Физиология,практика3, мышцы.ppt
- Количество слайдов: 58


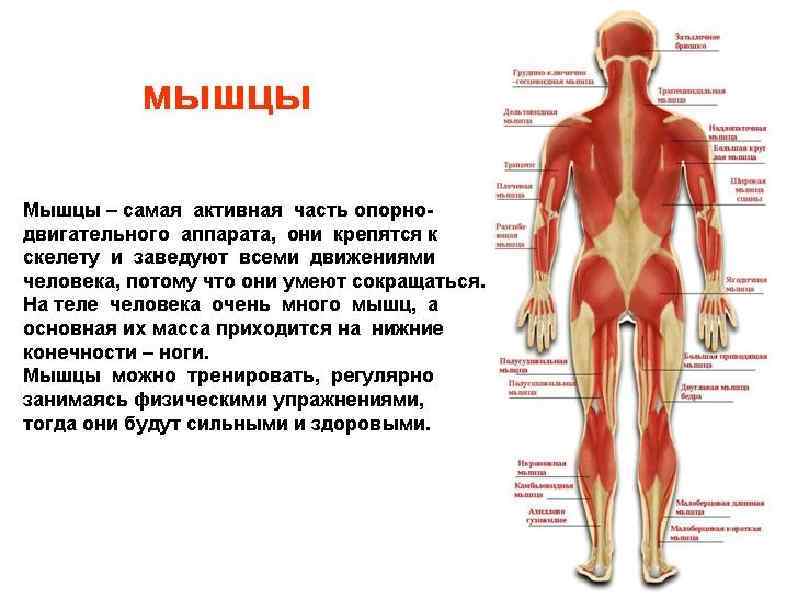
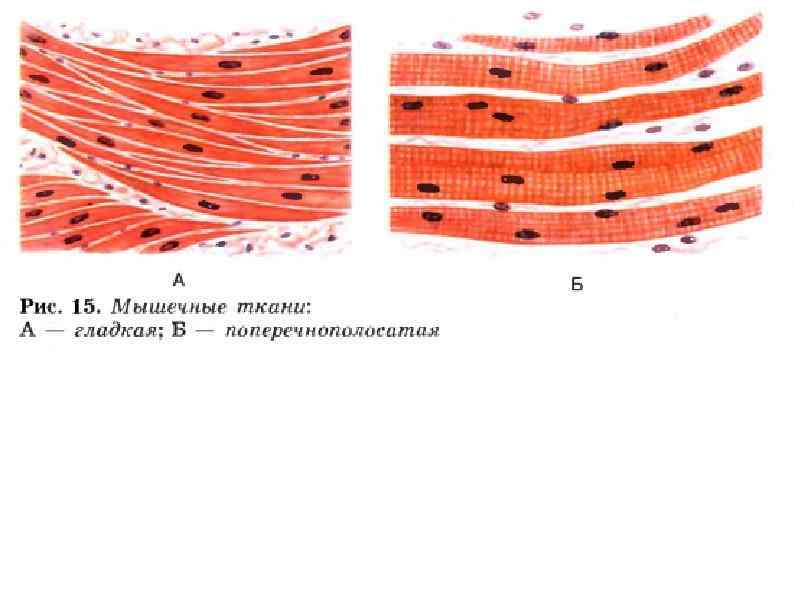
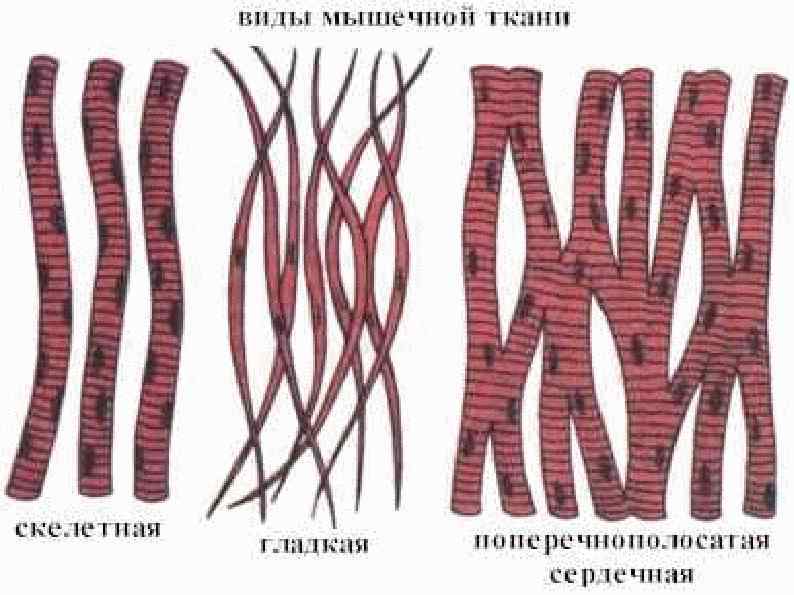

Мышцы Обеспечивают перемещение тела и его частей в пространстве Поддерживают тонус сосудов и полостных органов Генерируют тепло при мышечной работе и в расслабленном состоянии Обеспечивают работу сердца

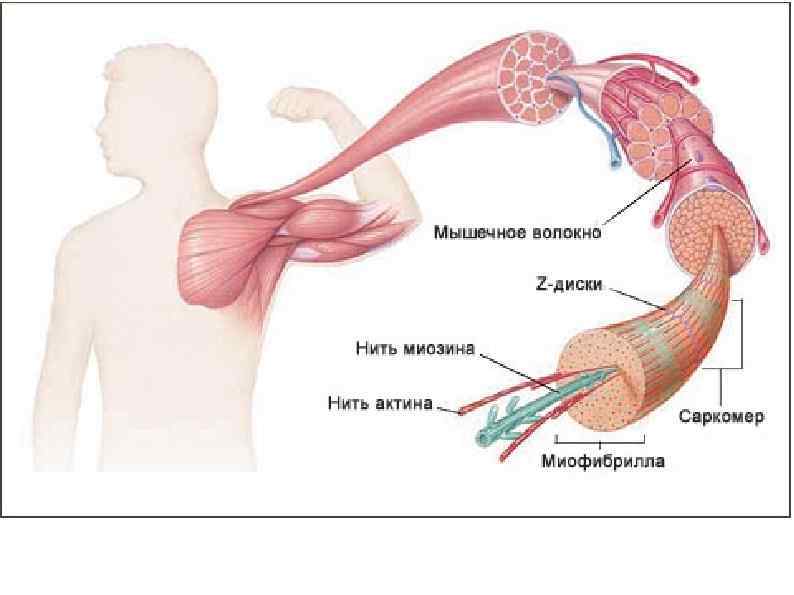
 структурная единица скелетных мышц")
МЫШЕЧНОЕ ВОЛОКНО (МИОЦИТ) структурная единица скелетных мышц

