lecture_8.ppt
- Количество слайдов: 63
 в лактозном опероне есть два «вспомогательных» оператора (О") Наряду с главным оператором (О 1) в лактозном опероне есть два «вспомогательных» оператора (О 2 и О 3). Уровень репрессии значительно снижается, если делетировать одновременно О 2 и О 3 тетрамер репрессора связывается одновременно с двумя участками узнавания на молекуле ДНК
Наряду с главным оператором (О 1) в лактозном опероне есть два «вспомогательных» оператора (О 2 и О 3). Уровень репрессии значительно снижается, если делетировать одновременно О 2 и О 3 тетрамер репрессора связывается одновременно с двумя участками узнавания на молекуле ДНК
 Репресор не препятствует связыванию РНК полимеразы с промотором Репрессия осуществляется на этапе перехода от открытого комплекса к элонгации
Репресор не препятствует связыванию РНК полимеразы с промотором Репрессия осуществляется на этапе перехода от открытого комплекса к элонгации
 лактозы нет O P R R лактоза алактоза А если есть и лактоза и глюкоза ?
лактозы нет O P R R лактоза алактоза А если есть и лактоза и глюкоза ?
 Почему lac-оперон не работает, когда в среде есть и глюкоза и лактоза? Катаболитная репрессия ? В действительности - активация CAP – Catabolite Activator Protein (активатор катаболитных оперонов) Работает в комплексе с цикло-АМФ, уровень которого существенно повышается при недостатке глюкозы CAP активирует целый ряд оперонов, обеспечивающих синтез ферментов, необходимых для использования альтернативных продуктов питания (gal, галактозный оперон; ara, арабинозный оперон)
Почему lac-оперон не работает, когда в среде есть и глюкоза и лактоза? Катаболитная репрессия ? В действительности - активация CAP – Catabolite Activator Protein (активатор катаболитных оперонов) Работает в комплексе с цикло-АМФ, уровень которого существенно повышается при недостатке глюкозы CAP активирует целый ряд оперонов, обеспечивающих синтез ферментов, необходимых для использования альтернативных продуктов питания (gal, галактозный оперон; ara, арабинозный оперон)
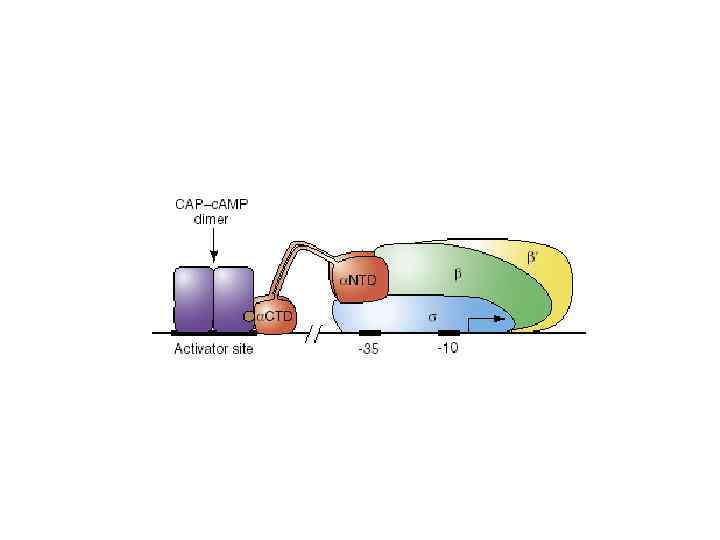
 CAPстимулирует образование открытого комплекса Участок связывания CAP находится перед промотором и содержит TGTGA последовательность Lac промотор и многие другие катаболитные промоторы являются очень слабыми. -35 бокс в этих промоторах сильно отличается от консенсуса Существуют мутанты laс промотора, которые обеспечивают транскрипцию без САP активатора
CAPстимулирует образование открытого комплекса Участок связывания CAP находится перед промотором и содержит TGTGA последовательность Lac промотор и многие другие катаболитные промоторы являются очень слабыми. -35 бокс в этих промоторах сильно отличается от консенсуса Существуют мутанты laс промотора, которые обеспечивают транскрипцию без САP активатора
 Анаболитные опероны Должны активироваться в условиях недостатка тех веществ, в синтезе которых участвуют кодируемые опероном ферменты триптофановый оперон: триптофан (корепрессор) связывается с неактивной формой репрессора, превращая ее в активную
Анаболитные опероны Должны активироваться в условиях недостатка тех веществ, в синтезе которых участвуют кодируемые опероном ферменты триптофановый оперон: триптофан (корепрессор) связывается с неактивной формой репрессора, превращая ее в активную
 обеспечивает дополнительную репрессию В начале оперона находится короткая ORF, кодирующая") Преждевременная остановка транскрипции (attenuation) обеспечивает дополнительную репрессию В начале оперона находится короткая ORF, кодирующая так называемый лидерный пептид После этой рамки считывания находится сигнал терминации транскрипции 90% транскрипционных комплексов диссоциируют
