Лекция 11-12 биологи ред.pptx
- Количество слайдов: 150


 0 Комменсализм Хищничество Детритофагия Паразитизм Нейтрализм 0 (в т.")
+ + Мутуализм (протокооперация, симбиоз) 0 Комменсализм Хищничество Детритофагия Паразитизм Нейтрализм 0 (в т. ч. условный) Аменсализм (аллелопатия, антибиоз) Конкуренция

Детритофагия detritofagia

ОПРЕДЕЛЕНИЕ • Детритофагия – использование видом в качестве пищи детрита (мертвого органического вещества или экскрементов – непереваренного животными орг. в-ва). Детритофаги или редуценты не могут влиять на собственную кормовую базу нет dr 1 обратной d. N связи 2 dr 2 d. N 1 0

экосистемное определение • В экосистемах детритофаги выполняют функции редуцентов всех порядков, т. е. разлагают отмершее органическое вещество
 всегда действуют на детрит в комплексе с")
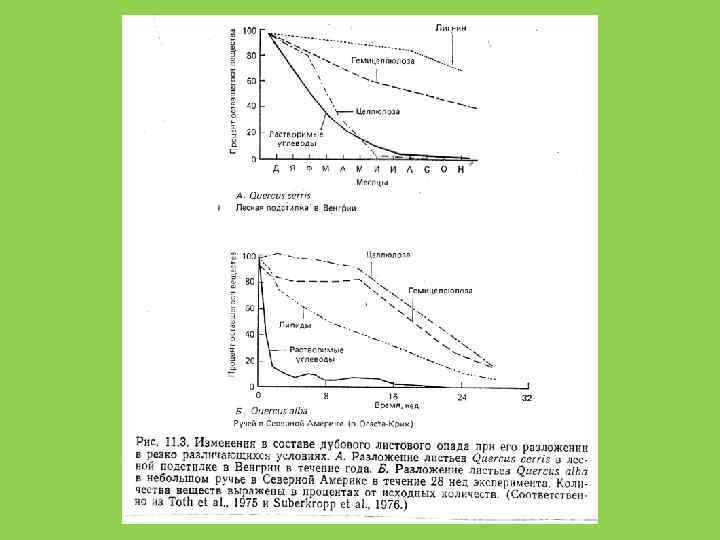
Особенности детритофагии • Редуценты (бактерии и грибы) всегда действуют на детрит в комплексе с детритофагами (животными, питающимися детритом) при этом взаимно повышается их эффективность • Детритофаг – хищник (консумент) и редуцент «в одном флаконе» • Детритофаги часто питаются не столько детритом, сколько редуцентами которые находятся на нем • Редуценты разлагают детрит до первичных веществ нужных растениям и определяют т. о. скорость первичной продукции • По мере разложения растительной ткани специализация редуцентов возрастает

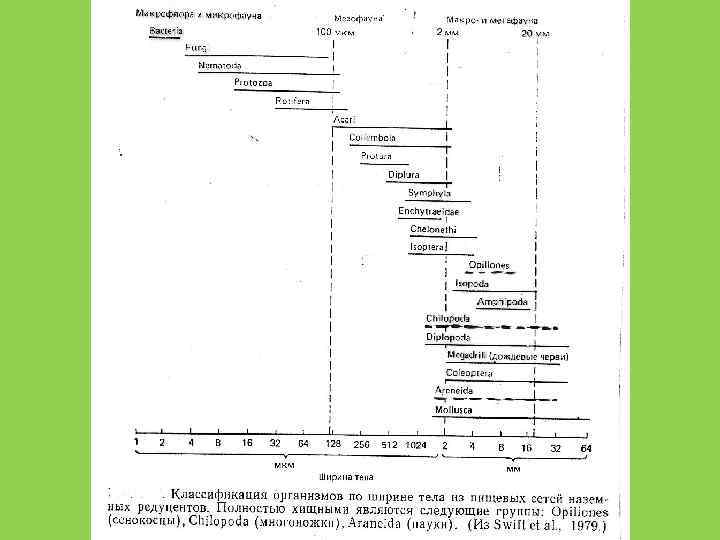
Особенности детритофагии • В водной среде нет детритофагов – специалистов • Только в водной среде есть фильтрация детрита • Существуют сообщества состоящие только из детритофагов, редуцентов и теми, кто ими питается
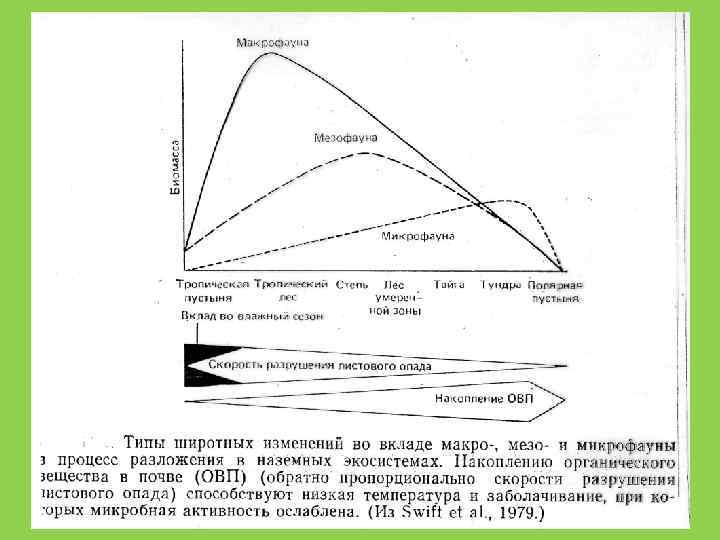
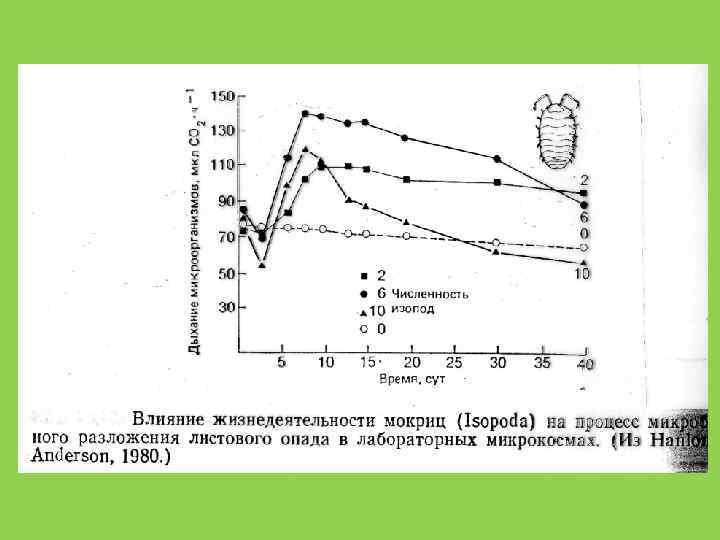
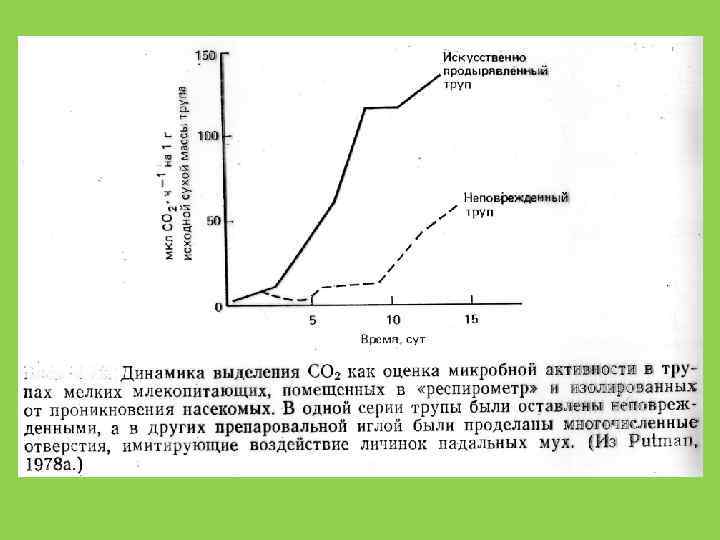

Отношение C : N В микроорганизмах 10: 1 В наземных растениях 40 -80 : 1 У животных 10: 1 это дополнительно ограничивает рост редуцентов на растительном детрите и, с другой стороны, ускоряет разложение животных

Копрофаги и трупоеды
 0 Комменсализм Хищничество Детритофагия Паразитизм Нейтрализм 0 (в т.")
+ + Мутуализм (протокооперация, симбиоз) 0 Комменсализм Хищничество Детритофагия Паразитизм Нейтрализм 0 (в т. ч. условный) Аменсализм (аллелопатия, антибиоз) Конкуренция

Мутуализм, симбиоз, протокооперация Mutualism, simbiosys, cooperation

ОПРЕДЕЛЕНИЕ • Мутуализм – форма взаимодействия между видами, заключающаяся во взаимовыгодном обмене ресурсами, что приводит к взаимному повышению организменных или популяционных характеристик dr 1 d. N 2 0 dr 2 d. N 1 0