ХИМИЧЕСКИЙ СОТАВ МИОЦИТА БЕЛКИ ферменты гликолиза, ферменты креатинкиназа, белок миоглобин (связывает молекулярный кислород и запасает его), АТФ, АДФ, АМФ, креатинфосфат, креатин, молочная кислота. УГЛЕВОДЫ. Гликоген ЖИР связан с белками (концентрация 1%)
 А – зона (анизотропная темная зона) – толстая нить")
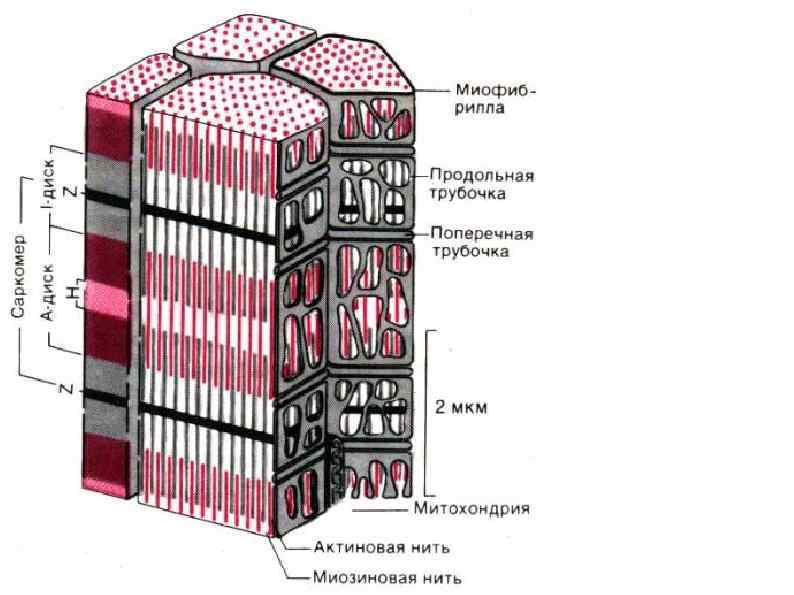
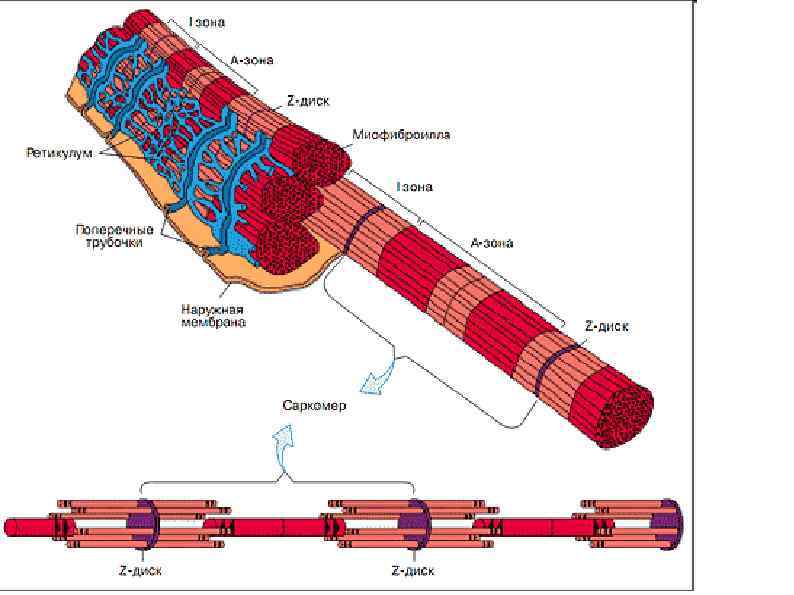
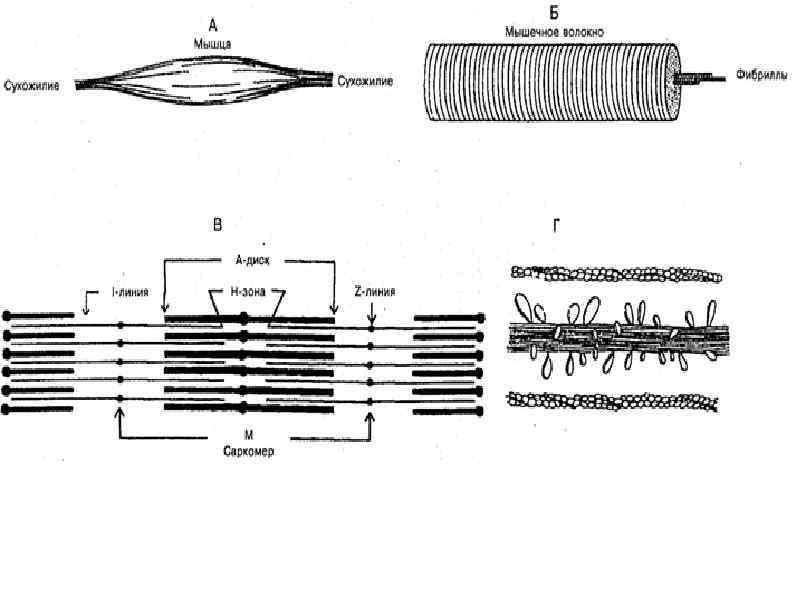
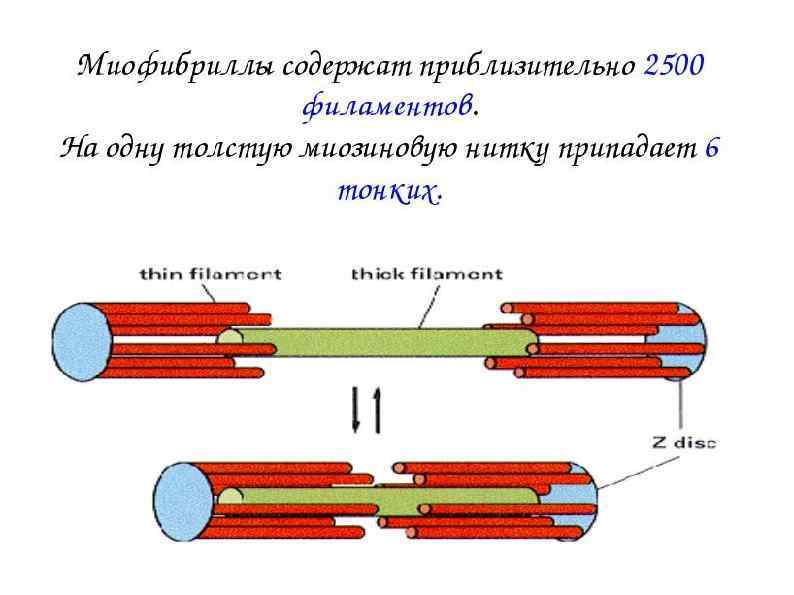
СТРОЕНИЕ МИОФИБРИЛЛЫ (сократительная нить миоцита) А – зона (анизотропная темная зона) – толстая нить (миозин) I – зона (изотропные светлые участки) – тонкая нить (актин, тропонин, тропомиозин) Н – светлая полоска, в пределах анизотропного диска Z – промежуточный диск, темная полоска в пределах изотропного диска. Комплекс – А, I, H – зоны между двумя Z – дисками – саркомер.
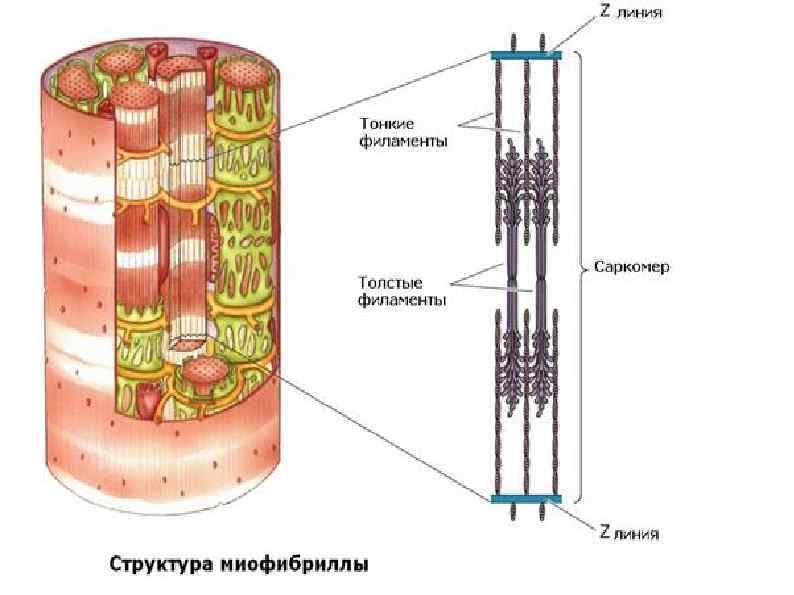