Преждевременная остановка транскрипции (attenuation) обеспечивает дополнительную репрессию В начале оперона находится короткая ORF, кодирующая так называемый лидерный пептид После этой рамки считывания находится сигнал терминации транскрипции 90% транскрипционных комплексов диссоциируют
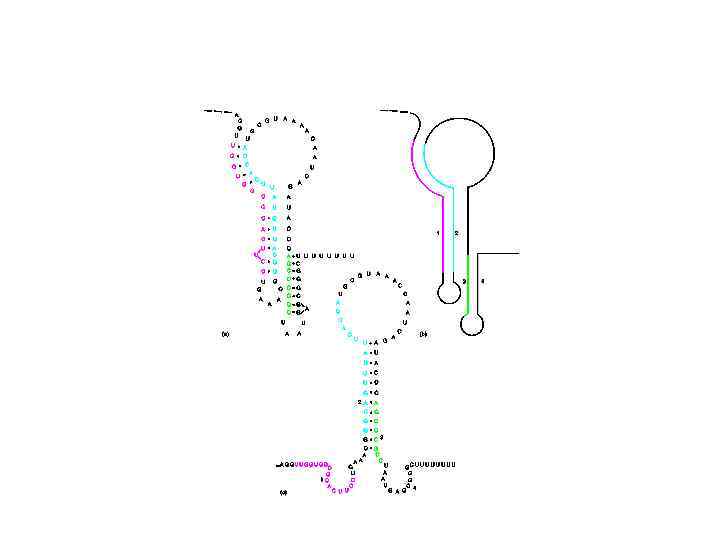
 У прокариот транскрипция и трансляция могут происходить одновременно При отсутствии триптофана рибосома задерживается на лидерной последовательности. Это смещает равновесие в сторону большой шпильки Регулируемые терминаторы транскрипции есть и в других анаболитных оперонах В гистидиновом опероне лидер содержит 7 кодонов гистидина
У прокариот транскрипция и трансляция могут происходить одновременно При отсутствии триптофана рибосома задерживается на лидерной последовательности. Это смещает равновесие в сторону большой шпильки Регулируемые терминаторы транскрипции есть и в других анаболитных оперонах В гистидиновом опероне лидер содержит 7 кодонов гистидина
 Транскрипция и") Транскрипция у эукариот Транскрипционный аппарат сосредоточен в ядре (есть еще митохондриальная полимераза) Транскрипция и трансляция пространственно разделены У эукариот существует несколько РНК полимераз (по меньшей мере три ядерных РНК полимеразы + митохондриальная)
Транскрипция у эукариот Транскрипционный аппарат сосредоточен в ядре (есть еще митохондриальная полимераза) Транскрипция и трансляция пространственно разделены У эукариот существует несколько РНК полимераз (по меньшей мере три ядерных РНК полимеразы + митохондриальная)
 Три типа эукариотических РНК-полимераз экстракт из ядер экстракт из ядрышек (названы в соответствии с порядком элюции с ионообменной колонки (DEAE сефадекса); по другой номенклатуре (П. Шамбон): А, В, С Тип Локализация субстрат Чувствительность к a-аманитину RNA Pol I Ядрышко Рибосомные гены (кроме 5 S р. РНК) Не чувствительна RNA Pol II Нуклеоплазма Все РНК, кодирующие белки, и некодирующие (регуляторные) РНК Чувствительна RNA Pol III Нуклеоплазма т. РНК, 5 S р. РНК, U 6 sn. РНК и ряд других малых РНК Относительно чувствительна
Три типа эукариотических РНК-полимераз экстракт из ядер экстракт из ядрышек (названы в соответствии с порядком элюции с ионообменной колонки (DEAE сефадекса); по другой номенклатуре (П. Шамбон): А, В, С Тип Локализация субстрат Чувствительность к a-аманитину RNA Pol I Ядрышко Рибосомные гены (кроме 5 S р. РНК) Не чувствительна RNA Pol II Нуклеоплазма Все РНК, кодирующие белки, и некодирующие (регуляторные) РНК Чувствительна RNA Pol III Нуклеоплазма т. РНК, 5 S р. РНК, U 6 sn. РНК и ряд других малых РНК Относительно чувствительна
 Субъединичный состав эукариотических РНК полимераз
Субъединичный состав эукариотических РНК полимераз
 полипептидные цепи, обязательно присутствующие в составе РНК полимеразы II, были выявлены посредством соосаждения с одной из субъединиц, содержащей дополнительный (искусственно добавленный к гену) эпитоп субъединицы, которые фосфорилируются
полипептидные цепи, обязательно присутствующие в составе РНК полимеразы II, были выявлены посредством соосаждения с одной из субъединиц, содержащей дополнительный (искусственно добавленный к гену) эпитоп субъединицы, которые фосфорилируются
 Сходство больших субъединиц дрожжевой РНК полимераза II с большими субъединицами РНК полимеразы II Е. Coli Домены с высокой степенью идентичности (Yeast/E/Coli)показаны красным Цифрами показаны позиции, мутации по которым приводят к нарушению функций фермента