Мутуализм – не «дружба» , а доведенный до крайней степени эгоизм! (поэтому в нем скрыт потенциальный конфликт)

Наличие мутуализма Прокари оты Царства Бактерии Простейшие Многокле животные точные животные Грибы Растения Бактерии + + + ? + + Простейшие животные Многоклето чные животные Грибы Растения Эукариоты
")
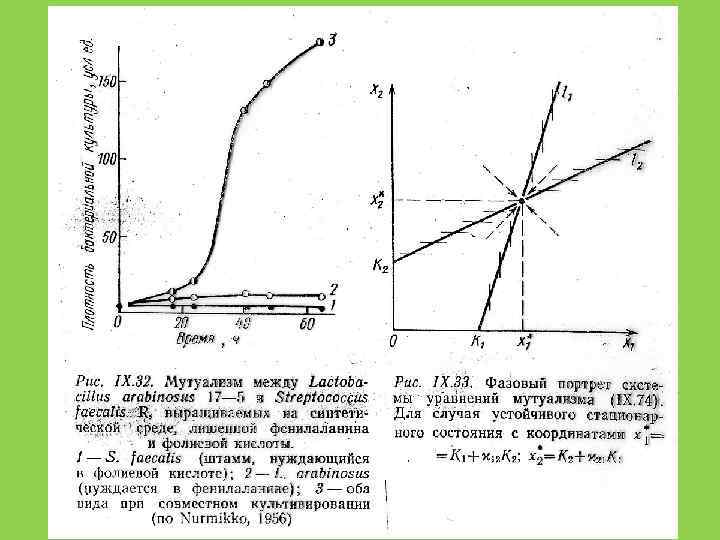
Модель роста двух видовсимбионтов: d. Ni /dt = ri. Ni (1 - Ni /Ki) i = 1, 2 d. N 1 /dt = r 1 N 1 (1 – N 1/(K 1 + μ 12 N 2)) d. N 2 /dt = r 2 N 2 (1 – N 2/(K 2 +μ 21 N 1)) При d. Ni /dt = 0 N 1 = K 1 + μ 12 N 2 = K 2 + μ 21 N 1
 N 2 N*2 K 2 N 02 K 1 N*1")
(N 01, N 02) N 2 N*2 K 2 N 02 K 1 N*1 N*2 N 01 время
 • 1. Мутуализм, включающий поведенческие взаимосвязи (обмен ресурсов: «пища")
Группы примеров мутуализма (1 -4) • 1. Мутуализм, включающий поведенческие взаимосвязи (обмен ресурсов: «пища на пищу» , «пища на защиту» , «защита на пищу + кров» ) • 2. Разведение животных и растений ( «пища на биотоп и расселение» ) • 3. Опыление растений насекомыми ( «пища на размножение и расселение» ) • 4. Мутуализм с участием организмов, населяющих пищевой тракт животных ( «пища на пищу + кров + размножение (биотоп)» )





, совершается самками небольших ночных бабочек Tegeticula yuccasella. Цветы открываются")
Опыление цветков юкки (Yucca filamentosa), совершается самками небольших ночных бабочек Tegeticula yuccasella. Цветы открываются ночью, и на их аромат слетаются самки бабочек, которые при помощи крупных щупиков собирает пыльцу растения. Перелетев на другой цветок, насекомое откладывает в завязь яйца и затем помещает комочек пыльцы в углубление рыльца, производя опыление. Гусеницы питаются молодыми семенами, но съедают лишь часть семян, тогда как остальные семена вызревают.


В одном миллилитре содержимого рубца овцы содержится: 1. 61 × 1010 бактерий 106 жгутиковых 3. 3 × 105 инфузорий объем желудка овцы 6 литров коровы – около 80 литров
 • 5. Симбионты тканей и клеток ( «пища на")
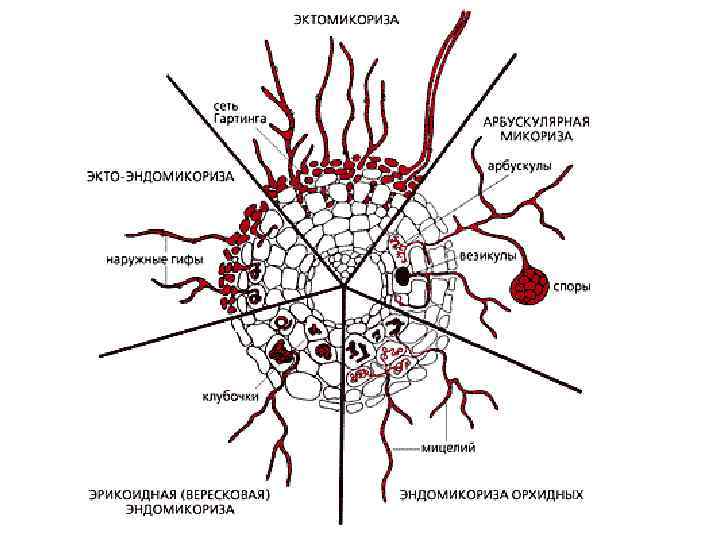
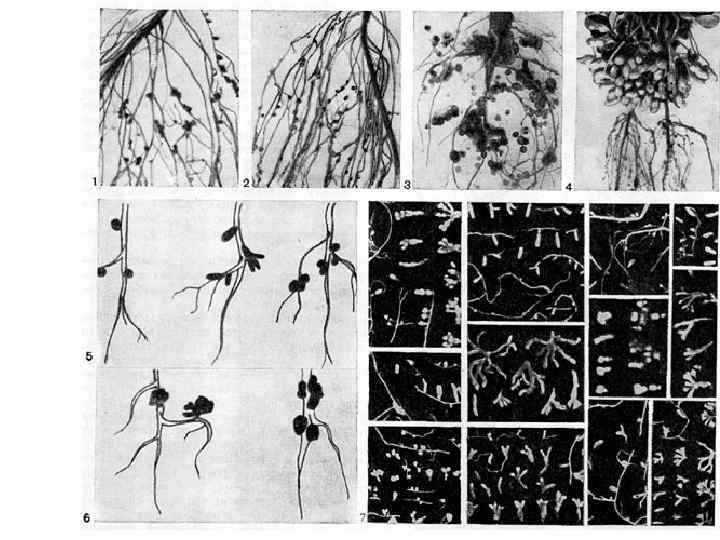
Группы примеров мутуализма (5 -9) • 5. Симбионты тканей и клеток ( «пища на пищу + кров + размножение (биотоп)» ) • 6. Мутуализм высших грибов и растений (микориза)( «пища на пищу, пища на биотоп» ) • 7. Водорослей и животных (коралловые рифы)( «пища на биотоп» ) • 8. Водоросли и грибы (лишайники) ( «пища на биотоп» ) • 9. Мутуалистическая азотфиксация ( «пища на пищу» , «пища на биотоп» )