Саркомер Сократительная единица мышечного волокна. Представляет собой повторяющийся сегмент миофибрилы, состоящая из системы толстых и тонких нитей
 ― структурный белок актиновой нити, (вытянутый в виде тяжа")
СТРОЕНИЕ АКТИНОВОГО ФИЛАМЕНТА Тропомиозин (Тм) ― структурный белок актиновой нити, (вытянутый в виде тяжа молекулы). Тропонин (Тн) - регуляторный белок актиновой нити. Состоит из трех субъединиц: Тропонин Т(Тн. Т) — обеспечивает связывание этих белков с тропомиозином. Тропонин I (Тн. I) — блокирует (ингибирует) взаимодействие актина с миозином. Тропонин С (Тн. С) ― это Са 2+-связывающий белок. В присутствии Са 2+ изменяется конформация тропонина С в результате чего открывается центр взаимодействия актина с миозином.

СТРОЕНИЕ МИОЗИНОВОГО ФИЛАМЕНТА

БЕЛКИ МЫШЕЧНОЙ ТКАНИ Саркоплазматические белки – белки-ферменты локализованные главным образом в митохондриях и катализирующие процессы окислительного фосфорилирования. А так же билок миоглобин (связь с кислородом) Миофибриллярные белки – обеспечивают сократительную функцию мышц
 Имеет")
Сократительные: миозин, актин и актомиозин Актин ― сократительный белок мышц (основа тонких нитей) Имеет две формы ― глобулярный G-актин и фибриллярный F-актин. Каждая молекула G-актина способна связывать один ион Са 2+ и одну молекулу АТФ или АДФ. Образование F-актина сопровождается полимеризацией G-актина при связывании его с АТФ и одновременным расщеплением АТФ до АДФ и фосфата. Нить F-актина может связывать большое число молекул миозина. Актин способен взаимодействовать с миозином, образуя актомиозиновый комплекс (его диссоциация происходит в присутствии АТФ и Мg 2+).

Миофибриллярные белки Сократительные: миозин, актин и актомиозин Миозин - основной сократительный белков мышц (толстые нити). Состоит из длинной фибриллярной части и глобулярной структуры (головки). Фибриллярная часть - двуспиральная структура. Функция - образовывать упорядоченные пучки миозиновых филаментов. Головка молекулы миозина - активный центр АТФ-азы и актинсвязывающий центр Обеспечивают расщепление АТФ и взаимодействие с актиновыми филаментами.