Сходство больших субъединиц дрожжевой РНК полимераза II с большими субъединицами РНК полимеразы II Е. Coli Домены с высокой степенью идентичности (Yeast/E/Coli)показаны красным Цифрами показаны позиции, мутации по которым приводят к нарушению функций фермента
 Наиболее хорошо изучена РНК полимераза дрожжей. Клонированы гены, кодирующие 12 субъединиц этой РНК полимеразы. Получены кристаллы (без слабо удерживаемых в составе комплекса субъединиц Rpb 4 и Rpb 7. Проведен рентгеноструктурный анализ/ Сужение канала
Наиболее хорошо изучена РНК полимераза дрожжей. Клонированы гены, кодирующие 12 субъединиц этой РНК полимеразы. Получены кристаллы (без слабо удерживаемых в составе комплекса субъединиц Rpb 4 и Rpb 7. Проведен рентгеноструктурный анализ/ Сужение канала
 3 А разрешение
3 А разрешение
 m. RNA start -110 -40 -25 Py. ANT/APy. Py CCAAT box CTF GC box TATA box Sp 1 downstream element inr TBP Эукариотическая РНК полимераза сама по себе не узнает ДНК. Для ее посадки на промотор требуются общие факторы транскрипции TBP высоко консервативен у эукариот связывается с малой бороздкой о вызывает изгиб ДНК (45 ) необходим для транскрипции даже с тех промоторов, которые не содержат ТАТА бокса
m. RNA start -110 -40 -25 Py. ANT/APy. Py CCAAT box CTF GC box TATA box Sp 1 downstream element inr TBP Эукариотическая РНК полимераза сама по себе не узнает ДНК. Для ее посадки на промотор требуются общие факторы транскрипции TBP высоко консервативен у эукариот связывается с малой бороздкой о вызывает изгиб ДНК (45 ) необходим для транскрипции даже с тех промоторов, которые не содержат ТАТА бокса
 ТВР является частью общего транскрипционного фактора TFIID = TBP + TAFIIS наиболее консервативны 8 TAFIIS
ТВР является частью общего транскрипционного фактора TFIID = TBP + TAFIIS наиболее консервативны 8 TAFIIS
 TAFII 250+TAFII 150 - inr TAFII 110 - SP 1
TAFII 250+TAFII 150 - inr TAFII 110 - SP 1
 факторов транскрипции") для инициации транскрипции РНК полимеразой II необходимы шесть так называемых «общих» (general) факторов транскрипции сайты связывания транскрипционных факторов (не всегда) UAS core promoter Upstream Activating Sequences TATA inr участок начала транскрипции YYANt/a. YY пиримидиновый бокс ген
для инициации транскрипции РНК полимеразой II необходимы шесть так называемых «общих» (general) факторов транскрипции сайты связывания транскрипционных факторов (не всегда) UAS core promoter Upstream Activating Sequences TATA inr участок начала транскрипции YYANt/a. YY пиримидиновый бокс ген
 UAS TFIID Upstream Activating") TAFs сайты связывания транскрипционных факторов core promoter TBP (не всегда) UAS TFIID Upstream Activating Sequences TATA inr участок начала транскрипции YYANt/a. YY пиримидиновый бокс ген
TAFs сайты связывания транскрипционных факторов core promoter TBP (не всегда) UAS TFIID Upstream Activating Sequences TATA inr участок начала транскрипции YYANt/a. YY пиримидиновый бокс ген
 различные транскрипционные факторы TAFs TFIID UAS TBP TATA core promoter inr участок начала транскрипции YYANt/a. YY пиримидиновый бокс ген
различные транскрипционные факторы TAFs TFIID UAS TBP TATA core promoter inr участок начала транскрипции YYANt/a. YY пиримидиновый бокс ген
 TAFs TFIID UAS TBP TFIIA TFIIB inr ген TFIIA стабилизируют взаимодействие TBP с ДНК, контактируя одновременно с ДНК и TBP
TAFs TFIID UAS TBP TFIIA TFIIB inr ген TFIIA стабилизируют взаимодействие TBP с ДНК, контактируя одновременно с ДНК и TBP
 TAFs TFIID UAS TBP TFIIA Pol II TFIIF TFIIB inr TFIIF обеспечивает посадку РНК полимеразы II на формирующийся коплекс TFIIB промежуточным звеном при связывании РНК полимеразы II и TBP ген