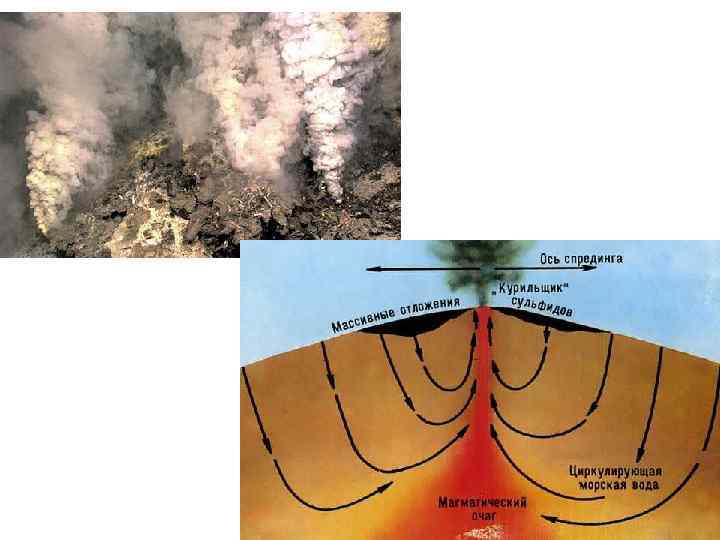
 • 10. Гидротермальные сообщества ( «пища на биотоп» ) открыты")
Группы примеров мутуализма (10) • 10. Гидротермальные сообщества ( «пища на биотоп» ) открыты в 1976 г.
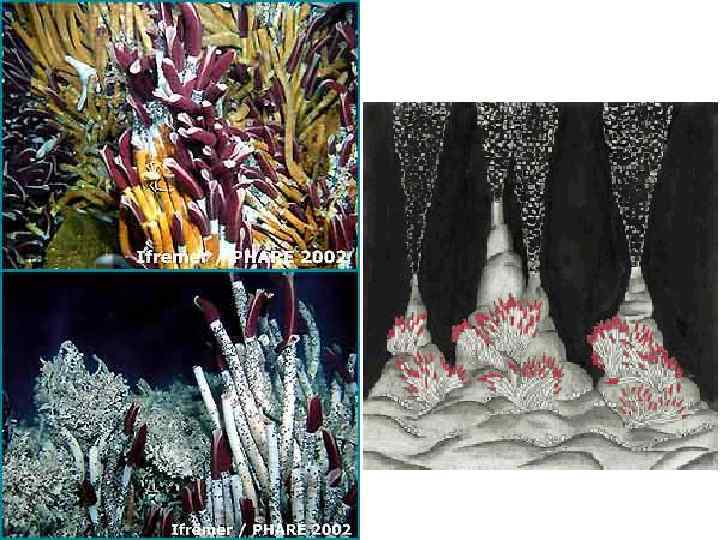
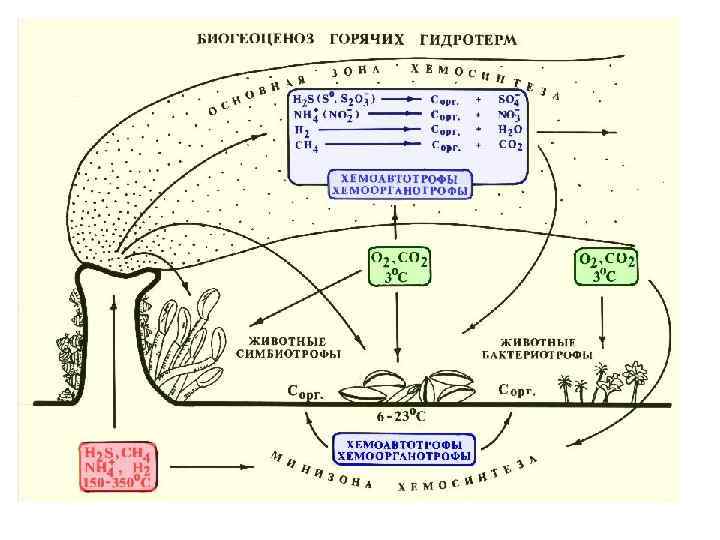
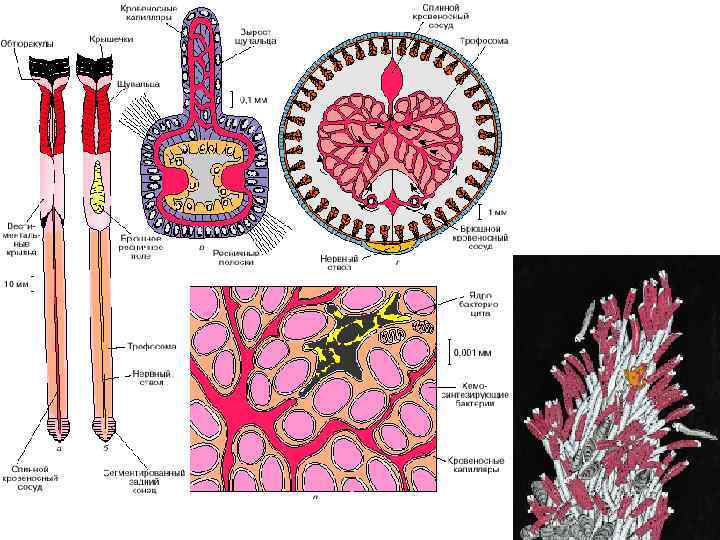

Не вполне очевидные случаи мутуализма «Помошничество между животными, растениями и бактериями» ; «истинный и взаимный альтруизм (мутуализм)»
")
СООБЩЕСТВО многовидовое (community)
 консорция Трофическая группировка (гильдия, ассоциация простое сообщество) синонимы: биотоп, косная среда,")
Биотическая компонента редуцированный) консорция Трофическая группировка (гильдия, ассоциация простое сообщество) синонимы: биотоп, косная среда, окружающая среда С о о б щ е с т в о Абиотическая компонента Биоценоз (полный или синоним: живое вещество Видовая популяция биогеоценоз (по В. Н. Сукачеву), экосистема по А. Тэнсли (1935) экосистема по Ф. Эвансу (1956)
 Видовая популяц и")
С о о б щ е с т в о (особей) Видовая популяц и я еская дия, офич Тр а (гиль ировк групп ростое ция, п ссоциа а ство) е сообщ Биоценоз (полный или редуцирован ный), консорция
: сообщество (как биотическая компонента экосистемы) – группа видов, устойчиво существующая в")
А. Тэнсли (1935): сообщество (как биотическая компонента экосистемы) – группа видов, устойчиво существующая в единообразном абиотическом окружении Ф. Эванс (1956): сообщество – любая группа особей, взаимодействующая с общим абиотическим окружением

A. G. Tensley: “Though the organisms may claim our primary interest, we cannot separate them from their physical environment, with which they form one physical system. ”
 – это объединение видов, которые приспособились к условиям среды и друг")
Биоценоз (Мебиус, 1887) – это объединение видов, которые приспособились к условиям среды и друг к другу, и благодаря размножению непрерывно владеют общим пространством
, консументов (животные) и редуцентов (детритофаги,")
Функциональное определение Биоценоз – группа популяций продуцентов (растений, бактерий), консументов (животные) и редуцентов (детритофаги, грибы и бактерии), приспособившихся к совместному обитанию в общем гомогенном пространстве
 + зооценоз + микробиоценоз + микоценоз")
Таксономическое определение: Биоценоз = (фитоценоз) + зооценоз + микробиоценоз + микоценоз
: ввел в осмысление термина сообщества понятия «пищевая цепь» и «пирамида численностей»")
Charles Elton (1927): ввел в осмысление термина сообщества понятия «пищевая цепь» и «пирамида численностей»
: обосновал точку зрения на сообщество, как термодинамическую систему")
A. J. Lotka (1925): обосновал точку зрения на сообщество, как термодинамическую систему
: объединил концепции экосистемы Тенсли и пищевой цепи Элтона в пирамиду энергетических")
Raymond Lindeman (1942): объединил концепции экосистемы Тенсли и пищевой цепи Элтона в пирамиду энергетических уровней

