нейрона, при достижении которого возникает быстрый разряд, называется порогом генерации потенциала действия (ПД). Длительность потенциала действия различна у позвоночных и беспозвоночных животных – у беспозвоночных она равна 0, 1 мс, а у позвоночных 1– 2 мс. Серия потенциалов действия, распределенных во времени, является основой для пространственно-временного кодирования. Нейронная активность
 Организм и среда
Организм и среда