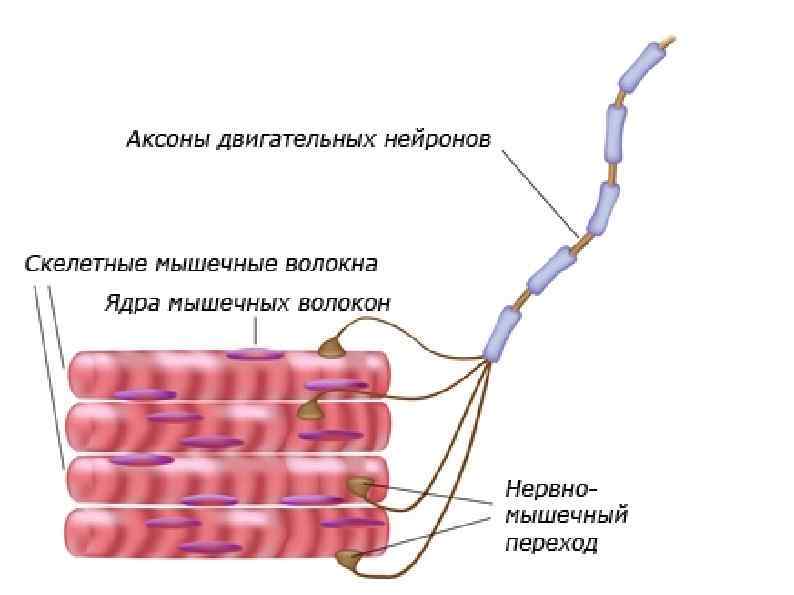
НЕРВНО-МЫШЕЧНЫЙ СИНАПС Место контакта нервного окончания с сарколемой
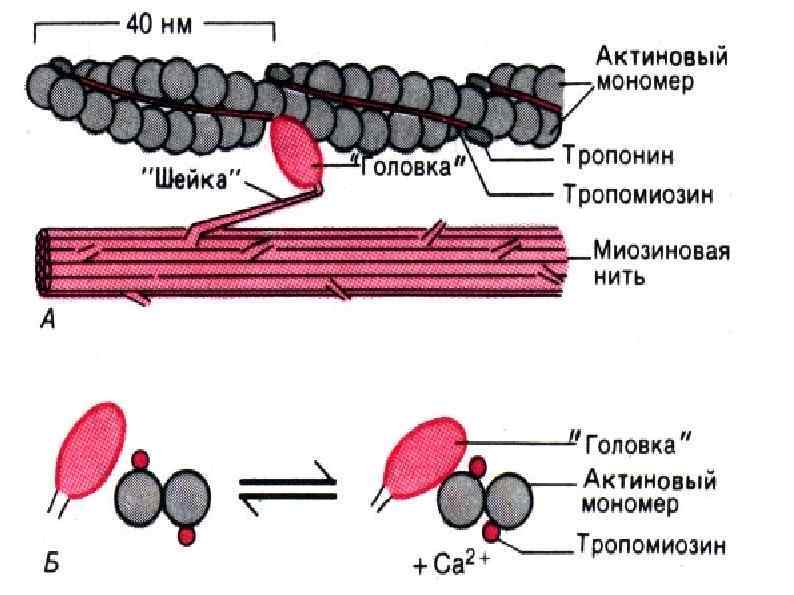
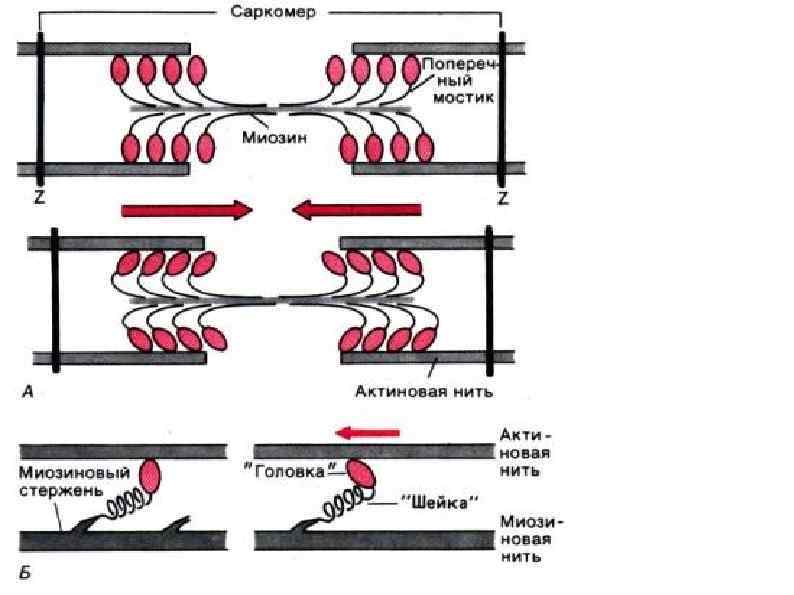
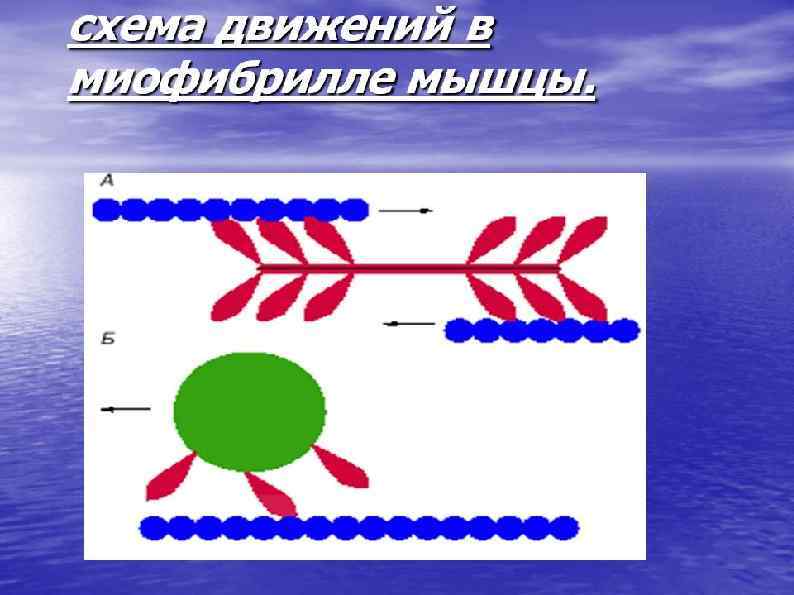

Механизм мышечного сокращения Сокращение запускается нервным импульсом Нервный импульс вызывает выход ионов кальция из саркоплазматического ретикулума. Ионы кальция присоединяются к тропонину, меняя его форму, открывается активный участок актина. Между тонкими и толстыми нитями миофибрилл возникают поперечные мостики, или спайки Повышается активность миозина, он стимулирует расщепление АТФ. С использованием энергии АТФ прооисходит скольжение тонких нитей вдоль толстых, что приводит к укорочению миофибрилл и всего мышечного волокна в целом.