Сообщества Гильдии Биоценозы Ассоциации Консорции
 не существуют в")
Биоценоз – не существует вне биотопа, так же как фито(зоо-, микробиоценозы) не существуют в отдельности от биоценоза Биоценоз – потенциально не ограничен во времени и пространстве Биоценоз - совокупность всех видов, принимающих активное участие в его функционировании
 – не просто совокупность видов и особей, это и их взаимосвязи Эмерджентные")
Биоценоз (сообщество) – не просто совокупность видов и особей, это и их взаимосвязи Эмерджентные свойства: 1. видовое разнообразие 2. трофическая структура
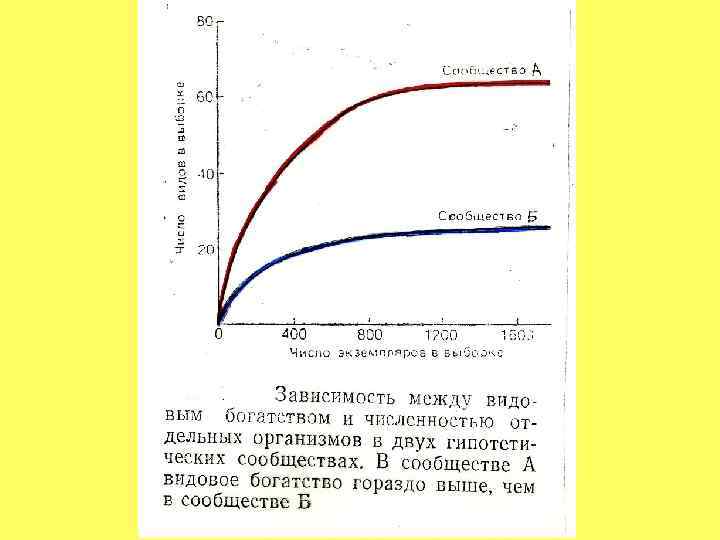
: А) ПТИЦ ДЛЯ РАЗНЫХ РАЙОНОВ США; Б)")
КРИВЫЕ НАКОПЛЕНИЯ ВИДОВ ( «КРИВЫЕ СБОРЩИКА» ): А) ПТИЦ ДЛЯ РАЗНЫХ РАЙОНОВ США; Б) БЕНТОСА СЕВЕРНОГО МОРЯ А ЧИСЛО ВИДОВ Б ЧИСЛО ПРОБ
 пресмыкающиеся Карибских островов Б) млекопитающие Скалистых Гор")
ЭМПИРИЧЕСКИЕ ЗАВИСИМОСТИ ЧИСЛА ВИДОВ ОТ ПЛОЩАДИ: А) пресмыкающиеся Карибских островов Б) млекопитающие Скалистых Гор (США)
 и выровненности (evenness)")
Видовое разнообразие оценивается по видовому богатству (species richness) и выровненности (evenness)
 (Whittaker, Pianka) 2. Индексы разнообразия")
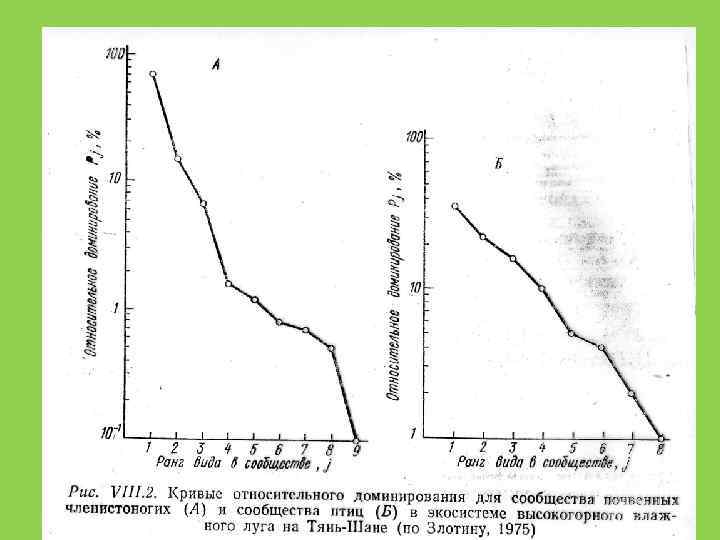
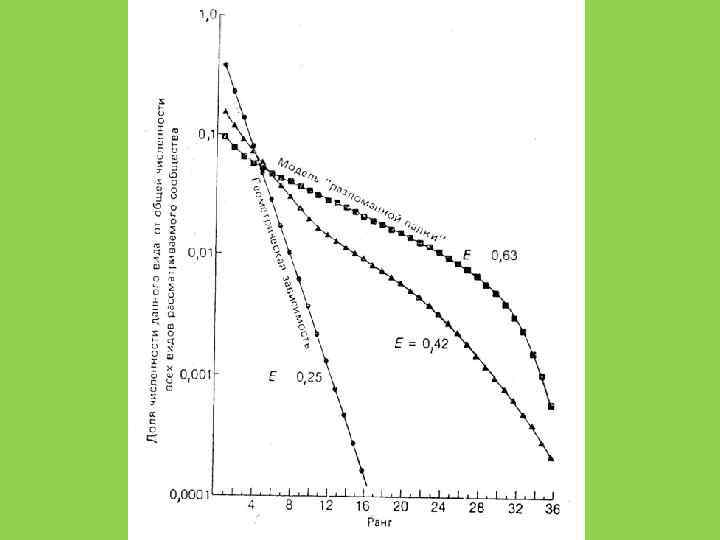
Способы оценки видового разнообразия 1. Кривые доминированияразнообразия (значимости) (Whittaker, Pianka) 2. Индексы разнообразия
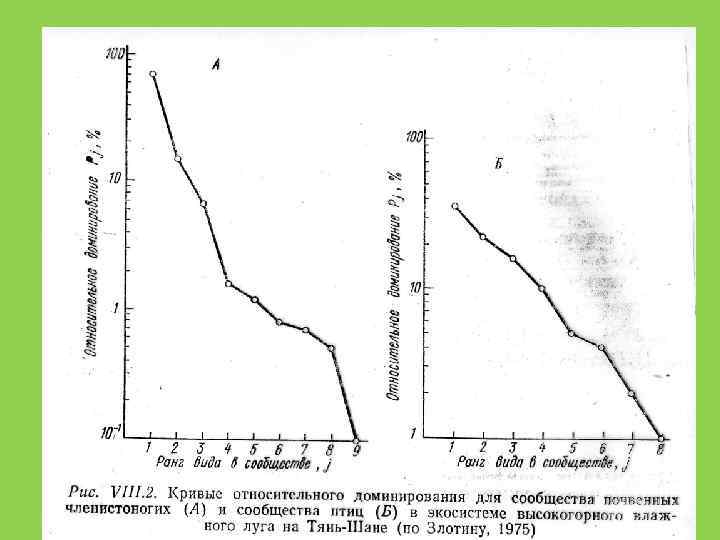
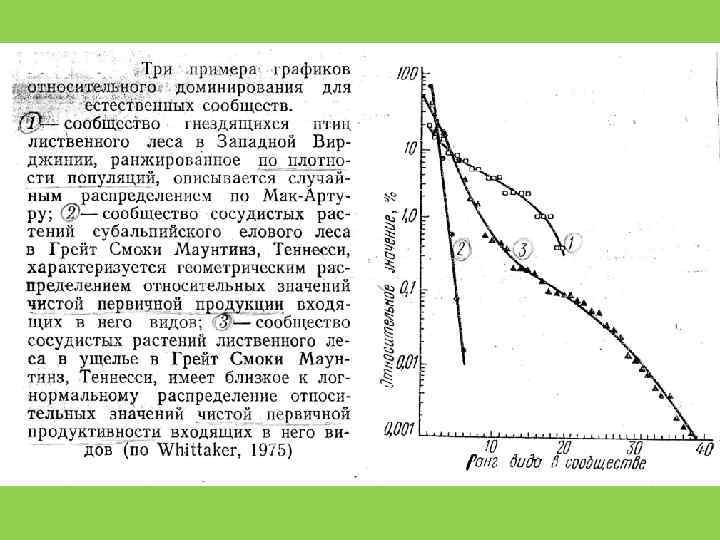
 Экологические ниши случайны и не перекрываются")
Распределение Роберта Макартура (модель «разломанного стержня» ) Экологические ниши случайны и не перекрываются

Геометрическое распределение Экологические ниши одномерны, плотно «упакованы» , и не перекрываются

Логнормальное распределение
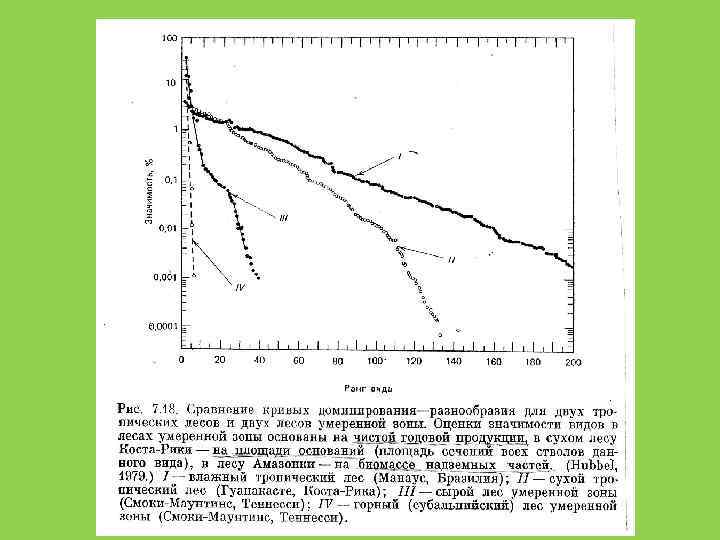

Когда мы говорим о видовой структуре сообществ, то обычно подразумеваем не столько их видовой состав, сколько соотношение численностей разных видов

Индекс видового разнообразия должен учитывать не только число видов, но и соотношение численностей (биомассы, продукции) разных видов
/ln. N S/1000 S – числов видов Индекс видового разнообразия")
Индексы видового богатства S/N (S-1)/ln. N S/1000 S – числов видов Индекс видового разнообразия Симпсона (Simpson): D = 1/ Σ(Pi 2) i = 1, S Pi- доля i-го вида в общей численности или D = (1 – Σ(Pi 2)) E = D/ Dmax = D/ 1/(S(1/S)2 = D/S
")
Индекс разнообразия Шеннона (Shannon)

1. Проба фитопланктона – «текст» 2. Разные виды – разные буквы 3. Численности видов – частоты букв в тексте 4. Минимальное количество двоичных символов, которыми можно записать текст, определяется формулой Шеннона 5. Данная формула может использоваться для оценки видового разнообразия сообщества
 J = H /Hmax = H /-(S(1/S ln (1/S)) =")
Индекс выровненности Шеннона (Shannon) J = H /Hmax = H /-(S(1/S ln (1/S)) = H /-ln(1/S) = H /ln. S
 и 2. Чем")
Показатели видового разнообразия тем выше: 1. Чем больше число видов (S) и 2. Чем более равномерно соотношение численностей разных видов (чем менее выражено доминирование немногих видов)