TAFs TFIID UAS TBP TFIIA Pol II TFIIF TFIIB inr TFIIF обеспечивает посадку РНК полимеразы II на формирующийся коплекс TFIIB промежуточным звеном при связывании РНК полимеразы II и TBP ген
 mediator TAFs TFIID UAS TBP Pol II TFIIF TFIIA TFIIB TFIIH ген TFIIE Последними к комплексу присоединяются TFIIE и TFIIH Включает киназу и АТФ-зависимую ДНК-хеликазу, которая обеспечивает локальное плавление ДНК в точке начала транскрипции
mediator TAFs TFIID UAS TBP Pol II TFIIF TFIIA TFIIB TFIIH ген TFIIE Последними к комплексу присоединяются TFIIE и TFIIH Включает киназу и АТФ-зависимую ДНК-хеликазу, которая обеспечивает локальное плавление ДНК в точке начала транскрипции
 Фрагменты ДНК, взаимодействующие с белками, можно идентифицировать с использованием метода band-shift. В дальнейшем сайты связывания белков можно картировать с использованием различных вариантов фут-принтинга band-shift экстракт специфический конкурент footprinting
Фрагменты ДНК, взаимодействующие с белками, можно идентифицировать с использованием метода band-shift. В дальнейшем сайты связывания белков можно картировать с использованием различных вариантов фут-принтинга band-shift экстракт специфический конкурент footprinting
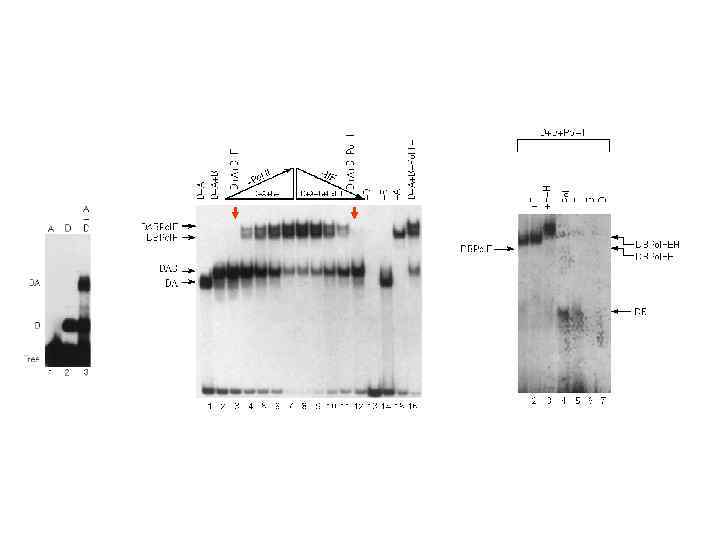
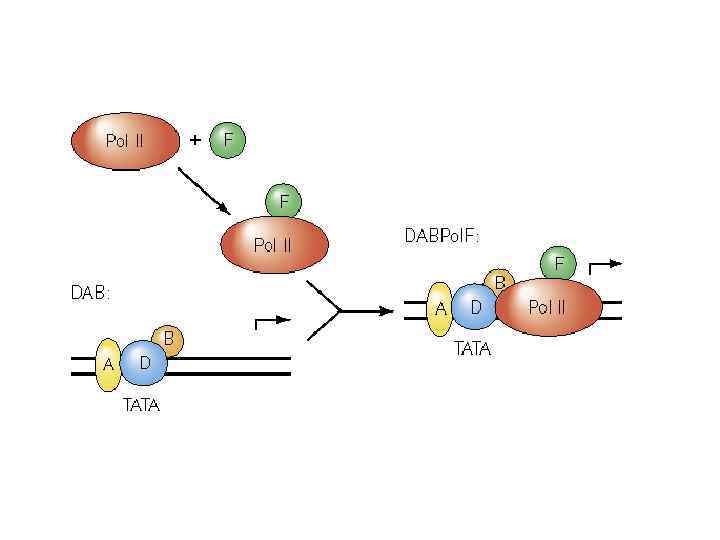
 комплекс DAB без TF 2 F и Pol II DAB +pol. F
комплекс DAB без TF 2 F и Pol II DAB +pol. F
 Перед началом элонгации выделяют особую стадию «освобождения промотора» При этом часть общих транскрипционных факторов остается на промоторе, тогда как другие (TFIIF) перемещаются вместе с элонгирующим полимеразным комплексом. В момент освобождения промотора элонгирующий комплекс остается крайне нестабильным и может легко «соскачить с ДНК» . Комплекс заметно стабилизируется после включения 8 -9 нуклеотидных остатков (именно такова длина ДНК-РНК гибрида в составе транскрипционного комплекса) Важную роль в переходе от «инициаторного» к элонгирующему комплексу играет фософорилирование сериновых остатков 2 и 5 в повторяющихся много раз гегтапептидах YSPTSPS С’-концевого домена каталитической субъединицы РНК-полимеразы II. Первичное фосфорилирование серинов 5 осуществляет киназная субъединица (Kin 28) транскрипционного фактора TFIIH
Перед началом элонгации выделяют особую стадию «освобождения промотора» При этом часть общих транскрипционных факторов остается на промоторе, тогда как другие (TFIIF) перемещаются вместе с элонгирующим полимеразным комплексом. В момент освобождения промотора элонгирующий комплекс остается крайне нестабильным и может легко «соскачить с ДНК» . Комплекс заметно стабилизируется после включения 8 -9 нуклеотидных остатков (именно такова длина ДНК-РНК гибрида в составе транскрипционного комплекса) Важную роль в переходе от «инициаторного» к элонгирующему комплексу играет фософорилирование сериновых остатков 2 и 5 в повторяющихся много раз гегтапептидах YSPTSPS С’-концевого домена каталитической субъединицы РНК-полимеразы II. Первичное фосфорилирование серинов 5 осуществляет киназная субъединица (Kin 28) транскрипционного фактора TFIIH
 Для РНК полимеразы II и ее аналогов у других организмов характерна тенденция к временной (пауза) или бессрочной (арест) остановке элонгации. Для обеспечения эффективной элонгации необходим целый ряд вспомогательных факторов, один из которых (TFIIF) является общим фактором транскрипции. Существует и негативный фактор элонгации NELF, который стимулирует остановку Pol. II. NELF связывается с комплексом Pol. II-DSIF стимулирует РНказную активность полимеразы II