МЕХАНИЗМ МЫШЕЧНОГО РАССЛАБЛЕНИЯ • Прекращается нервный импульс • Ионы кальция обратно переносятся в саркоплазматический ртикулум. • Изменяется форма тропонина он освобождается от связи с кальцием. • Тропомиозин препятствует образованию связей актина и миозина. • С использованием энергии АТФ миофибрилла вворачивается в исходное состояние
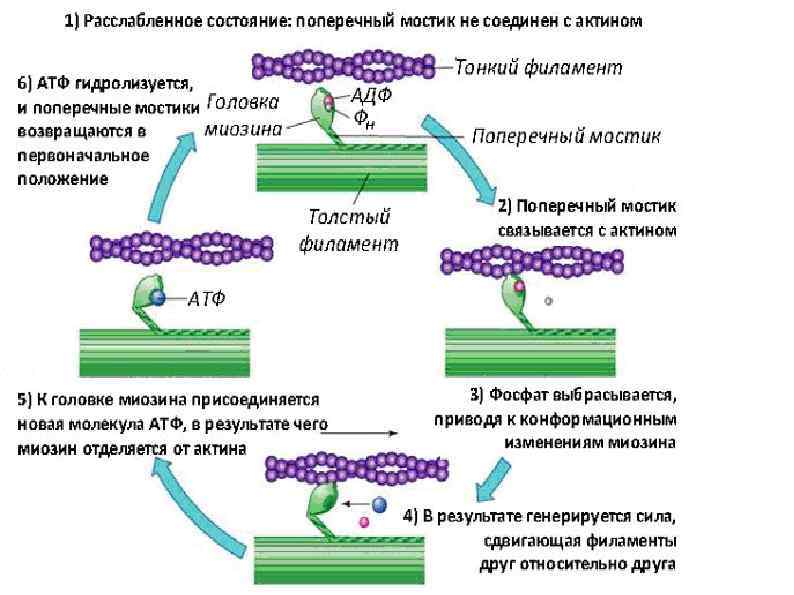

Энергетика мышечного сокращения обе фазы сокращение и расслабление мышечной деятельности - - протекают при обязательном использовании энергии, . При функционировании мышц в них одновременно протекают два процесса: гидролиз АТФ, дающий необходимую энергию для сокращения и расслабления, и ресинтез АТФ, восполняющий потери этого вещества. ПУТИ РЕСИНТЕЗА АТФ: анаэробная (бескислородная) аэробная (кислородная).

При любой мышечной работе функционируют ТРИ ПУТИ РЕСИНТЕЗА АТФ • КРЕАТИНФОСФАТНАЯ РЕАКЦИЯ – в первые секунды работы • затем подключается ГЛИКОЛИЗ • ТКАНЕВОЕ ДЫХАНИЕ - приходит на смену гликолизу по мере продолжения работы
 источник энергии АТФ,")
ЗОНЫ ОТНОСИТЕЛЬНОЙ МОЩНОСТИ МЫШЕЧНОЙ РАБОТЫ • Максимальная мощность (15 -20 сек) источник энергии АТФ, креатинфосфат. • Субмаксимальная мощность (до 5 мин) механизм ресинтеза АТФ – гликолитический • Большая мощность (до 30 мин) механизм ресинтеза АТФ – гликолиз и тканевое дыхание • Умеренная мощность (свыше 30 мин) аэробный путь ресинтеза АТФ (тканевое дыхание)

МОЛЕКУЛА АТФ В МЫШЕЧНОМ ВОЛОКНЕ (ее хватает примерно на 1 -2 сокращения, т. е. 1, 5 -2 сек) КРЕАТИН ФОСФАТ (КФ) образуется в мышцах во время отдыха при взаимодействии креатина (К) и АТФ К + АТФ = КФ Креатин образуется в печени с использованием аминокислот КФ + АДФ = АТФ + К (креатин) АДФ - образуется при распаде АТФ Креатинфосфат (КФ) – исчерпывается и в процесс ресинтеза АТФ включаются другие реакции
 =АТФ (Н действует")
ГЛИКОГЕН МОЛОЧНАЯ КИСЛОТА + 2 Н АДФ + Ф (фосфорная кислота) =АТФ (Н действует на миофибриллы, прикрепляется к актину, поэтому кальций не может прикрепиться к тропонину и мышца не сокращается) Гликолитический путь ресинтеза АТФ Источник энергии - мышечный гликоген. При анаэробном распаде гликоген превращается в молочную кислоту (лактат), эти процессы сопровождаются отщеплением фосфатных групп, которые переносятся на АДФ с образованием АТФ. Гликолиз играет важную роль в энергообеспечении упражнений, продолжительность которых составляет до 30 мин. К ним относятся бег на средние дистанции, плавание на 100 и 200 м, велосипедные гонки на треке и др

АЭРОБНОЕ ОКИСЛЕНИЕ ТКАНЕВОЕ ДЫХАНИЕ является важнейшей частью обмена веществ и энергии в организме. В результате тканевое дыхание при участии специфических ферментов происходит окислительный распад крупных органических молекул до более простых и в конечном счете до СО 2 и Н 2 О с высвобождением энергии АТФ. Осуществляется в митохондрих.

1. НИКОТИН АМИДНЫЕ ДЕГИДРОГЕНАЗЫ отнимают два 2 Н и присоединяют их к своему коферменту НАД АН 2 (окисляемое вещество) + НАД = А (окисленное вещество) + НАД*Н 2 2. ФЛАВИНОВЫЕ ДЕГИДРОГЕНАЗЫ отщепляют 2 Н от НАД*Н 2 и присоединяют их к своему коферменту ФМН НАД*Н 2 + ФМН = НАД + ФМН*Н 2 Иногда Флавиновые дегидрогеназы используют кофермент ФАД АН 2 (окисляемое вещество) + ФАД = А (окисленное вещество) + ФАД*Н 2 3. Активный кислород связывается с ионами Н, которые отщепляются от ФАД*Н 2 или ФМН*Н 2