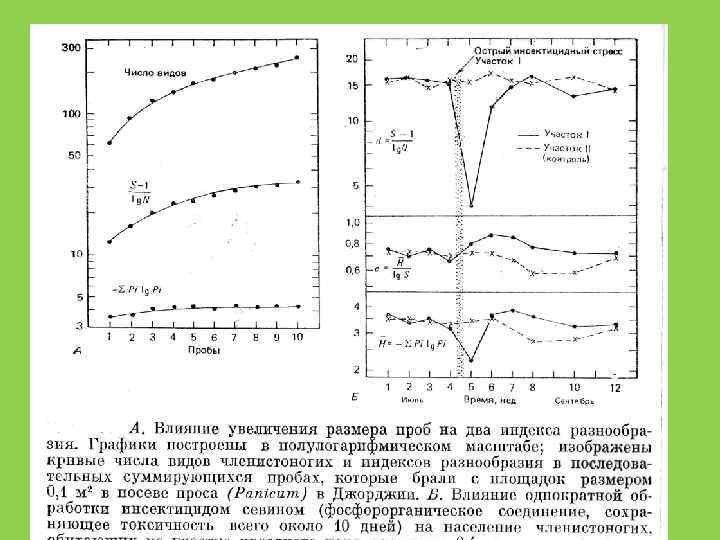
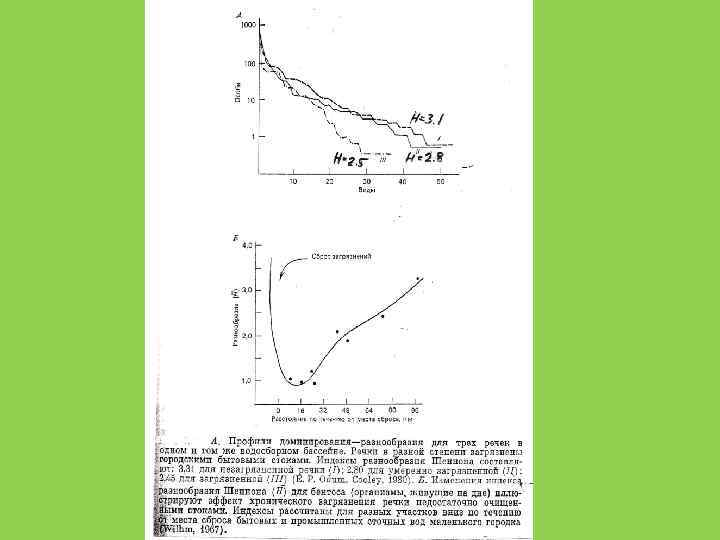

Видовое разнообразие и пространственный масштаб

ВИДОВОЕ БОГАТСТВО И МАСШТАБ • КАКАЯ ГРУППА БОГАЧЕ ВИДАМИ: ИНФУЗОРИИ ИЛИ ПОЗВОНОЧНЫЕ?
 α 1 α β POINT (SAMPLE) DIVERSITY γ")
УРОВНИ РАЗНООБРАЗИЯ по УИТТЕККЕРУ (Whittaker, 1960) α 1 α β POINT (SAMPLE) DIVERSITY γ α 3

= γ / β ab ab a = 2 β = 1 γ = 2 ab d c e a = 1. 25 β = 4 γ = 5

ВИДОВОЕ БОГАТСТВО ЗАВИСИТ ОТ МАСШТАБА ЕГО НУЖНО НОРМИРОВАТЬ. КАК? • НА ПЛОЩАДЬ (НА м 2 ? НА ГЕКТАР? ) • НА РАЗМЕР ОСОБИ? • НА ЧИСЛО ОСОБЕЙ?

103 106 109 Относительные диапазоны пространственной неоднородности сообществ бентоса литорали Белого моря (по А. И. Азовскому, 2001). Среднее сходство (Съеренсен-Чекановский): F = 2 C/ (A+B)
")
Из работ А. И. Азовского (Azovsky)
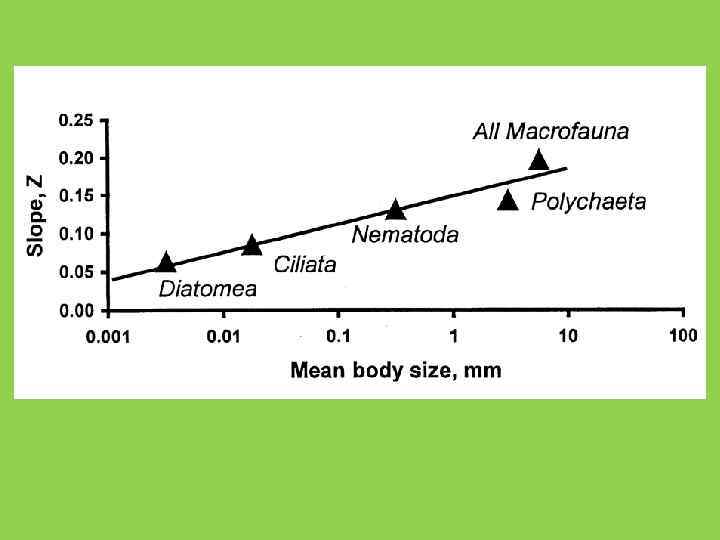

НАКЛОН КРИВЫХ «ВИДЫ-ПЛОЩАДЬ» И СТЕПЕНЬ ИЗОЛЯЦИИ ТИПИЧНЫЕ ЗНАЧЕНИЯ z: ОСТРОВА: 0. 2 - 0. 33 МАТЕРИКИ: 0. 12 - 0. 17 МУРАВЬИ ИНДОНЕЗИИ (по: Wilson, 1961) Вверху - районы Новой Гвинеи; Внизу - отдельные острова
 высшие растения Микронезии Б) птицы озер")
КРИВЫЕ «ВИДЫ-ПЛОЩАДЬ» ПОД МИКРОСКОПОМ: «ЭФФЕКТ МАЛОГО ОСТРОВА» А) высшие растения Микронезии Б) птицы озер США ОБЫЧНОЕ ОБЪЯСНЕНИЕ: ОГРАНИЧЕННЫЙ ПУЛ ВИДОВ + СЛУЧАЙНЫЕ ( «МЕСТНЫЕ» ) ФАКТОРЫ

ВЫВОД: ЧИСЛО И СКОРОСТЬ ПРИРОСТА ВИДОВ В СООБЩЕСТВЕ ЗАВИСЯТ ОТ: • ХАРАКТЕРА СРЕДЫ (ИЗОЛИРОВАННОСТЬ, НЕОДНОРОДНОСТЬ) • МАСШТАБА ПРОСТРАНСТВА • РАЗМЕРОВ И ПОДВИЖНОСТИ ОРГАНИЗМОВ

Осмысление проблемы организации сообществ началось в 20 -е годы ХХ века Тогда были сформулированы ДВА ПОДХОДА:
 Концепция «сверхорганизма» Ф. Клеменса (Clements, 1916) (2) «индивидуалистическая» концепция Г. Глизона (Gleason, 1926)")
(1) Концепция «сверхорганизма» Ф. Клеменса (Clements, 1916) (2) «индивидуалистическая» концепция Г. Глизона (Gleason, 1926)
 Сообщество – совокупность разных организмов, обитающих в одном месте и образующих высокоинтегрированное целое,")
(1) Сообщество – совокупность разных организмов, обитающих в одном месте и образующих высокоинтегрированное целое, характеризующееся специфическим развитием, поведением, а возможно, и эволюцией
")
Лидеры двух школ, развивавшие представления о сообществах как целостных образованиях Frederic Clements (1874 -1945) Владимир Николаевич Сукачев (1880 -1967)
 Сообщество – совокупность организмов разных видов, попавших в одно место и нашедших там")
(2) Сообщество – совокупность организмов разных видов, попавших в одно место и нашедших там подходящие условия для обитания. «Целостность» сообщества – артефакт подхода исследователей
")
Еретики, трактовавшие сообщество как условную единицу, выделяемую для удобства исследования Henry Gleason (1882 -1975) Леонтий Григорьевич Раменский (1884 -1953)
")
Каковы принципы устройства сообщества согласно 1 -му подходу? (сообщество, как высоко интегрированное целое)

1. Сосуществующие в сообществе виды обязательно различаются экологически – они занимают РАЗНЫЕ НИШИ
")
2. Подбор сосуществующих видов происходит по принципу КОМПЛЕМЕНТАРНОСТИ НИШ (“niche assembled communities” )

3. Конкуренция между особями разных видов гораздо СЛАБЕЕ конкуренции между особями одного вида

Каковы принципы устройства сообщества согласно 2 -му подходу? (сообщество, как набор особей разных видов, оказавшихся в одном месте)
")
1. Подбор сосуществующих видов определяется прежде всего особенностями РАССЕЛЕНИЯ организмов (“dispersal assembled communities”)

2. Сосуществующие виды могут быть ЭКОЛОГИЧЕСКИ БЛИЗКИМИ. Cходство может быть результатом отбора, протекавшего в сходных условиях и приведшего к конвергенции, а не дивергенции ниш