Для РНК полимеразы II и ее аналогов у других организмов характерна тенденция к временной (пауза) или бессрочной (арест) остановке элонгации. Для обеспечения эффективной элонгации необходим целый ряд вспомогательных факторов, один из которых (TFIIF) является общим фактором транскрипции. Существует и негативный фактор элонгации NELF, который стимулирует остановку Pol. II. NELF связывается с комплексом Pol. II-DSIF стимулирует РНказную активность полимеразы II
 Для РНК полимеразы II и ее аналогов у других организмов характерна тенденция к временной (пауза) или бессрочной (арест) остановке элонгации. Для обеспечения эффективной элонгации необходим целый ряд вспомогательных факторов, один из которых (TFIIF) является общим фактором транскрипции. Существует и негативный фактор элонгации NELF, который стимулирует остановку Pol. II. NELF связывается с комплексом Pol. II-DSIF стимулирует РНказную активность полимеразы II
Для РНК полимеразы II и ее аналогов у других организмов характерна тенденция к временной (пауза) или бессрочной (арест) остановке элонгации. Для обеспечения эффективной элонгации необходим целый ряд вспомогательных факторов, один из которых (TFIIF) является общим фактором транскрипции. Существует и негативный фактор элонгации NELF, который стимулирует остановку Pol. II. NELF связывается с комплексом Pol. II-DSIF стимулирует РНказную активность полимеразы II
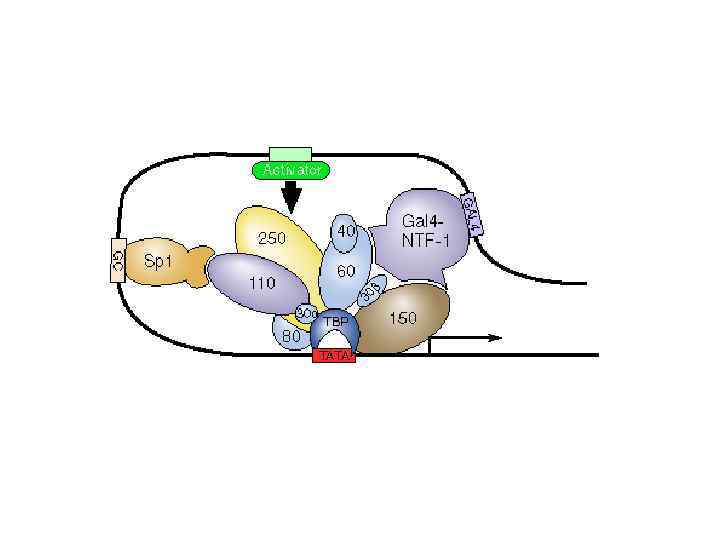
 энхансер минимальный") Энхансеры и сайленсеры модулируют «скорость» транскрипции 1 минимальный промотор 2 ген-репортер (САТ) энхансер минимальный промотор ген-репортер (САТ) 3 энхансер минимальный промотор ген-репортер (САТ) количество продукта или м. РНК 1 2 3 номер конструкта
Энхансеры и сайленсеры модулируют «скорость» транскрипции 1 минимальный промотор 2 ген-репортер (САТ) энхансер минимальный промотор ген-репортер (САТ) 3 энхансер минимальный промотор ген-репортер (САТ) количество продукта или м. РНК 1 2 3 номер конструкта
 Энхансеры могут располагаться где угодно перед промотором иногда непосредственно и тогда они фактически выполняют роль UAS иногда на большом расстоянии (более 100 Kb) р р ген внутри гена р ген после гена р ген
Энхансеры могут располагаться где угодно перед промотором иногда непосредственно и тогда они фактически выполняют роль UAS иногда на большом расстоянии (более 100 Kb) р р ген внутри гена р ген после гена р ген
 Не вполне ясно, что именно регулируют энхансеры А. Скорость работы промотора (число инициаций в единицу времени) ? Б. Статус промотора (активный / неактивный) ? Механизм работы энхансеров также остается неясным
Не вполне ясно, что именно регулируют энхансеры А. Скорость работы промотора (число инициаций в единицу времени) ? Б. Статус промотора (активный / неактивный) ? Механизм работы энхансеров также остается неясным
 Все известные в настоящее время энхансеры представляют собой площадки для связывания транскрипционных факторов. Транскрипционные факторы могут быть универсальными и тканеспецифичными Соответсвенно, и энхансеры могут быть универсальными и тканеспецифичными Большинстви вирусных энхансеров являются универсальными Энхансеры вирусов SV-40, Polyoma, CMV универсальный транскрипционный фактор Sp 1 Примером тканеспецифичных энхансеров могут служить энхансеры глобиновых генов эритроид-специфичный транскрипционный фактор GATA-1