МИОКИНАЗНАЯ РЕАКЦИЯ Происходит в мышце при значительном увеличении концентрации АДФ в саркоплазме, когда возможности других путей почти исчерпаны или близки к тому. Суть этой реакции состоит в том, что при взаимодействии 2 молекул АДФ образуется 1 молекула АТФ: АДФ + АДФ = АТФ Условия для включения миокиназной реакции возникают при выраженном мышечном утоплении. Поэтому миокиназную реакцию следует рассматривать как "аварийный" механизм

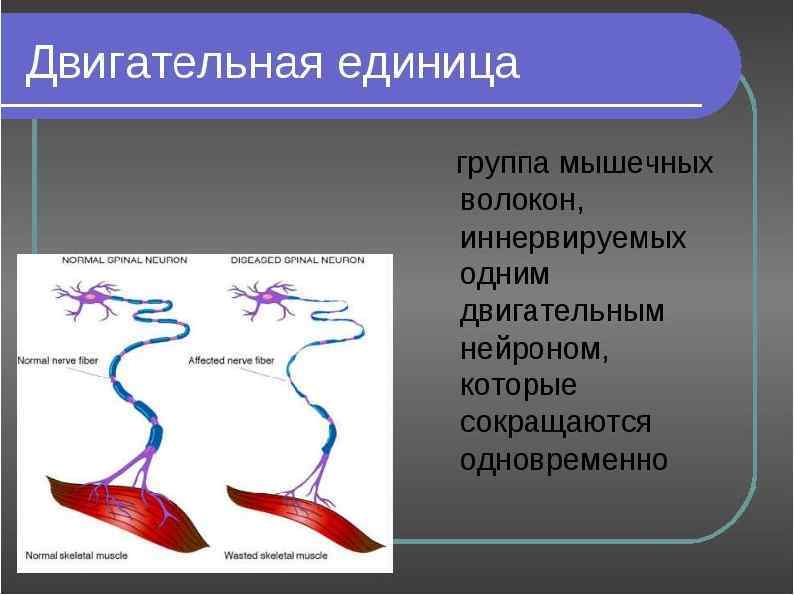
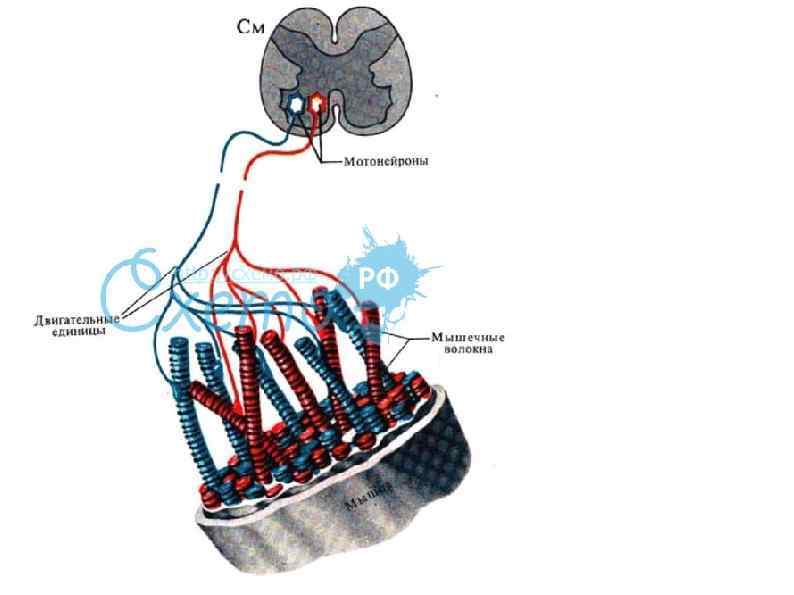
Инервация скилетных мышц К каждой мышце подходит много двигательных нейронов, которые ветвятся и иннервируют группы мышечных волокон. Каждая такая группа вместе с иннервирующим ее аксоном называется двигательной единицей. При стимуляции все мышечные волокна в ней сокращаются одновременно. Число волокон в двигательной единице варьирует и зависит от того, насколько сложным должно быть управление данной мышцей.
 ТОНИЧЕСКИЕ МЕДЛЕННЫЕ (красные)")
МЫШЕЧНЫЕ ВОЛОКНА ФАЗНЫЕ БЫСТРЫЕ (белые) ТОНИЧЕСКИЕ МЕДЛЕННЫЕ (красные)


ТОНИЧЕСКИЕ МЫШЦЫ Способностью длительное время работать без утомления Часть волокон напряжена, а остальные – расслаблены. При тоническом сокращении мышечные волокна функционируют асинхронно: участки напряжения плавно чередуются с участками расслабления, в результате чего мышечное напряжение может поддерживаться в течение длительного времени (Поза, удержание положения тела)

Типы двигательных единиц Медленные неутомляемые Быстрые устойчивые к утомлению Быстрые утомляемые Чем больше в мышце процент медленных волокон, тем более она приспособлена к работе на выносливость. Чем больше процент быстрых сильных волокон, тем более она приспособлена к работе, требующей большой силы и скорости сокращения мышц.
 Низкие пороги активации мотонейронов Небольшая толщина аксона и скорость проведения")
Медленные двигательные единицы (неутомляемые) Низкие пороги активации мотонейронов Небольшая толщина аксона и скорость проведения возбуждения по нему. Низкая частота разрядов. Начинают функционировать при малых мышечных усилиях. Способны поддерживать постоянную частоту разрядов в течение десятков минут. Развивают небольшую силу при сокращении в связи с наличием в них небольшого, количества миофибрилл. Скорость сокращения этих волокон в 1, 5 -2 раза меньше, чем быстрых, т. к. низкая активность миозин АТФ-азы и скорость выхода ионов кальция и его связывания с тропонином. Малоутомляемы. Хорошо развита капиллярная сеть. Достаточное количество кислорода. Большое количество митохондрий и высокая активность окислительных ферментов.
 Крупные. Толстый аксон, разветвляющийся на большое число концевых веточек и")
Быстрые двигательные единицы (утомляемые) Крупные. Толстый аксон, разветвляющийся на большое число концевых веточек и иннервирующий большую группу мышечных волокон. Высокий порог возбуждения. Высокая скоростью проведения нервных импульсов. Содержат большее число сократительных элементов ― миофибрилл, поэтому при сокращении развивают большую силу. Высокая активность миозиновой АТФ-азы, следовательно высокая скорость сокращения. Много гликолитических ферментов. Мало митохондрий и миоглобина, капилляров. Быстро утомляются. Выполнение кратковременной, но мощной работы.
 Промежуточные положения между ДЕ I и II- В")
Быстрые двигательные единицы (устойчивые к утомлению) Промежуточные положения между ДЕ I и II- В типов. Сильные, быстро сокращающиеся волокна. Большая аэробная выносливость. Используют для получения энергии как аэробные, так и анаэробные процессы.