3. Конкуренция между особями разных видов по крайней мере НЕ СЛАБЕЕ конкуренции между особями одного вида
 • Широкое перекрытие ниш и")
Принцип Л. Г. Раменского – Г. Глизона (Gleason, 1926) • Широкое перекрытие ниш и рассредоточенность видов вдоль градиента условий среды приводит к плавному переходу одного сообщества в другое
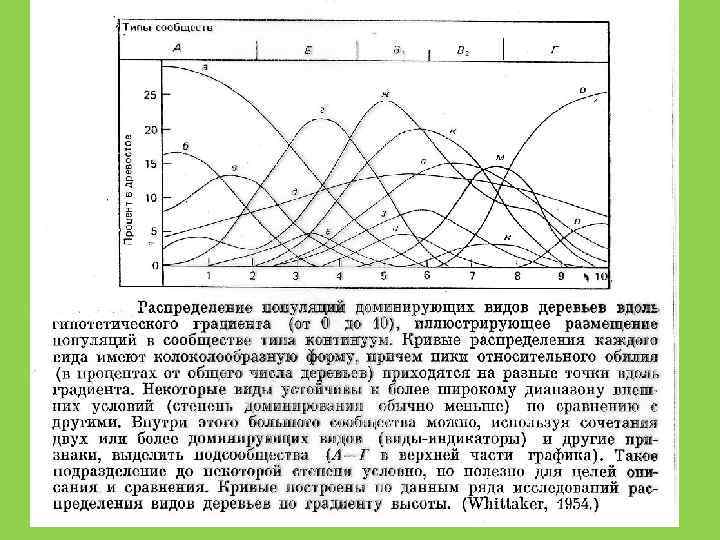
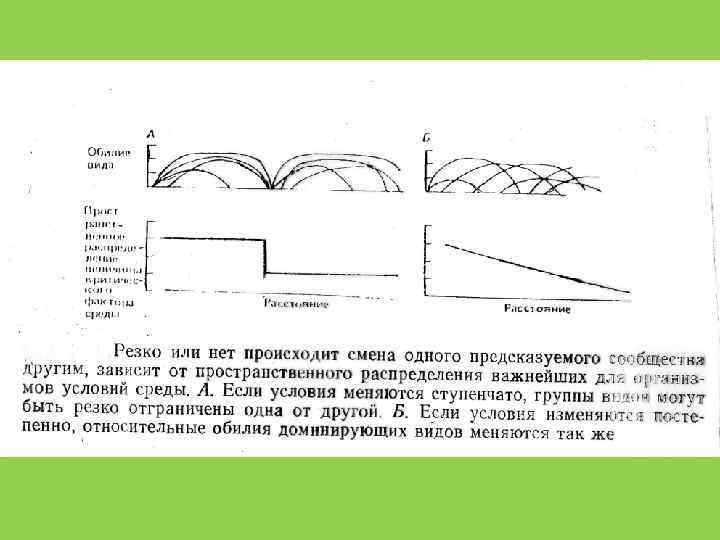
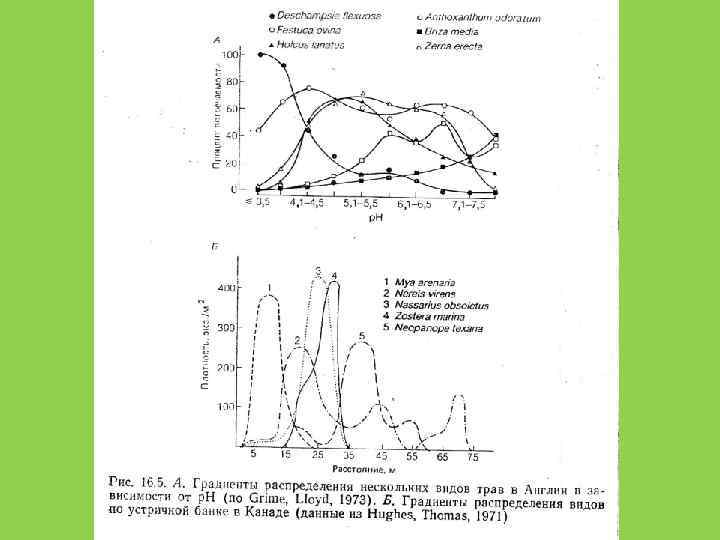
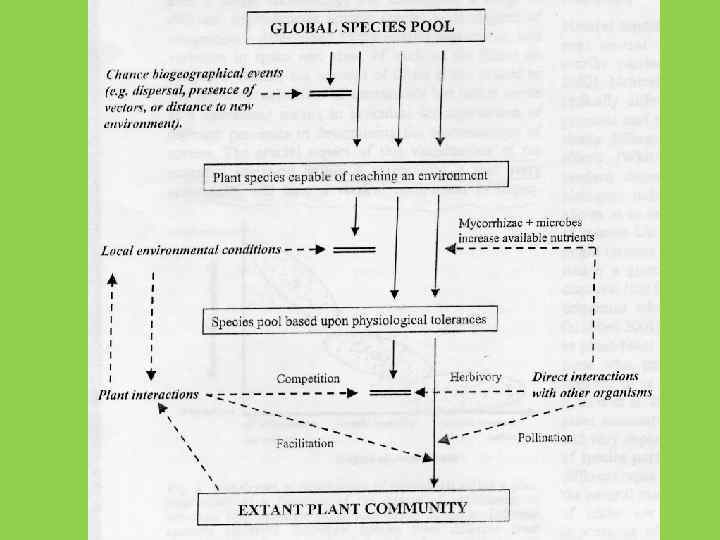
 и Э. Уилсон (Edward Wilson)")
Теория островной биогеографии Р. Мак Артура (Robert Mac. Arthur) и Э. Уилсон (Edward Wilson) (1967): Объяснение того, почему на каждом из островов, входящих в состав архипелага, обитает меньше видов, чем на равной по площади территории на материке

Для каждого острова существует точка равновесия, при которой скорость заселения и скорость вымирания совпадают (но видовой состав в равновесной точке может меняться)

Чем меньше остров, тем интенсивнее вымирание, и тем меньше равновесное количество видов

Чем ближе остров к материку, тем интенсивнее заселение, и тем больше равновесное количество видов
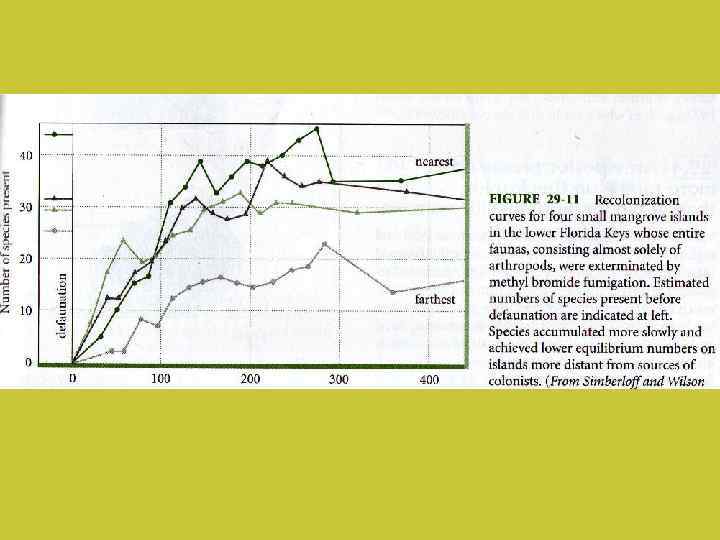

В теории островной биогеографии Макартура – Уилсона не предполагается расхождения видов по разным нишам. Сообщество формируется процессом расселения организмов, а не конкуренцией между уже прибывшими на остров видами Такие сообщества называют “dispersal assembled” в отличие от “niche assembled”