Все известные в настоящее время энхансеры представляют собой площадки для связывания транскрипционных факторов. Транскрипционные факторы могут быть универсальными и тканеспецифичными Соответсвенно, и энхансеры могут быть универсальными и тканеспецифичными Большинстви вирусных энхансеров являются универсальными Энхансеры вирусов SV-40, Polyoma, CMV универсальный транскрипционный фактор Sp 1 Примером тканеспецифичных энхансеров могут служить энхансеры глобиновых генов эритроид-специфичный транскрипционный фактор GATA-1
 Транскрипционные факторы обычно содержат ДНК связывающий и активаторный домены Во многих случаях эти домены являются функционально независимыми Активаторный домен дрожжевого фактора GAL 4 можно присоединить к ДНКсвязывающему домену прокариотического репрессора Lex. A. При наличии в дрожжевых клетках такого химерного фактора сайты связывания Lex. A будут стимулировать транскрипцию
Транскрипционные факторы обычно содержат ДНК связывающий и активаторный домены Во многих случаях эти домены являются функционально независимыми Активаторный домен дрожжевого фактора GAL 4 можно присоединить к ДНКсвязывающему домену прокариотического репрессора Lex. A. При наличии в дрожжевых клетках такого химерного фактора сайты связывания Lex. A будут стимулировать транскрипцию
 Activators Have Two Domains: DNA Binding and Activation Domains DNA Binding Activation • Zn-containing modules • Acidic domains • Homeodomains • Glutamine-rich domains • b. ZIP and b. HLH domains • Proline-rich domains
Activators Have Two Domains: DNA Binding and Activation Domains DNA Binding Activation • Zn-containing modules • Acidic domains • Homeodomains • Glutamine-rich domains • b. ZIP and b. HLH domains • Proline-rich domains
 DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • b. ZIP and b. HLH domains • First identified in transcription factor TFIIIA • NMR used to solve first structure (1989) • Some transcription factors have multiple Zn fingers. Zif 268 from mouse has three. Residues from each -helix make sequence-specific contacts in the major groove (1991). • Zn fingers bind with sequence specificity, but no simple code exists to predict the DNA sequence that a given zinc finger motif will recognize.
DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • b. ZIP and b. HLH domains • First identified in transcription factor TFIIIA • NMR used to solve first structure (1989) • Some transcription factors have multiple Zn fingers. Zif 268 from mouse has three. Residues from each -helix make sequence-specific contacts in the major groove (1991). • Zn fingers bind with sequence specificity, but no simple code exists to predict the DNA sequence that a given zinc finger motif will recognize.
 DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • b. ZIP and b. HLH domains GAL 4 DNA binding domain • Controls a set of genes responsible for galactose metabolism • Binds to DNA as a dimer formed by coiled-coil interaction • Each monomer binds two Zn ions
DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • b. ZIP and b. HLH domains GAL 4 DNA binding domain • Controls a set of genes responsible for galactose metabolism • Binds to DNA as a dimer formed by coiled-coil interaction • Each monomer binds two Zn ions
 DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • Members of helix-turn-helix family • Discovered in regulatory genes of Drosophila • Helix 3 occupies major groove and an extension of helix 1 occupies minor groove • b. ZIP and b. HLH domains T. Kornberg, C. Pabo and colleagues, 1990
DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • Members of helix-turn-helix family • Discovered in regulatory genes of Drosophila • Helix 3 occupies major groove and an extension of helix 1 occupies minor groove • b. ZIP and b. HLH domains T. Kornberg, C. Pabo and colleagues, 1990
 DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • b. ZIP and b. HLH domains • b. ZIP dimer forms a ‘leucine zipper’ coiled-coil. Each helix contacts DNA in the major groove. • ‘b’ for basic region, which contacts DNA ‘ZIP’ for the dimerization (zipper) region consisting of amphipathic helices that form parallel coiled-coil interactions Structures of GCN 4 Yeast protein that up-regulates amino acid biosynthesis
DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • b. ZIP and b. HLH domains • b. ZIP dimer forms a ‘leucine zipper’ coiled-coil. Each helix contacts DNA in the major groove. • ‘b’ for basic region, which contacts DNA ‘ZIP’ for the dimerization (zipper) region consisting of amphipathic helices that form parallel coiled-coil interactions Structures of GCN 4 Yeast protein that up-regulates amino acid biosynthesis
 DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • b. ZIP and b. HLH domains Myo. D Involved in muscle development
DNA Binding Domains DNA Binding • Zn-containing modules • Homeodomains • b. ZIP and b. HLH domains Myo. D Involved in muscle development
.") Pol I промотор состоит из двух важных элементов: CORE и UCA (Upstream Control Element). Гомология между существенными элементами Pol. I промотора разных организмов практически отсутствует. Pol. I промотор человека
Pol I промотор состоит из двух важных элементов: CORE и UCA (Upstream Control Element). Гомология между существенными элементами Pol. I промотора разных организмов практически отсутствует. Pol. I промотор человека
 Расстояние между двумя элементами Pol. I промотора существенно для его работы
Расстояние между двумя элементами Pol. I промотора существенно для его работы
 -165 -75 A B UCE UBF Может работать c Pol I разных видов core SL 1 = TBP (тот же, который входит в состав TFIID) + Pol I TAFs TAF I 110 TAF I 63 TAF I 48
-165 -75 A B UCE UBF Может работать c Pol I разных видов core SL 1 = TBP (тот же, который входит в состав TFIID) + Pol I TAFs TAF I 110 TAF I 63 TAF I 48
 Pol III промоторы Ряд Pol III генов содержат промоторы внутри генов Другие Pol III гены (U 6) имеют промоторы сходные с промоторами Pol II генов 5 S р. РНК промотор связываются факторы TFIIIA, TFIIIB, TFIIIC t. РНК промотор и связываются факторы TFIIIB и TFIIIC неканонические Pol III промоторы TFIIIB включает TBP и TAFIIIs
Pol III промоторы Ряд Pol III генов содержат промоторы внутри генов Другие Pol III гены (U 6) имеют промоторы сходные с промоторами Pol II генов 5 S р. РНК промотор связываются факторы TFIIIA, TFIIIB, TFIIIC t. РНК промотор и связываются факторы TFIIIB и TFIIIC неканонические Pol III промоторы TFIIIB включает TBP и TAFIIIs
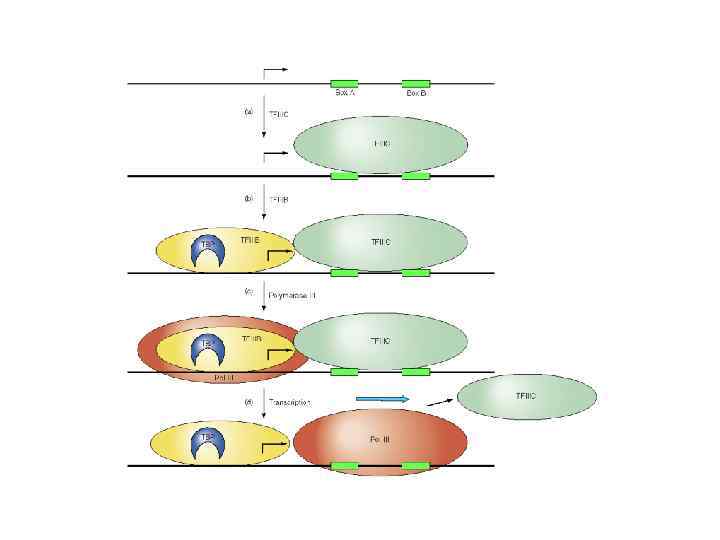
 нужен для инициации транскрипции всеми тремя классами эукариотических") TBP (в ряде случаев его аналоги) нужен для инициации транскрипции всеми тремя классами эукариотических РНК полимераз
TBP (в ряде случаев его аналоги) нужен для инициации транскрипции всеми тремя классами эукариотических РНК полимераз
 УРОВНИ УПАКОВКИ ДНК В ЯДРЕ ЭУКАРИOТИЧЕСКОЙ КЛЕТКИ Размер человеческой ДНК - 1, 8 м Размер ядра - 10 мкм Степень упаковки - 40 000 х
УРОВНИ УПАКОВКИ ДНК В ЯДРЕ ЭУКАРИOТИЧЕСКОЙ КЛЕТКИ Размер человеческой ДНК - 1, 8 м Размер ядра - 10 мкм Степень упаковки - 40 000 х
 ДНК НУКЛЕОСОМЫ ДОМЕНЫ ХРОМОСОМЫ
ДНК НУКЛЕОСОМЫ ДОМЕНЫ ХРОМОСОМЫ
 Компактизация ДНК в ядре осуществляется при посредстве сложного комплекса белков, среди которых принято выделять гистоны и негистоновые белки Гистоны являются чрезвычайно консервативными белками. Выделяют 5 основных типов гистонов Н 1, Н 2 А, Н 2 В, Н 3, Н 4 Гистоны Н 2 А, Н 2 В, Н 3, Н 4 входят в состав минимальной нуклеосомы (так называемой “Core Particle”). По-этому их нередко называют «коровыми» гистонами