ВИДЫ МЫШЕЧНЫХ СОКРАЩЕНИЙ В зависимости от частоты импульсации мышца сокращается по разному: Вид сокращения Одиночное мышечное сокращение Зубчатый тетанус Гладкий тетанус Частота раздражителя 6 -8 Гц 20 -40 Гц 50 Гц и выше.

Тетаническое мышечное сокращение Импульсы следуют с частотой большей, чем период одиночного мышечного сокращения, т. е. мышца, не успев расслабиться, получает следующий. В мышцах возникает явление суммации, в результате которого они приходят в состояние длительного укорочения, называемого ТЕТАНУСОМ. Различают: Зубчатый тетанус - При частоте, когда каждый последующий нервный импульс воздействует на начавшую расслабляться мышцу, при этом происходит неполная суммация сокращений. Гладкий тетанус(бо льшая амплитуда) - при частоте раздражения, когда каждое последующий воздействие импульса происходит в конце периода укорочения возникает длительное непрерывное сокращение, что приводит к полной суммации сокращений.
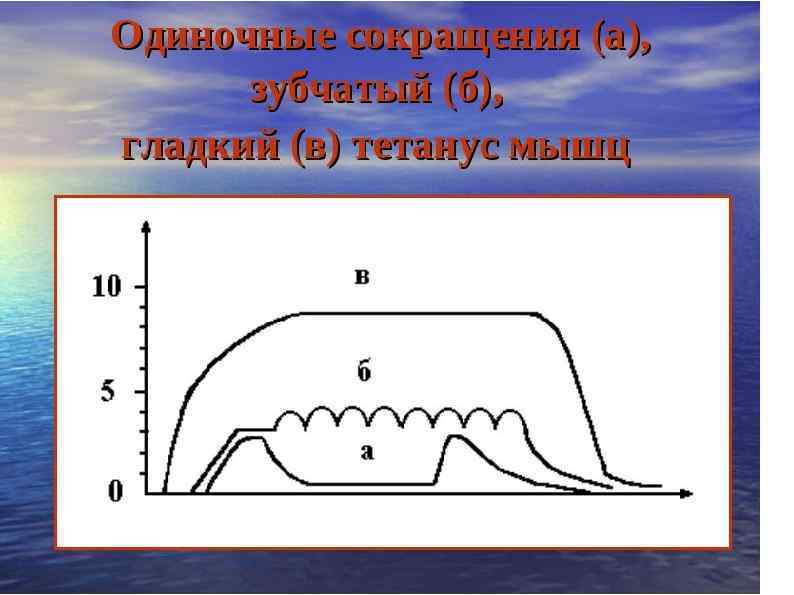

Латентный период - 0, 01 сек. Период укорочения - 0, 04 сек. Период расслабления - 0, 05 сек. Период сокращения мышцы - 0, 1 сек.

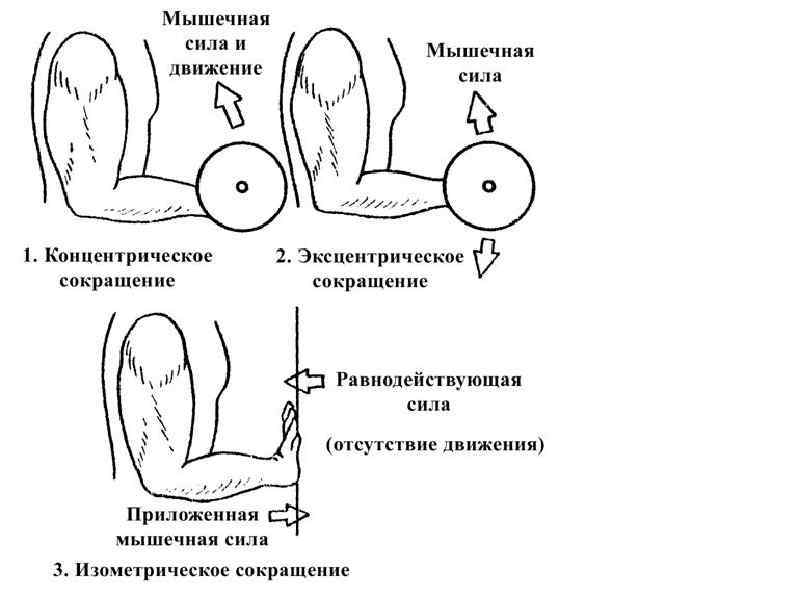
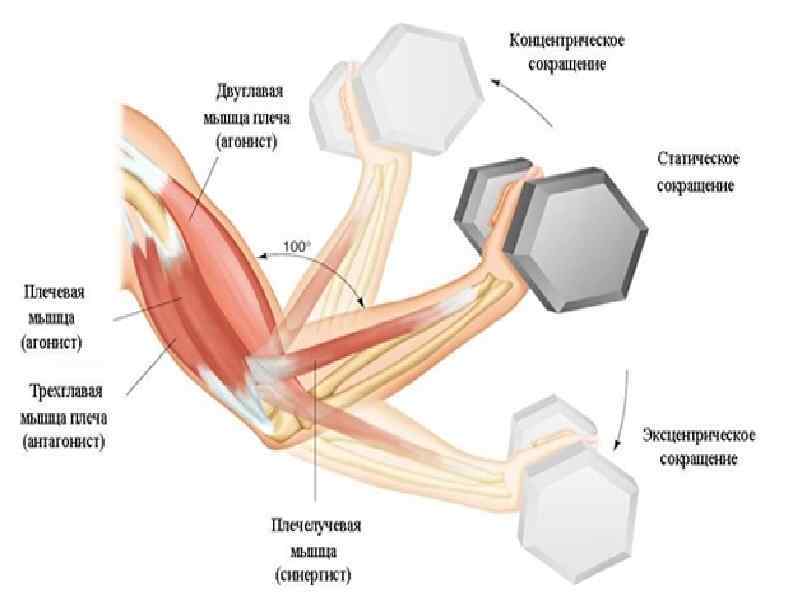
Типы мышечного сокращения 1. Изотонический ― мышечные волокна укорачиваются при постоянной внешней нагрузке. В реальных движениях чисто изотоническое сокращение практически отсутствует 2. Изометрический ― мышца развивает напряжение без изменения своей длины. Изометрическое сокращение лежит в основе статической работы. 3. Ауксотонический или анизотонический тип ― мышца развивает напряжение и укорачивается. Ходьба, бег и т. д. При динамической работе выделяют: 1. Концентрический тип сокращения ― когда внешняя нагрузка меньше, чем развиваемое мышцей напряжение. При этом она укорачивается и вызывает движение; 2. Эксцентрический тип сокращения ― когда внешняя нагрузка больше, чем напряжение мышцы. В этих условиях мышца, напрягаясь, все же растягивается (удлиняется), совершая при этом отрицательную (уступающую) динамическую работу