• От каких факторов зависит видовое богатство?
: ЗАКОН, ЗАГАДКА ИЛИ АРТЕФАКТ? А) Рукокрылые Нового Света Б)")
ШИРОТНЫЕ ГРАДИЕНТЫ РАЗНООБРАЗИЯ (А. Уоллес): ЗАКОН, ЗАГАДКА ИЛИ АРТЕФАКТ? А) Рукокрылые Нового Света Б) Морские беспозвоночные
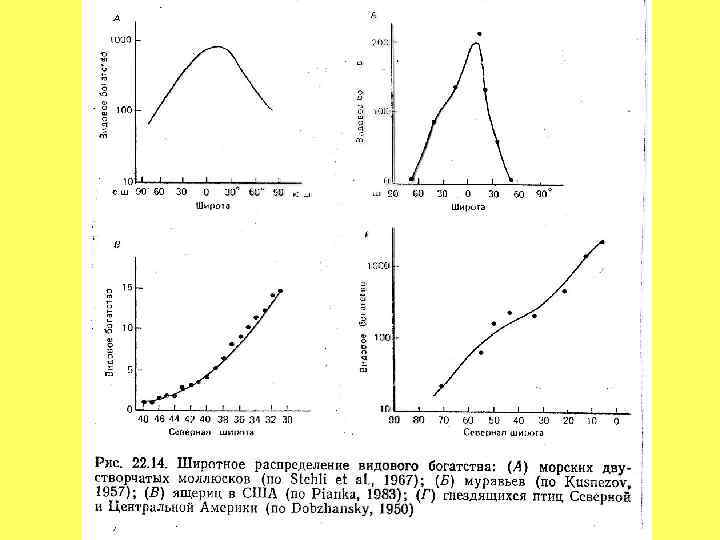
 • ПЛОЩАДЬ (ГЕОМЕТРИЯ) • ЭНЕРГИЯ")
ПРИЧИНЫ И МЕХАНИЗМЫ широтного градиента видового богатства: (ОСНОВНЫЕ ГИПОТЕЗЫ) • ПЛОЩАДЬ (ГЕОМЕТРИЯ) • ЭНЕРГИЯ и ПРОДУКТИВНОСТЬ • СТАБИЛЬНОСТЬ И ПРЕДСКАЗУЕМОСТЬ КЛИМАТА • ГЕТЕРОГЕННОСТЬ СРЕДЫ • ЭВОЛЮЦИОННАЯ ЗРЕЛОСТЬ • СКОРОСТЬ ВИДООБРАЗОВАНИЯ И ВЫМИРАНИЯ • МЕЖВИДОВЫЕ ОТНОШЕНИЯ
 ГИПОТЕЗЫ: • Эффективного Эволюц. Времени: ЭЭВ = время эволюции × скорость")
БОЛЕЕ СЛОЖНЫЕ (КОМБИНИРОВАННЫЕ) ГИПОТЕЗЫ: • Эффективного Эволюц. Времени: ЭЭВ = время эволюции × скорость эволюции • Эффективной Продуктивности: ЭП = средняя продуктивность × стабильность (= продуктивность / VAR продуктивности)
 (А. И. Азовский) 1 -")
ДОСТОВЕРНОСТЬ ШИРОТНЫХ ГРАДИЕНТОВ РАЗНООБРАЗИЯ ДЛЯ РАЗНЫХ РАЗМЕРНЫХ ГРУПП (альфа-разнообразие) (А. И. Азовский) 1 - деревья, 2 – рептилии, 3 – рыбы, 4 - млекопитающие, 5 - амфибии, 6 - птицы, 7 - прочие сосудистые растения, 8 - макроводоросли, 9 - макрозообентос, 10 - насекомые, 11 - зоопланктон, 12 - мейобентос, 13 - простейшие, 14 – микроводоросли.
 3. Гетерогенность среды (разнообразие биотопа) 4.")
1. Эволюционное время 2. Стадия развития сообществ (сукцессии) 3. Гетерогенность среды (разнообразие биотопа) 4. Устойчивость и предсказуемость климата
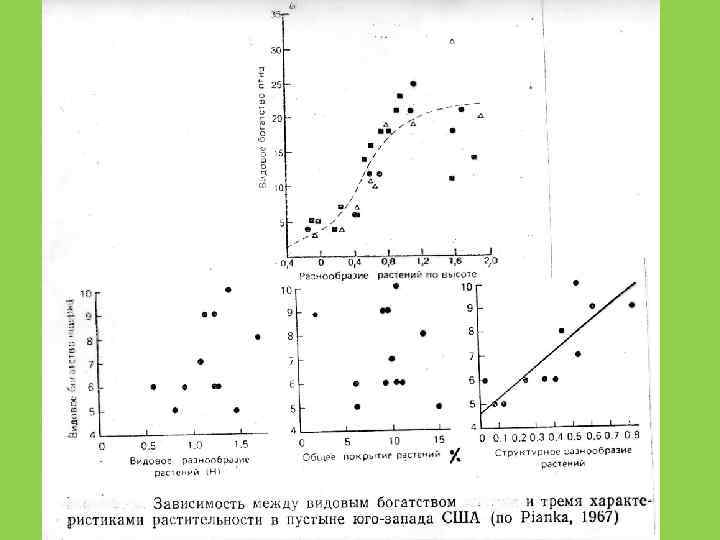
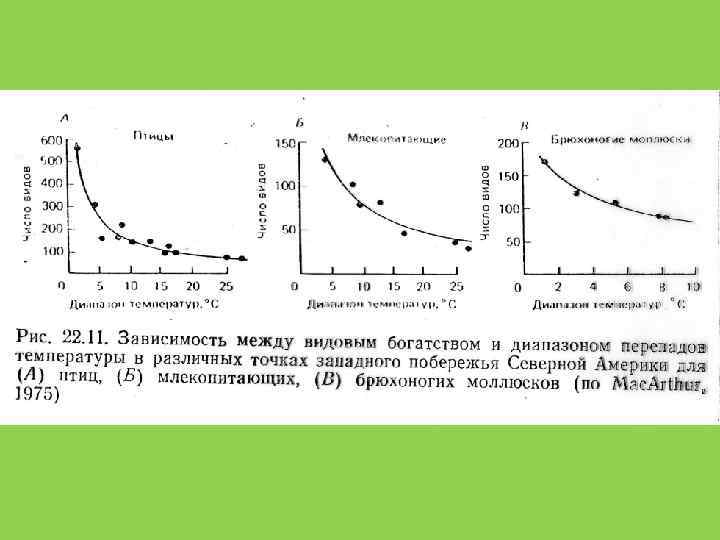

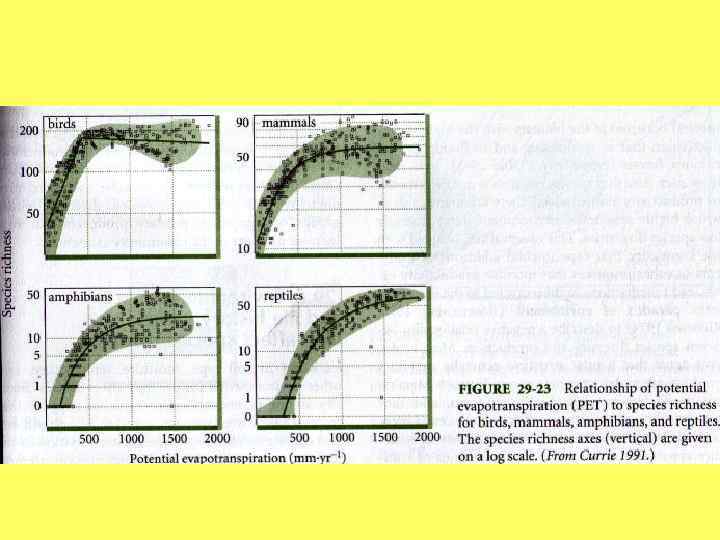
5. Продуктивность сообщества
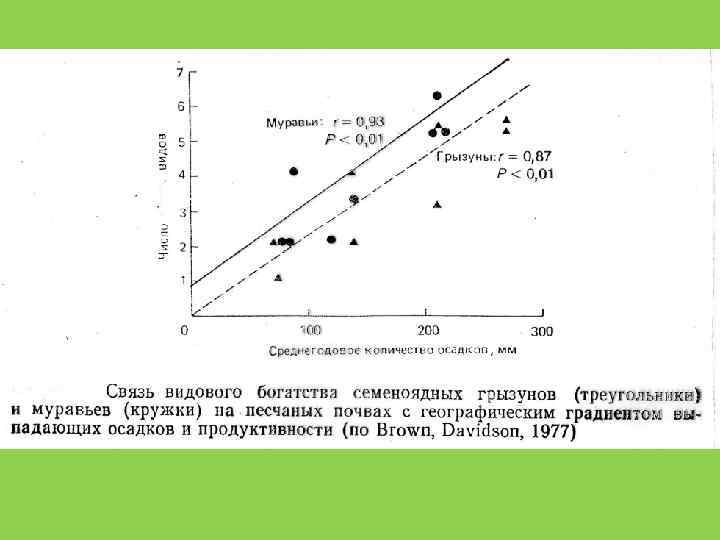
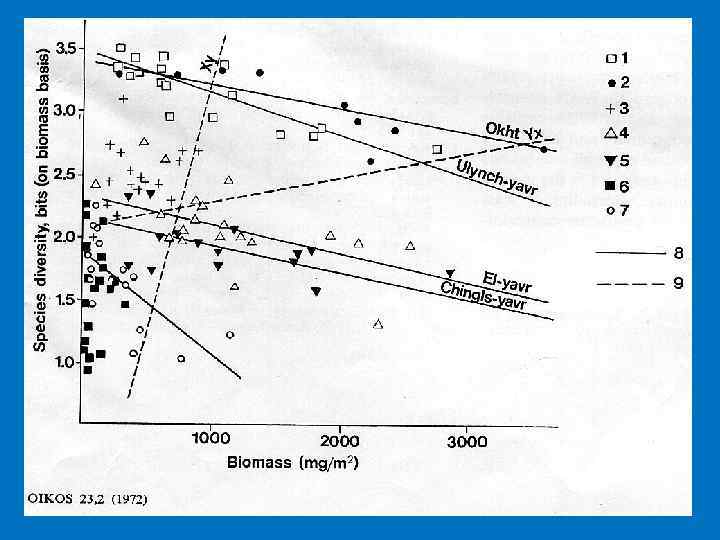
 сообщества")
Видовое разнообразие ? ? ? Продуктивность (биомасса) сообщества

Science 25 August 2000: Vol. 289. no. 5483, p. 1255 M. A. Huston, L. W. Aarssen, M. P. Austin, B. S. Cade, J. D. Fridley, E. Garnier, J. P. Grime, J. Hodgson, W. K. Lauenroth, K. Thompson, J. H. Vandermeer, D. A. Wardle No Consistent Effect of Plant Diversity on Productivity Hector et al. (1) reported on BIODEPTH, a major international experiment on the response of plant productivity to variation in the number of plant species. They found "an overall log-linear reduction of average aboveground biomass with loss of species, " leading to what the accompanying Perspective (2) described as "a rule of thumb--that each halving of diversity leads to a 10 to 20% reduction in productivity. . . We agree with Hector et al. that environmental conditions have a major effect on plant productivity and that overyielding does occur in some multispecies mixtures, particularly those containing nitrogen-fixing legumes. In light of our analysis of the data presented in their figure 2, however, we conclude that species richness per se has no statistically or biologically significant effect on plant productivity across the eight sites of the BIODEPTH experiment.