Компактизация ДНК в ядре осуществляется при посредстве сложного комплекса белков, среди которых принято выделять гистоны и негистоновые белки Гистоны являются чрезвычайно консервативными белками. Выделяют 5 основных типов гистонов Н 1, Н 2 А, Н 2 В, Н 3, Н 4 Гистоны Н 2 А, Н 2 В, Н 3, Н 4 входят в состав минимальной нуклеосомы (так называемой “Core Particle”). По-этому их нередко называют «коровыми» гистонами
 линкерный гистоны нуклеосомного ядра
линкерный гистоны нуклеосомного ядра
 К открытию нуклеосом привели две группы экспериментов: 1. Электронная микроскопия препаратов хроматина в низкой ионной силе 2. Анализ продуктов расщепления хроматина стафилококковой нуклеазой
К открытию нуклеосом привели две группы экспериментов: 1. Электронная микроскопия препаратов хроматина в низкой ионной силе 2. Анализ продуктов расщепления хроматина стафилококковой нуклеазой
 Нуклеосома является базовой структурной единицей первого уровня упаковки ДНК в хроматине. Она представляет собой белковую глобулу, или, точнее говоря некое подобие диска, на который намотан фрагмент ДНК протяженностью 146 п. н. (1, 65 витка) Глобула состоит из восьми молекул гистонов: тетрамера (Н 3)2 -(Н 4)2 и двух димеров Н 2 А-Н 2 В. Диаметр глобулыдиска составляет ~11 нм, а высота - ~ 5, 7 нм H 1 связывается с ДНК, входящей на нуклеосому, и ДНК, выходящей из нуклеосомы, стабилизируя глобулу и обеспечивая замыкание двух полных витков ДНК
Нуклеосома является базовой структурной единицей первого уровня упаковки ДНК в хроматине. Она представляет собой белковую глобулу, или, точнее говоря некое подобие диска, на который намотан фрагмент ДНК протяженностью 146 п. н. (1, 65 витка) Глобула состоит из восьми молекул гистонов: тетрамера (Н 3)2 -(Н 4)2 и двух димеров Н 2 А-Н 2 В. Диаметр глобулыдиска составляет ~11 нм, а высота - ~ 5, 7 нм H 1 связывается с ДНК, входящей на нуклеосому, и ДНК, выходящей из нуклеосомы, стабилизируя глобулу и обеспечивая замыкание двух полных витков ДНК
 Модульный принцип посторения нуклеосомнорго ядра отражает характер его сборки
Модульный принцип посторения нуклеосомнорго ядра отражает характер его сборки
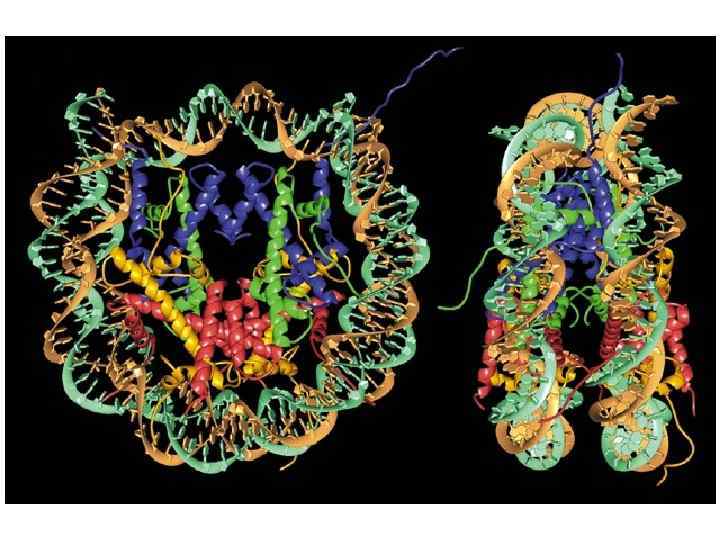
 Гистоны контактируют с фосфодиэфирным остовом молекулы ДНК. Контакты реализуются через каждые 10 пар оснований, когда малая бороздка ДНК оказывается развернутой внутрь. В целом, накрученная на нуклеомную глобулу ДНК образует 14 прочных электростатических контактов с молекулами гистонов.
Гистоны контактируют с фосфодиэфирным остовом молекулы ДНК. Контакты реализуются через каждые 10 пар оснований, когда малая бороздка ДНК оказывается развернутой внутрь. В целом, накрученная на нуклеомную глобулу ДНК образует 14 прочных электростатических контактов с молекулами гистонов.
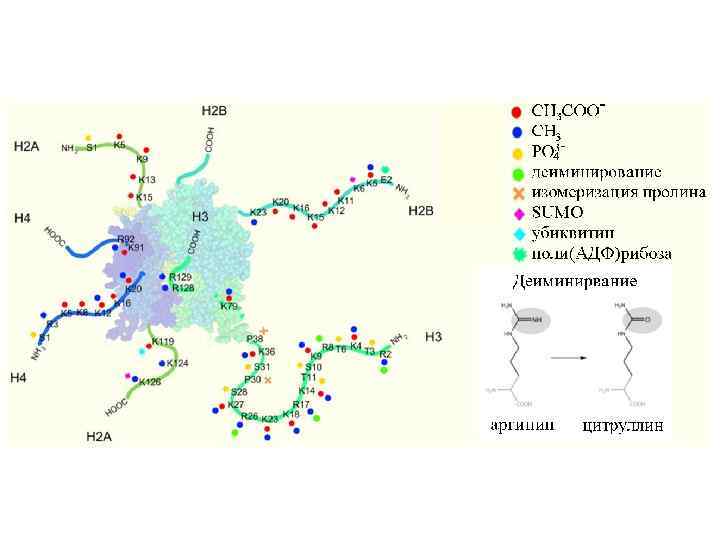
 N-концевые фрагменты гистонов выходят за пределы нуклеосомного ядра. На них находится много мишеней для модификаций, имеющих сигнальное значение
N-концевые фрагменты гистонов выходят за пределы нуклеосомного ядра. На них находится много мишеней для модификаций, имеющих сигнальное значение
 Бороздки в двух витках ДНК ориентированы параллельно. В этой связи остается Достаточно места для выхода «за пределы глобулы» хвостов гистонов Н 3 и Н 2 В
Бороздки в двух витках ДНК ориентированы параллельно. В этой связи остается Достаточно места для выхода «за пределы глобулы» хвостов гистонов Н 3 и Н 2 В