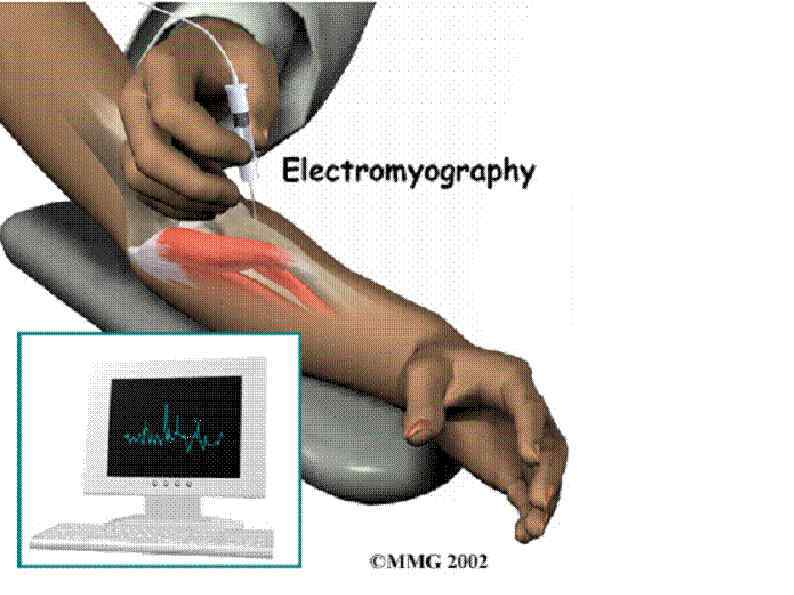
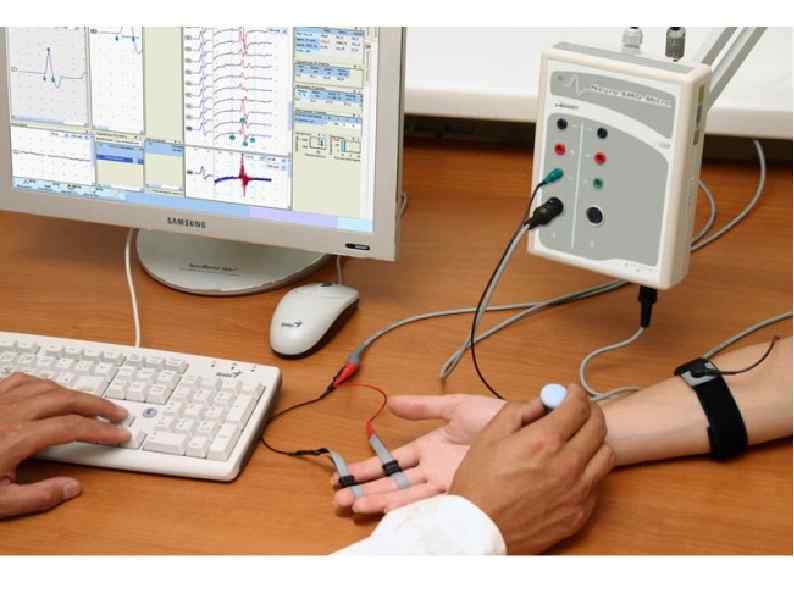
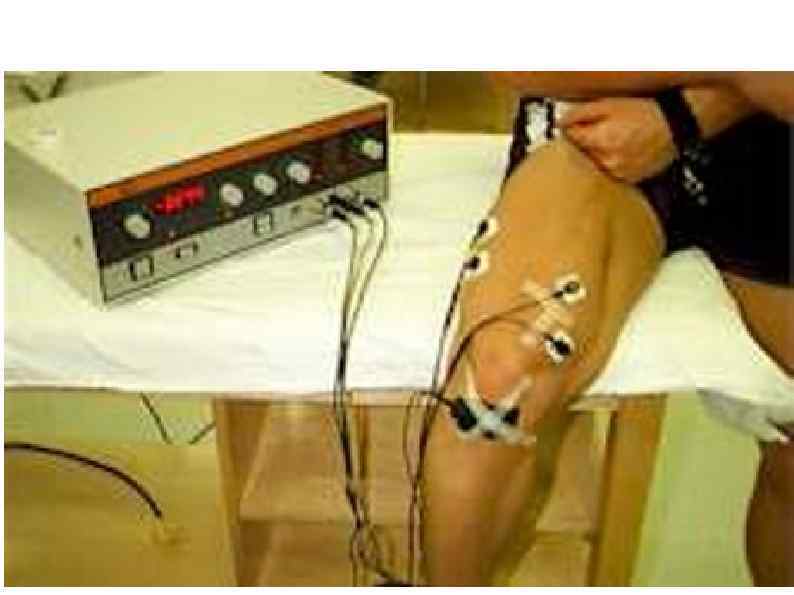
ЭЛЕКТРОМИОГРАФИЯ исследование электрической активности мышц. Исследование проводится в состоянии покоя и во время произвольного сокращения. Могут использоваться поверхностные либо игольчатые электроды. ЭМГ – позволяет получить информацию о состоянии мышечных волокон, их иннервации, количестве и организации двигательных единиц.

Физиология,практика3, мышцы.ppt