6. Конкуренция и хищничество

Схема взаимодействия механизмов организации биоценоза по Дж. Коннеллу: 1 - численность популяции; 2 - смертность, вызванная неблагоприятными абиотическими факторами среды обитания; 3 – смертность, обусловленная хищничеством; А – популяции, численность которых ограничивается неблагоприятными физическими факторами; Б - популяции, численность которых ограничивается интенсивным хищничеством

ЧТО ЗАЩИЩАЕТ РЕДКИЕ ВИДЫ ОТ ВЫМИРАНИЯ? В КАКОМ СЛУЧАЕ РЕДКИМ БЫТЬ ВЫГОДНЕЕ ЧЕМ МНОГОЧИСЛЕННЫМ?

Гипотеза Джанзена - Коннелла Воздействие специализированных животных фитофагов и патогенных микроорганизмов резко снижает выживаемость семян и проростков деревьев, если они находятся вблизи материнского растения. Для каждого отдельного растения шанс дорасти до зрелости выше, если оно располагается дальше от взрослых растений того же вида.

Выживаемость проростков Sebastiana longicuspis при разной плотности и поражении патогенными грибами (Thomas Bell, Robert P. Freckleton and Owen T. Lewis. Plant pathogens drive densitydependent seedling mortality in a tropical tree // Ecology Letters. 2006. V. 9. P. 569 -574)

Однако лучшая выживаемость редких видов тропических деревьев в сравнении с массовыми может объясняться не только гипотезой Джанзена - Коннела. Эффект может быть связан с некоторым разделением экологических ниш, в результате чего интенсивность межвидовой конкуренции оказывается ниже внутривидовой Эффект может объясняться положительным влиянием соседних растений в том случае, когда эти растения оказываются другого вида. Это так называемая «гипотеза облегчения» (facilitation)

Возможное разделение ниш: Nathan J. B. Kraft, Renato Valencia, David D. Ackerly Functional Traits and Niche-Based Tree Community Assembly in an Amazonian Forest // Science. 2008. Vol. 322. P. 580 -582 В Yasuni Forest в восточной части Эквадора. Здесь обнаружено максимальное известное на Земле видовое разнообразие деревьев. На модельном участке леса в 25 га (Yasuni Forest Dynamics Plot) - более 1123 видов деревьев (с диаметром ствола не менее 1 см на высоте груди человека). Все эти деревья (более 150 тыс. ) определены до вида, измерены, и нанесены на карту

Возможное разделение ниш: Nathan J. B. Kraft, Renato Valencia, David D. Ackerly // Science. 2008. Vol. 322. P. 580582 В качестве характеристик жизненных стратегий фигурировали: удельная поверхность листьев (площадь листьев, отнесенная к их массе), содержание в листьях азота, средний вес семени, максимальный диаметр на уровне груди (косвенный показатель максимальной высоты дерева), плотность древесины. Некоторые получены для всех 1123 видов деревьев, другие для большей части видов. Мерило пространственной близости деревьев попадание их в одну учетную площадку 20× 20 м. Нуль-гипотеза (нейтрализм) предполагала случайную комбинацию произрастающих вместе видов.
, цветом выделены значения «удельной (на единицу")
А. Показан рельеф местности (горизонтали через 2 м), цветом выделены значения «удельной (на единицу массы) площади листьев» . На «возвышенностях» и в «понижениях» преобладают деревья, различающиеся по удельной площади листьев, что указывает на «фильтрацию» , производимую средой (в долинах преобладают деревья с более тонкими листьями). B. Реальное распределение массы семян деревьев, произрастающих по соседству, то есть попавших на одну учетную площадку 20 × 20 м (сплошная линия — observed quadrat) и распределение, ожидаемое согласно нулевой гипотезе (пунктирная линия — null expectation), предполагающее независимое случайное распределение растений друг относительно друга. Видно, что для деревьев, оказавшихся рядом, наблюдается больший разброс значений этого признака, чем ожидаемый при случайной выборке. Соседствующие деревья разных видов в большей степени различаются по размеру семян, чем это предсказывается на основе случайного их распределения.

Каковы бы ни были конкретные процессы, происходящие в сообществе, очевидно, что вероятность выживания на ранних стадиях у редких видов выше, чем у массовых Эти результаты в принципе работают против гипотезы нейтральности (хотя сам Хаббел является одним из 34 авторов данной статьи )
")
7. Площадь местообитания (размер биотопа)

Может ли сообщество быть насыщено видами?

Существует ли внутренняя регуляция в биоценозах?

Есть ли связь между видовым разнообразием и устойчивостью сообществ?
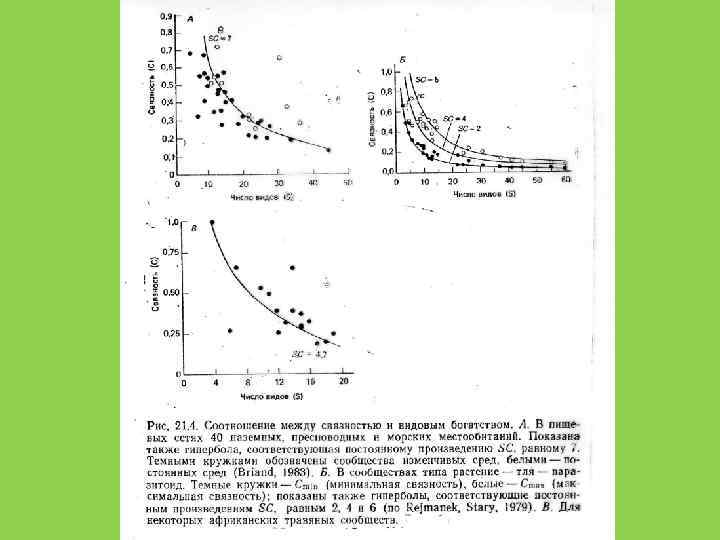
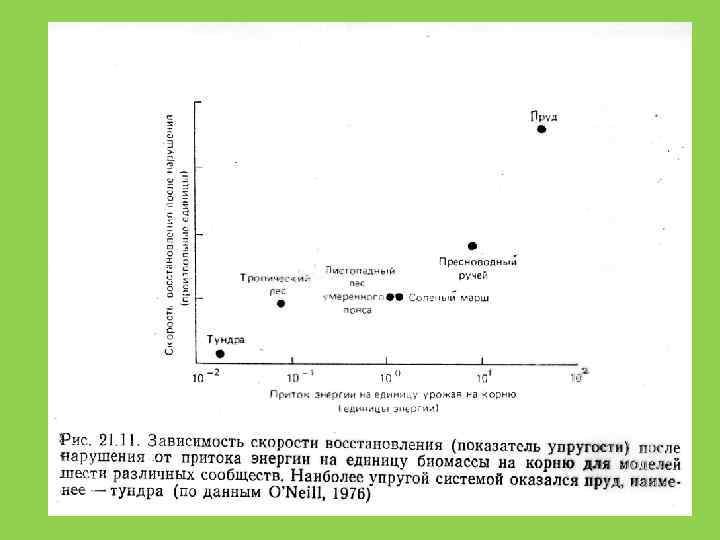

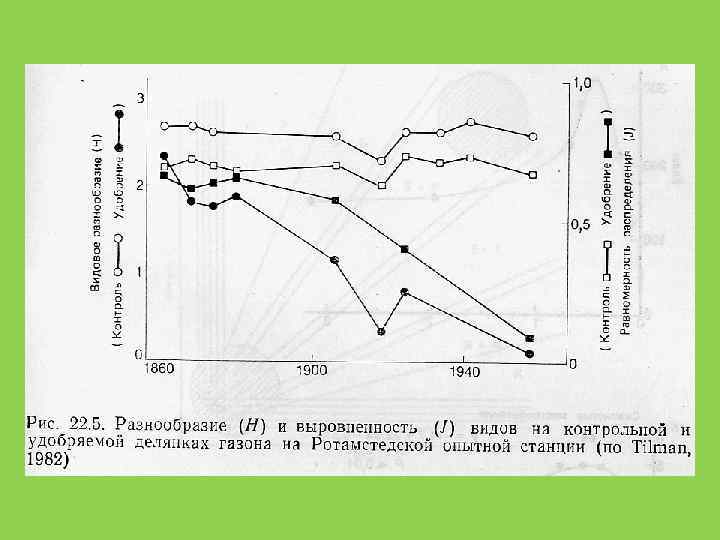
Устойчивость сообщества не является прямой функцией числа видов. Число видов может расти, пока это не снижает эффективность использования сообществом потока вещества и энергии в данной точке

“Волчок жизни” по Н. Ф. Реймерсу: слева - для упрощённой модельной экосистемы Ю. Одума: люцерна - телятина - мальчик (с включением других консументов и редуцентов); справа - для биоценоза Земли в целом

Лекция 11-12 биологи ред.pptx